
Abstract
This investigation aimed to examine the virulence genes and antimicrobial resistance profiles of Shiga toxin–producing Escherichia coli (STEC) strains found in diarrheal calves in Xinjiang between 2016 and 2022. A total of 800 samples, including 232 fecal and 568 rectal swabs from calves under 2 months old with diarrhea, were analyzed for Shiga toxin (Stx)–encoding genes using polymerase chain reaction. The study characterized the isolates based on their stx subtypes, virulence genes, O serogroups, phylogenetic groups, hemolytic phenotypes, antibiotic resistance, and resistance genes. This investigation detected 32 STEC isolates, with stx1 and stx2 genes in 37.5% and 12.5% of the isolates, respectively. Additionally, half of the isolates contained both genes. The combination of stx subtypes was predominantly observed as follows: stx1a/stx2d (31.3%), stx1a only (31.3%), stx1a/stx2a/stx2d (12.5%), stx2d only (9.4%), stx1a/stx1d/stx2d (6.2%), stx1a/stx1d (6.2%), and stx2a/stx2d (3.1%). The eae gene was found in one isolate that carried stx1, stx2, and hlyA, defining it as enterohemorrhagic E. coli. Additionally, six STEC carrying the astA gene were identified as EAST1+ STEC. These isolates did not correspond to the serotypes in the “top seven” category. The phylogenetic analysis revealed that most strains belonged to groups B1 (43.8%) and A (40.6%), with 56.3% exhibiting hemolytic activity. Moreover, among the 34.4% resistant isolates (2021∼2022), the drug resistance rates of STEC isolates to tetracycline, florfenicol, ceftazidime, ampicillin, cefotaxime, gentamicin, and sulphamethoxazole ranged from 21.9% to 34.4%, to streptomycin, piperacillin, ceftazidime, aztreonam–polymyxin B, and cefepime ranged from 3.1% to 18.8%. All isolates exhibited multiple drug resistance. The most common resistance genes identified were floR (31.3%), tetA (25.0%), sulR (15.6%), strA and strB (12.5%), and blaCTX-M-1, blaTEM-1, and mph(A) (9.4%). These findings offer valuable information that might aid in developing preventative measures and enhance the comprehension of the epidemiological characteristics of STEC in calves.
Introduction
Newborn calves infected with enterovirulent Escherichia coli cause significant economic losses for the worldwide cattle industry (Algammal et al., 2020). These pathogenic E. coli strains are categorized based on their virulence traits and the clinical symptoms they elicit in hosts. The categories include Shiga toxin–producing E. coli (STEC), enterohemorrhagic E. coli (EHEC), and enterotoxin-producing E. coli (ETEC). ETEC, STEC, and EHEC are known to induce acute diarrhea in neonatal calves (Chekole et al., 2022).
STEC is the primary pathogen that produces Shiga toxin (Stx), which causes colibacillosis in newborn calves. The Stx family is categorized into two main groups: Stx1, which consists of similar variations like Stx1a, Stx1c, and Stx1d, and Stx2, which is a diverse group encompassing variants from Stx2a to Stx2g (Scheutz et al., 2012). Additionally, EHEC is a subset of STEC that may produce Stx. It is distinguished by LEE, a gene complex that encodes the “intimin” protein, which allows the bacteria to attach to enterocytes (Chekole et al., 2022). ETEC can generate heat-labile enterotoxin (LT), heat-stable enterotoxin (ST), and/or enteroaggregative heat-stable toxin 1 (EAST1), all of which are critical factors in causing severe diarrhea in newborn calves (Ménard and Dubreuil, 2002).
The frequent use of antibiotics in animals and humans to treat diarrhea has inadvertently facilitated the spread of multidrug-resistant (MDR) E. coli bacteria. The existence of this resistance introduces complexity to the progression of the disease, leading to prolonged morbidity and reduced possibilities of recovery. Moreover, antibiotic treatment can stimulate an increase in the release of Stx from deceased bacteria. Additionally, it can lead to alterations in the composition of the gut microbiota, resulting in a more conducive environment for the adherence of STEC to the lining of the intestines (Mühlen et al., 2020). In China, instances of diarrhea in calves infected with STEC have not been well studied and are seldom compared with isolates from humans and healthy cattle in comparative analysis.
Therefore, this investigation aimed to isolate and characterize STEC strains from diarrheal calves in Xinjiang, China. Subsequently, drug resistance and virulence genes were examined to provide baseline data for clinical treatment and farming techniques.
Materials and Methods
Sample collection
A total of 800 samples were collected from 2016 to 2022. These samples included 232 fecal and 568 rectal swabs. All of the samples were obtained from calves under 2 months old who were experiencing diarrhea. The specimens were collected from four major cities (Aksu, Changji, Kashgar, and Urumqi) in China’s Xinjiang region. The sample was obtained using a sterile cotton swab and placed in a tube containing 3 mL of E. coli broth. The samples were returned to the laboratory for analysis and maintained on ice to preserve their integrity.
Bacterial culture and strain isolation
The collected samples were incubated in E. coli overnight at 37°C. The DNA was isolated by the boiling method and was analyzed for stx (stx1 and/or stx2) genotypes via polymerase chain reaction (PCR) (Scheutz et al., 2012). For samples testing positive, serial dilutions were prepared in a 10-fold series. Then, 100 μL from each dilution was evenly spread on MacConkey agar (MAC; Hopebiol) using a coated rod. Five pink-colored colonies as presumptive STEC per sample were subcultured onto MAC and reconfirmed by PCR targeting stx genes. Each confirmed isolate was introduced into brain-heart infusion (BHI; Hopebiol) broth and then placed in an incubator at 37°C overnight. Glycerol stocks were produced from these cultures and stored at −80°C for further analysis.
Subtyping of stx genes
From each positive sample, one isolate was tested for the stx gene subtype. The stx1a, stx1c, and stx1d subtypes were identified using a triple PCR procedure, and the stx2a-stx2g subtypes were determined by the multiplex PCR method described by Scheutz et al. (2012).
Profiling of virulence genes
The identification of virulence genes eae (Bai et al., 2010), hlyA (Paton and Paton, 1998), astA, st, and lt (Zhang et al., 2007) was conducted using specific primers according to previously established protocols.
Serotyping using PCR
The serotyping of STEC isolates, specifically for the “top seven” significant and human-pathogenic serogroups (O26, O45, O103, O111, O121, O145, and O157), was conducted using a multiplex-PCR method developed by DebRoy et al. (2011).
Phylogrouping of STEC strains
The phylogenetic group (A, B1, B2, C, D, E, and F) was determined following the methodology outlined by Clermont et al. (2013).
Hemolytic activity test
The pure cultures of each STEC isolate obtained were then cultivated on Shibam plates. The plates contained BHI medium that was supplemented with 10 mM CaCl2, 5% phosphate buffered saline (PBS)-washed defibrinated sheep blood, and 0.5 μg/mL mitomycin C. The hemolytic activity of each STEC isolate was evaluated at 6- and 24-h intervals, following standard protocols (Lin et al., 2012). The assay was performed twice to ensure the reliability of the data.
Antibiotic susceptibility tests
Following the Clinical and Laboratory Standards Institute 2019 standards, the Kirby–Bauer disc diffusion method was used for antibiotic susceptibility testing. The following antimicrobials were assessed: ampicillin (AMP), piperacillin (PIP), cefotaxime (CTX), ceftazidime (CAZ), cefepime (FEP), aztreonam (ATM), AMP-sulbactam (SAM), PIP-tazobactam (TZP), amoxicillin–clavulanic acid (AMC), gentamicin (GEN), amikacin (AMI), streptomycin (STR), cotrimoxazole (SXT), florfenicol (FFC), levofloxacin, ciprofloxacin, azithromycin (AZM), polymyxin B (PB), and tetracycline (TET; OXOID). E. coli (ATCC25922) was utilized as a quality control strain. Isolates that showed resistance to at least three different classes of representative antimicrobial drugs were classified as MDR (Magiorakos et al., 2012).
Detection of resistance genes
The following resistance genes were amplified by PCR: blaCTX-M-1G (Pagani et al., 2003), blaCTX-M-2G (Saladin et al., 2002), or blaCTX-M-9G (Lin et al., 2010), and blaTEM (Eckert et al., 2004), which encodes β-lactamases, aminoglycoside resistance genes [strA and strB], FFC resistance gene [floR], sulfonamide resistance gene [sulR], macrolide resistance genes [mph(A)], and the tetA gene encoding TET efflux pumps (Shi et al., 2022). Following that, Sangon Biotech (Shanghai) received the PCR products for sequencing. Afterward, the DNA sequences and inferred amino acid sequences were compared with different sequences recorded in GenBank to confirm the subtypes of the resistance gene.
Statistical analysis
The chi-square test examined the positive rates in various regions and sample types. The study employed Statistical Package for the Social Sciences (SPSS) software version 21 (SPSS, Inc., Chicago, IL, USA). A p value < 0.05 was deemed statistically significant.
Results
Prevalence of STEC in diarrheic calves within specific regions of Xinjiang
Table 1 indicates that between 2016 and 2022, 32 STEC isolates were obtained from 232 fecal and 568 rectal swab samples taken from calves with diarrhea. These isolates were recovered using specific isolation and confirmation procedures. Regarding the distribution based on sample type, 15 isolates (6.5%) were obtained from fecal samples, and 17 isolates (3.0%) were collected from rectal swabs. This difference in prevalence rates between the two sample types was statistically significant (p < 0.05). The detection rate of STEC varied across different cities, ranging from 2.3% to 8.2%, with a pronounced difference observed between the Aksu and Changji regions (p < 0.05). Importantly, there were no significant differences observed in the rates of detection across the other regions (p > 0.05). Furthermore, 50% of the STEC isolates contained stx1 and stx2 genes, but a smaller percentage of isolates had the stx1 or stx2 gene, accounting for 37.5% and 12.5%, respectively.
Information of Sample Collection and Identification of Stx Virulence Genes By PCR
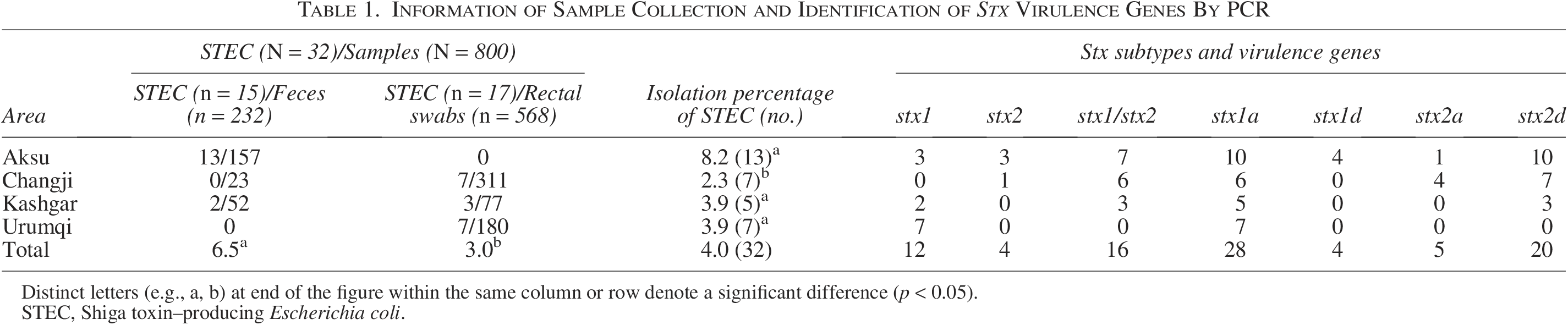
Distinct letters (e.g., a, b) at end of the figure within the same column or row denote a significant difference (p < 0.05).
STEC, Shiga toxin–producing Escherichia coli.
Subtypes of stx1 and stx2 genes
Among the 28 isolates testing positive for stx1, all were classified under the stx1a subtype. Two subtypes were identified for the 20 stx2-positive isolates: 5 instances of stx2a and 20 of stx2d (Table 1). Furthermore, seven different stx subtype profiles were detected across the isolates: stx1a/stx2d (31.3%), stx1a only (31.3%), stx1a/stx2a/stx2d (12.5%), stx2d only (9.4%), stx1a/stx1d/stx2d (6.2%), stx1a/stx1d (6.2%), and stx2a/stx2d (3.1%).
Virulence gene profiling
The presence of eae, hlyA, lt, st, and astA genes was detected via PCR. The presence of pathogenic E. coli strains was investigated as follows: the genes eae, stx1, stx2, and hlyA were examined for EHEC, while the lt gene and/or the st gene were analyzed for ETEC. One (3.1%) isolate harbored eae, stx1, stx2, and hlyA genes, defining it as EHEC. Six STEC isolates were found to carry astA, resulting in EAST1+ STEC strains. No ETEC was detected. In total, 12 diverse virulence profiles were identified, including stx1a/astA (n = 6), stx1a/stx2d (n = 5), stx1a/stx2d/hlyA (n = 4), stx1a/stx1d/stx2d/hlyA (n = 2), stx1a/stx2a/stx2d/hlyA (n = 3), stx1a (n = 4), stx2d (n = 2), stx1a/stx1d/hlyA (n = 2), stx1a/stx2a/stx2d (n = 1), stx1a/stx2d/eae/hlyA (n = 1), stx2a/stx2d/hlyA (n = 1), and stx2d/hlyA (n = 1). No significant difference was observed among cities regarding the prevalence of the virulence gene profiles (Table 2).
Distribution of Virulence Gene Profiles, Phylogenetic Groups, and Hemolytic Activity in 32 STEC Strains Isolated from Diarrheal Calves

STEC, Shiga toxin–producing Escherichia coli; EAST, enteroaggregative heat-stable toxin; EHEC, enterohemorrhagic Escherichia coli; U, Urumqi; A, Aksu; C, Changji; K, Kashgar.
Detected O-serogroups, phylotypes, and hemolytic activity
STEC isolates did not belong to the seven clinically significant serogroups. Regarding phylogenetic categorization, the majority of STEC isolates from calves with diarrhea were mostly found in groups B1 (43.8%) and A (40.6%). It is essential to note that the isolates belonging to phylogroup A were mainly found in Changji. Conversely, individuals belonging to phylogroup B1 were predominantly acquired from Aksu. Markers for phylogroups C and E were detected in four isolates and one isolate, respectively, as shown in Table 2. Eighteen (56.3%) of the 32 STEC isolates had an enterohemolytic phenotype defined by hemolysis, producing a very thin zone surrounding the colony.
Distribution of antibiotic resistance profiles and antibiotic resistance genes
During the analysis period from 2016 to 2020, a substantial proportion of the STEC isolates, specifically 65.6%, did not display any resistance to the evaluated antimicrobials. The resistance rates of STEC isolates to TET, FFC, CAZ, AMP, CTX, GEN, and SXT varied between 21.9% and 34.4%. The resistance rate to STR, PIP, CAZ, ATM, PB, and FEP ranged from 3.1% to 18.8%. Notably, all resistant isolates displayed MDR phenotype, particularly against TET, AMP, and FFC (Table 3). A total of 10 distinct resistance patterns, known as resistotypes, were observed. The most common resistotype was AMP/CTX/SXT/TET resistance, accounting for 63.6% of cases. This was followed by FFC/GEN/STR/TET resistance at 27.3%. In addition, 10 different resistance genes were detected, corresponding to resistance against six classes of antimicrobials (Table 3). The most common resistance genes identified were floR (31.3%), tetA (25.0%), sulR (15.6%), strA and strB (12.5%), and blaCTX-M-1, blaTEM-1, and mph(A) (9.4%).
Characteristics of STEC Isolates Carrying Drug Resistance

AMP, ampicillin; ATM, aztreonam; AZM, azithromycin; CAZ, ceftazidime; CTX, cefotaxime; FEP, cefepime; FFC, florfenicol; GEN, gentamicin; PB, polymyxin B; PIP, piperacillin; STR, streptomycin; SXT, cotrimoxazole; TET, tetracycline.
Discussion
Given the suggestion that AZM can be used to treat STEC infection in humans (Bizot et al., 2022), the public health community should be mainly concerned by the recent finding of STEC strains carrying the mph(A) gene in diarrheal calves. Furthermore, the number of STEC strains from humans and animals has increased the production of beta-lactamases (Elmonir et al., 2021; Joseph and Kalyanikutty, 2022). The analysis revealed a frequent cooccurrence of beta-lactamase genes, such as blaCTX-M and blaTEM, with floR and mph(A) genes in STEC. This suggests that the strains recovered from diarrheal calves are developing higher levels of drug resistance and that resistance genes are accumulating.
The main goal was to investigate STEC prevalence associated with calf diarrhea, explicitly focusing on the virulence genes and evaluating the antimicrobial resistance patterns and related genes. An overall STEC prevalence of 4.0% was found, which was lower compared to findings in diarrheic calves from Egypt (20.2%) (Algammal et al., 2020) and Belgium (62%) (Srivani et al., 2017). However, similar prevalences have been reported in Poland (7.6%) (Osek et al., 2000) and Iran (8%) (Badouei et al., 2010). Factors such as the age of the calves, management procedures, health standards, and geographic location may all influence the variation in the prevalence of STEC identified in different research studies. Specifically, this study found that Aksu had a higher STEC isolation rate than Changji. Variables such as environmental and sanitary settings and the types of samples collected may have contributed to this result.
Virulence genes present in various combinations are important in identifying E. coli pathotypes. The present findings show that stx1 was the most common stx gene, whether found alone or in combination with stx2. This finding aligns with previous studies that have demonstrated the widespread occurrence of stx1 in populations of calves with diarrhea (Badouei et al., 2010; Boriollo et al., 2021). The results of this study showed that both stx1 and stx2 could play a role in causing diarrhea in calves, with the stx1a and stx2d subtypes being the primary carriers. Upon comparison, it is often observed that STEC strains in adult cattle commonly exhibit stx1a and stx2a subtypes (Tong et al., 2021). Epidemiological studies indicate that STEC strains that produce stx2a, stx2c, and stx2d were more frequently found in persons with hemolytic uremic syndrome and hemorrhagic colitis compared with strains that express different stx variants (Melton-Celsa, 2014). Thus, the results suggest that calves could be a substantial source of STEC infections in adults, emphasizing the potential public health implications.
Interestingly, the eae gene was detected in 3.1% of STEC isolates obtained from rectal swab samples collected from calves exhibiting diarrhea. Despite the earlier findings (Tong et al., 2021), which were unable to detect the eae gene in non-O157 STEC isolates taken from adult cattle rectal swabs although the gene’s apparent prevalence in several other studies (Algammal et al., 2020; Badouei et al., 2010).
The hlyA gene was detected in a significant proportion (43.8%) of STEC isolates, indicating its likely link with plasmid encoding. This association enables the gene to be easily transferred across bacteria via plasmids. This is consistent with previous research conducted by Dastmalchi and Ayremlou (2012), which demonstrated that the hlyA gene was present in 57.69% of fecal samples from both healthy and diarrheic calves that tested positive for stx1 and/or stx2.
The EAST1 has also been observed in various E. coli strains, including enteroaggregative E. coli and enteropathogenic E. coli, which are associated with diarrhea in both animals and humans (Ménard and Dubreuil, 2002). The present study has successfully identified the occurrence of the astA gene within STEC isolates. It strongly suggests that these EAST1+ STEC may be a potential public health risk and a factor in the diarrheal symptoms observed in calves.
Furthermore, none of the 32 STEC isolates belonged to the “top seven” serogroups. The predominant serotypes identified in diarrheal calves in Egypt were O128, O26, and O111 (Algammal et al., 2020). Similarly, in Iran, the primary serotype of STEC in diarrheal calves was O26 (Shahrani et al., 2014). This suggests that the serotypes of STEC isolates from diarrheal calves vary across different regions. Interestingly, similar to the results, in China, nearly half of the STEC isolates from patients with diarrhea have also been found to lack distinguishable serotypes (Bai et al., 2016). Although non-O157 STEC isolates of adult cattle and diarrheal calves mainly contain groups A and B1 (Tong et al., 2021), group B1 was more dominant in diarrheal calves. The proportion of hemolytic strains was higher than that of adult cattle, suggesting that attention should be given to the threat of STEC carried by calves to animal and human health.
STEC strains showed significant resistance to three antimicrobial agents: TET, AMP, and FFC. However, a Brazilian study on diarrheal calves found that the most prevalent resistance patterns were against STR, TET, SXT, and cephalothin (Boriollo et al., 2021). Moreover, research conducted in Egypt showed that E. coli bacteria originating from calves with diarrhea had significant resistance to neomycin, GEN, STR, and AMI (Algammal et al., 2020). The drug resistance trends shown in this and other investigations highlight the worldwide concern over an increasing number of MDR organisms, which is fueled by the escalating use of antimicrobial medications in both human health care and cattle farming (Moulin et al., 2008). The upward trend in MDR is most likely due to the accumulation of antibiotic-resistance genes, which can be transmitted via the bacterial chromosome or plasmids (Scheutz et al., 2012).
Conclusions
Calves in specific regions of Xinjiang have diarrhea caused by a diverse collection of Stx-producing E. coli, including STEC and EHEC strains. These strains are becoming more resistant to drugs, particularly TET, AMP, and FFC, and they contain corresponding genes that confer resistance. Plasmid transfer between various strains can facilitate the transmission of this resistance, creating a significant challenge for controlling and preventing calf diarrhea and posing a risk to public health. Thus, it is crucial for cattle farms to systematically regulate antibiotic usage to decelerate and diminish the emergence of bacterial resistance.
Authors’ Contributions
L.W.—Conceptualization, data curation, formal analysis (lead), investigation, methodology, and writing—original draft (lead) and editing. Y.B.—Conceptualization, formal analysis, investigation (lead), and writing. X.S.—Formal analysis and investigation. X.Z.—Supervision and writing—review and editing. Y.L. and Z.S.—Formal analysis, methodology, and writing—review and editing. Y.Z. and M.Z.—Formal analysis. J.X. and P.T.—Supervision and writing—review and editing (equal). All authors contributed to the revising of the article. The authors read and approved the final article.
Footnotes
Acknowledgment
The authors thank Professor H.C.R. Wang for his instructive suggestions in article preparation.
Funding Information
This work was funded by Major Science and Technology Project of the Xinjiang Uygur Autonomous Region, China (Project Name: Key Technologies for Prevention and Control of Common and Multi-Diseases in Cattle, Grant Number: 2023A02007-2) and grants from the National Natural Science Foundation of China (31960695).
Disclosure Statement
The authors declare no conflict of interest.