
Abstract
This study aimed at evaluating the occurrence, antibiotic resistance, and β-lactamase production in Staphylococcus isolates recovered from meat and meat products, as well as the incidence of antimicrobial resistance (AMR) genes in these bacterial isolates. The prevalence of Staphylococcus was very high (75% and 50%) in street kebab and raw buffalo meat, respectively. The antibiotic resistance and susceptibility behavior showed that 82% of the Staphylococcus isolates were resistant to β-lactam antibiotics such as aztreonam, followed by methicillin (68%), oxacillin (54%), cefepime (36%), ceftazidime (34%), cefaclor (24%), cefotaxime (22%), ertapenem (4%), meropenem and imipenem (2%). Among non-β-lactam antibiotics, the most widespread resistance was observed against nalidixic acid (80%), sulfadiazine (76%), vancomycin (24%), erythromycin (10%), chloramphenicol (6%), and kanamycin and gatifloxacin (4%). One hundred percent of the isolates were susceptible to ciprofloxacin, tetracycline, gemifloxacin, and cefotaxime/clavulanic acid. In vitro drug-resistant characteristics revealed 36 distinct resistance patterns of Staphylococcus isolates, with 82% of them being multidrug resistant (MDR). Iodometric assay showed that 48% of the Staphylococcus isolates produced β-lactamase and 24% of the isolates were capable of producing extended-spectrum beta-lactamases phenotypically. The most commonly detected AMR gene was mecA (29.2%), followed by Sul 1 (25%) and qnrS and qnrB (20.8%), in Staphylococcus isolates. Current findings show widespread occurrence of MDR Staphylococcus strains in raw meat and street meat products, which is a potential risk to public health. Therefore, the study suggests strict monitoring of hygiene through the whole food chain and judicious use of antibiotics.
Introduction
The ever-increasing global demand for food has led to the extensive production of animal-based high-protein diet, which in turn has increased the use of antimicrobials for the overall maintenance of livestock health and productivity (Martin, Thottathil and Newman, 2015). Generally, 73% of all the antimicrobials marketed on the planet are often used in food-producing animals (Van Boeckel et al., 2017). Indiscriminate use of antibiotics in intensive farming has immensely contributed to the emergence of antibiotic resistance in bacteria and the occurrence of antibiotic residues in livestock and in animal-originated food products (Moudgil et al., 2018). The World Health Organization (WHO) reported that antibiotic resistance is one of the most pressing global challenges of the 21st century (World Health Organization, 2024).
Staphylococcus aureus, a Gram-positive, nonspore former, is widely recognized as one of the leading foodborne pathogens in food items such as dairy, bakery, meat, and poultry products (Wang et al., 2017; Kumari and Kulkarni, 2022). This pathogen can cause several infections ranging from localized pyogenic infections, such as skin and soft-tissue infections, to systemic life-threatening infections, such as septicemia, necrotizing fasciitis, infective endocarditis, and toxic shock syndrome. The pathogenicity of staphylococci is mediated by various virulence factors such as cell wall components, toxins, and extracellular enzymes, and the prevalence of antibiotic resistance in staphylococci further complicates the treatment of such infections (Li et al., 2019). Meat and meat products are highly perishable and more prone to microbial attack because of their high moisture content (75%), favorable pH, and plenty of nutrients (Costache et al., 2019). Several meat and meat products are recognized as reservoirs for S. aureus and have been linked to a number of outbreaks across the globe (Igbinosa et al., 2023). During processing, packaging, and storage of meat products, this bacterium has the tendency to infect them and may enter into the food chain. The mesophilic growth condition (optimal temperature 35°C), osmotolerant nature (up to 15% of NaCl concentration), survival in dry state for a long period, and the ability to proliferate in low-water-activity food items (aw = 0.83–0.86) facilitate the survival of S. aureus in adverse climatic conditions (Tapia et al., 2020).
The widespread use of antimicrobials in human and animal therapy has led to the emergence of antibiotic resistance in S. aureus (Silva et al., 2023). Staphylococcus aureus is one of the most common drug-resistant bacterial pathogens that exhibit resistance against a number of antibiotics such as tetracycline, methicillin, oxacillin, kanamycin, gentamicin, streptomycin, and many others. Among S. aureus strains, the occurrence of MDR (multi drug resistance), particularly methicillin-resistant S. aureus (MRSA), in humans as well as in food items of animal origin has risen globally (Fox et al., 2017).
Antimicrobial resistance (AMR) in bacteria is basically mediated by AMR genes that are more commonly present on mobile genetic elements; they can also be present on the chromosomes (Partridge et al., 2018; Robinson et al., 2016). These transferable genes include those encoding penicillinases and cephalosporinases (bla genes), acetyl transferases (aac genes; responsible for alteration in aminoglycoside), erm-gene (that can contribute to antibiotic target modification), methicillin resistance gene (mecA), quinolone/fluoroquinolone resistance gene (qnrA, qnrB, and qnrS), and glycopeptide resistance gene (van genes) (Bitrus et al., 2017; Lerminiaux and Cameron, 2019).
In staphylococci, methicillin resistance is mediated by acquisition of mecA gene (also encodes a low-affinity β-lactam penicillin-binding protein PBP2a). This gene is widely distributed among methicillin-resistant coagulase-negative staphylococci (Argudín and Butaye, 2016; Fowoyo and Ogunbanwo, 2017). Several other AMR genes have also been reported in Staphylococcus isolated from animal sources (Mesbah et al., 2021); Wendlandt et al. (2015) reported 44 different AMR genes in staphylococci from human and animal origin.
Thus, antibiotic residues and drug-resistant bacteria in animal-originated food items pose a direct health risk to consumers, as the drug-resistant bacteria have the potential to cause foodborne diseases that are difficult to treat (Owusu-Doubreh et al., 2023). Therefore, monitoring microbial contamination, their detrimental role, and antibiotic resistance status in bacteria isolated from meat and meat products is of public health importance.
The present study was carried out with the aim of detecting the prevalence of staphylococci in fresh and cooked meat; the antibiotic profiling of the isolates, including the presence of β-lactamase production; and detecting AMR genes in the isolates.
Materials and Methods
Sample preparation
A total of 90 samples comprising raw buffalo minced meat (n = 70) and street meat kebabs (n = 20) were randomly collected from the local retail shops of Aligarh City (Uttar Pradesh, India) in a sterile polythene bag and transferred immediately to the laboratory in an ice box container kept away from direct sunlight. Homogenate was prepared according to the methods described by Anas and Malik (2021).
Isolation and identification of Staphylococcus spp.
For the isolation of Staphylococcus spp., each sample was processed according to the Food and Drug Administration’s standard procedure (Tallent et al., 2016). Aliquots (0.1 mL) of previously prepared homogenates (of meat) were spread onto Baird-Parker agar (BPA) plates supplemented with Egg Yolk Tellurite Emulsion (FD046L, HiMedia, India) and incubated at 37°C for 24 h. Convex, black shiny colonies with narrow white margin surrounded by an opaque zone were considered as Staphylococcus spp. (Tallent et al., 2016). The isolated bacterial strains were primarily characterized based on colony morphology (Tallent et al., 2016), Gram’s reaction, KOH (3%) string test (Halebian et al., 1981), enzymatic reactions, biochemical tests, MALDI-TOF-MS (Bruker Daltonics, Germany), and 16S rRNA sequencing.
Scanning electron microscopy
The morphology of bacterial isolates was analyzed using a scanning electron microscope (JEOL JSM-6510 LV, Japan). Samples were prepared according to Kaláb et al. (2008) by inoculating bacterial strains in nutrient broth followed by incubation at 37°C for 24 h. Aliquots (5 mL) of cultures were pelleted by centrifugation at 12,000 × g for 10 min; the pellets were washed thrice with 1× phosphate-buffered saline (PBS) and prefixed with 2.5% glutaraldehyde overnight at 4°C. Glutaraldehyde was removed by performing three successive washings of the pellet using PBS at 10,000 × g for min; the specimens were dehydrated in a graded series (30, 50, 70, 90, and 100%) of ethanol and resuspended in PBS. Aliquots (5 µL) of bacterial suspension were smeared onto a glass slide and air dried. The specimens were mounted and visualized under a scanning electron microscope to record the morphology of bacterial cells.
Antibiotic susceptibility testing
Antibiotic susceptibility test of bacterial isolates was carried out by the Kirby–Bauer disk diffusion method (Hudzicki, 2009) according to interpretive criteria recommended in the 28th edition of Clinical and Laboratory Standards Institute guidelines M100 (CLSI, 2018). The following antibiotics were tested: methicillin (M30), vancomycin (VA30), sulfadiazine (SZ300), cefaclor (Cj30), nalidixic acid (Na30), gatifloxacin (GAT5), gemifloxacin (GEM5), ertapenem (ETP10), meropenem (MRP10), imipenem (IPM10), cefepime (CPM30), ceftazidime (CAZ30), cefotaxime (CTX30), cefotaxime/clavulanic acid (CEC30/10), aztreonam (AT30), tetracycline (T30), oxacillin (OX5), chloramphenicol (C30), ciprofloxacin (CIP5), kanamycin (K30), and erythromycin (E15) (HiMedia, Mumbai, India). Isolates showing resistance to at least one of the key antibiotics of the three or more antimicrobial classes were designated to possess multidrug resistance (MDR) (Rahman et al. 2020).
The multiple antibiotic resistance index
The multiple antibiotic resistance index (MARI) was derived mathematically as described previously (Blasco et al., 2008).
Detection of β-lactamase production in bacterial isolates
β-lactamase production in bacterial isolates was carried out using iodometric assay as described by Livermore and Brown (2001). Bacterial isolates (grown heavily on agar plate) were suspended in benzyl penicillin solution (6 mg/mL benzyl penicillin in 0.1M phosphate buffer of pH 6) to get a dense suspension in a small tube. The suspension was held at room temperature for 30–60 min, and then, 30 µL of starch solution (1%) and 30 µL of iodine reagent (2% iodine in 53% aqueous potassium iodide) were added in a reaction tube. As a result of the starch–iodine interaction, a blue color develops. On shaking, rapid decolorization of blue color within 5 min indicates the presence of β-lactamase. No change in blue color showed negative result. Escherichia coli RP4 (MTCC-391) and E. coli (ATCC 25922) were used as positive and negative controls, respectively.
Combined disc diffusion assay for phenotypic detection of extended-spectrum beta-lactamase in bacterial isolates
For phenotypic detection of extended-spectrum beta-lactamase (ESBL) in bacterial isolates, the combined disc diffusion assay (CDDA) was performed according to the guidelines of CLSI (2018) using both cefotaxime (30 µg) and ceftazidime (30 µg) alone and in combination with clavulanic acid (10 µg) (CLSI, 2018). Overnight grown culture (0.5 McFarland’s opacity) was spread onto the MHA plates. Discs of cefotaxime (30 µg) alone and cefotaxime in combination with clavulanic acid (30 µg/10 µg) were placed 30 mm apart on MHA plates. Similarly, ceftazidime alone and ceftazidime in combination with clavulanic acid (30 µg/10 µg) were placed on the MHA plates and incubated at 37°C overnight. Isolates showing diameter of inhibition zone ≥5 mm around the combined disc compared with antibiotic disc alone were considered as positive for ESBL production. Positive and negative controls of bacterial strains Klebsiella pneumoniae (ATCC 700603) and E. coli (ATCC 25922), respectively, were used for ESBL production.
Isolation of DNA and polymerase chain reaction-based detection of the antibiotic resistance genes
Bacterial DNA was isolated according to the standard phenol–chloroform method (Sambrook et al., 1989) and used as template for detecting the presence of AMR genes. Polymerase chain reaction (PCR) was performed using the specific primer pairs as listed in Table 1. A 25 µL reaction volume was used for each PCR, containing MgCl2 at 2 mM, dNTPs 150 µM, Taq polymerase 1.5 U, and 10 pM of each primer. PCR amplification was carried out in a thermocycler (Bio ER XP cycler, Germany). The following PCR conditions were used: initial denaturation at 95°C for 2 min, followed by 35 cycles of amplification at 95°C for 30 s, annealing (at 57°C for qnr genes, 60°C for mecA, and 65°C for Sul1) for 45 s, extension at 72°C for 1 min, and followed by a final extension at 72°C for 5 min. An aliquot of 5 µL of the amplified product was run on agarose gel (1.2%) to ascertain the size of amplicon, and images were captured using the gel documentation system (Gel Doc, BioRad, USA).
Primers Used for the Detection of Antibiotic Resistance Genes, Their Annealing Temperature, and Product Size (bp)
AMR, antimicrobial resistance.
Results
Isolation and biochemical characterization of Staphylococcus spp.
A total of 50 Staphylococcus isolates that showed gray-black shiny colonies with an opaque zone on BPA medium were selected. All the isolates gave a positive reaction to the Gram’s stain and a negative reaction to the KOH string test, and the cell shape under a microscope was observed to be spherical. Biochemical characterization of the isolates is given in Table 2. Morphological features of selected isolates were further confirmed using a scanning electron microscope, with cells appearing spherical in shape under low and high resolution (Fig. 1).

Scanning electron micrograph showing the morphology of S. aureus MSA73 under low and high resolution, A: ×5000; B: ×10,000.
Cultural and Biochemical Characterization of Staphylococcus Isolates
*When the cell walls of Gram-negative bacteria are lysed, cellular DNA is released, which makes the mixture viscous (stringy). The formation of a string in 3% KOH within 60 s is a positive test result and an indication that an isolate is a Gram-negative organism.
Antibiotic resistance, susceptibility behavior, and MARI of Staphylococcus isolates
In vitro antibiotic sensitivity assay of the bacterial isolates showed that 82% of Staphylococcus isolates were resistant to aztreonam, followed by nalidixic acid (80%), sulfadiazine (76%), oxacillin (54%), cefepime (36%), ceftazidime (34%), vancomycin and cefaclor (24%), cefotaxime (22%), erythromycin (10%), chloramphenicol (6%), and kanamycin, gatifloxacin, and ertapenem (4%), and least resistance was observed against meropenem and imipenem (2%), whereas resistance was not observed against ciprofloxacin, tetracycline, gemifloxacin, and cefotaxime/clavulanic acid (Fig. 2).
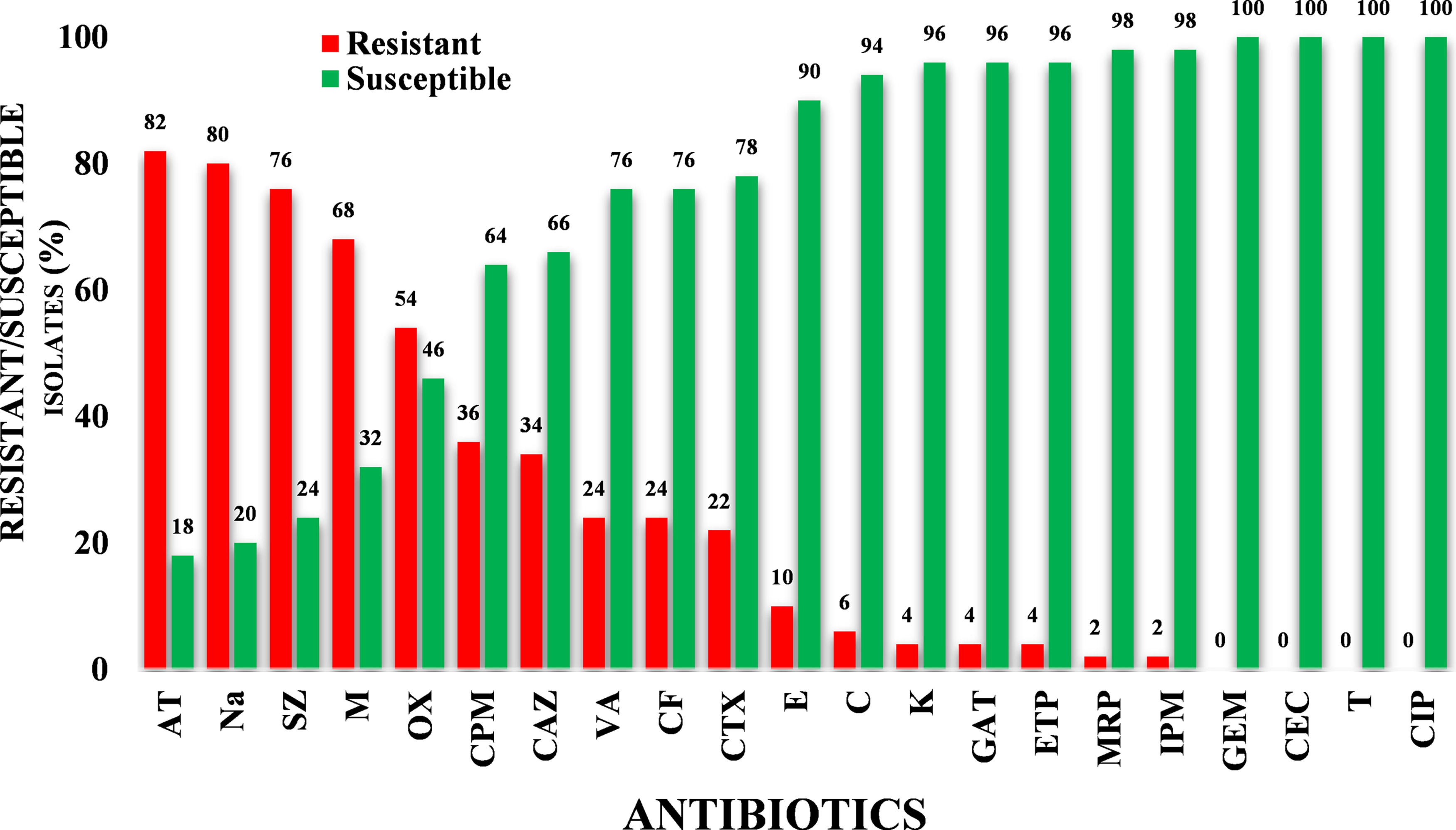
Antimicrobial susceptibility profile of Staphylococcus isolates against tested antibiotics.
The multidrug resistance pattern of each isolate, including the multidrug resistance, was observed against the abovementioned antibiotics. Different resistance patterns among isolates within the same genus indicated the strain diversity. Staphylococcus isolates showed 36 different antibiotic resistance patterns, with 82% of them showing multidrug resistance (Fig. 3). Two percent of the isolates showed the highest MDR pattern against 13 antibiotics of varying classes. It was noticed that the MARI varied among Staphylococcus isolates and ranged between 0.1 and 0.6 (Fig. 4).
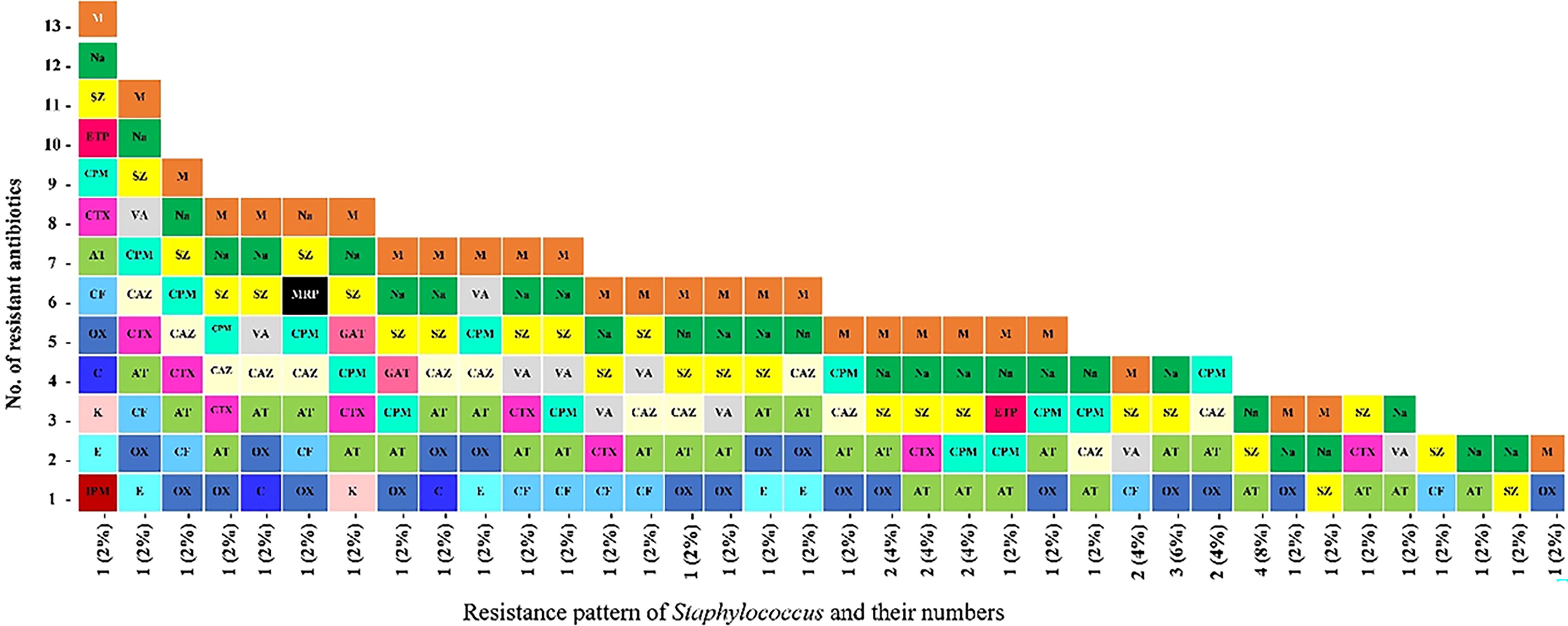
Multidrug resistance pattern of staphylococci isolates from raw and processed meat.
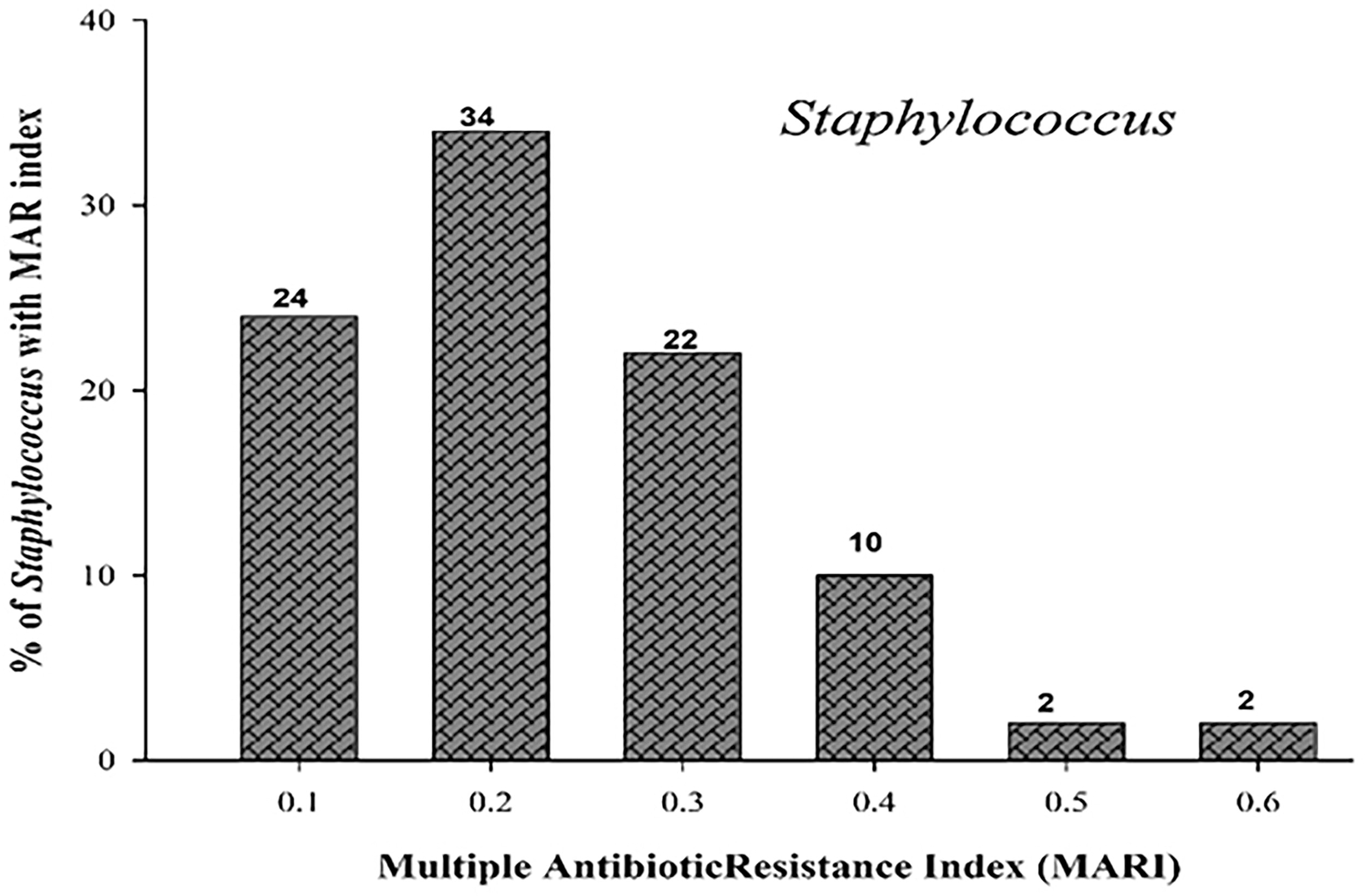
Multiple antibiotic resistance index (MARI) of Staphylococcus isolates.
Phenotypic detection of β-lactamase and ESBL production
To detect ESBL production in Staphylococcus, isolates were primarily subjected to iodometric assay for the detection of β-lactamase production using penicillin as a substrate. The incidence of β-lactamase production was recorded in 48% of the tested isolates (Table 3). Moreover, β-lactamase-producing isolates (24) were subjected to CDDA for the phenotypic detection of ESBL production, and it was found that 50% of them were able to produce ESBL phenotypically (Table 4).
Confirmed β-Lactamase-Producing Staphylococcus Isolates
(+) indicates the hydrolysis of penicillin used as substrate.
Phenotypic Determination of Extended-Spectrum Beta-Lactamase Production in the Bacteria Isolated from Meat and Meat Products
Those isolates having comparative zone diameter >5 mm, with and without clavulanic acid, are assumed as positive for ESBL; R denotes no zone; in this case, difference is calculated by excluding disc size, that is, 6 mm; CTX30 and CAZ30 symbolize cefotaxime and ceftazidime with disc potency 30 µg/mL, whereas CEC30/10 and CAC30/10 symbolize cefotaxime + clavulanic acid and ceftazidime+ clavulanic acid with disc potency 30 + 10 µg/mL each.
PCR-based detection of AMR gene in the bacteria isolated from meat and meat products
All the MDR and β-lactamase-producing Staphylococcus isolates were subjected to PCR for the detection of AMR genes (Table 1). Gel electrophoresis showed amplification of different products corresponding to their respective genes: a 600-bp product for qnrS and a 497-bp product for qnrB were present in 20.8% of the isolates. A 733-bp product corresponding to mecA genes was present in 29.2% of the isolates, whereas a 158-bp product corresponding to Sul 1 gene was observed in 25% of the bacterial isolates (Fig. 5).
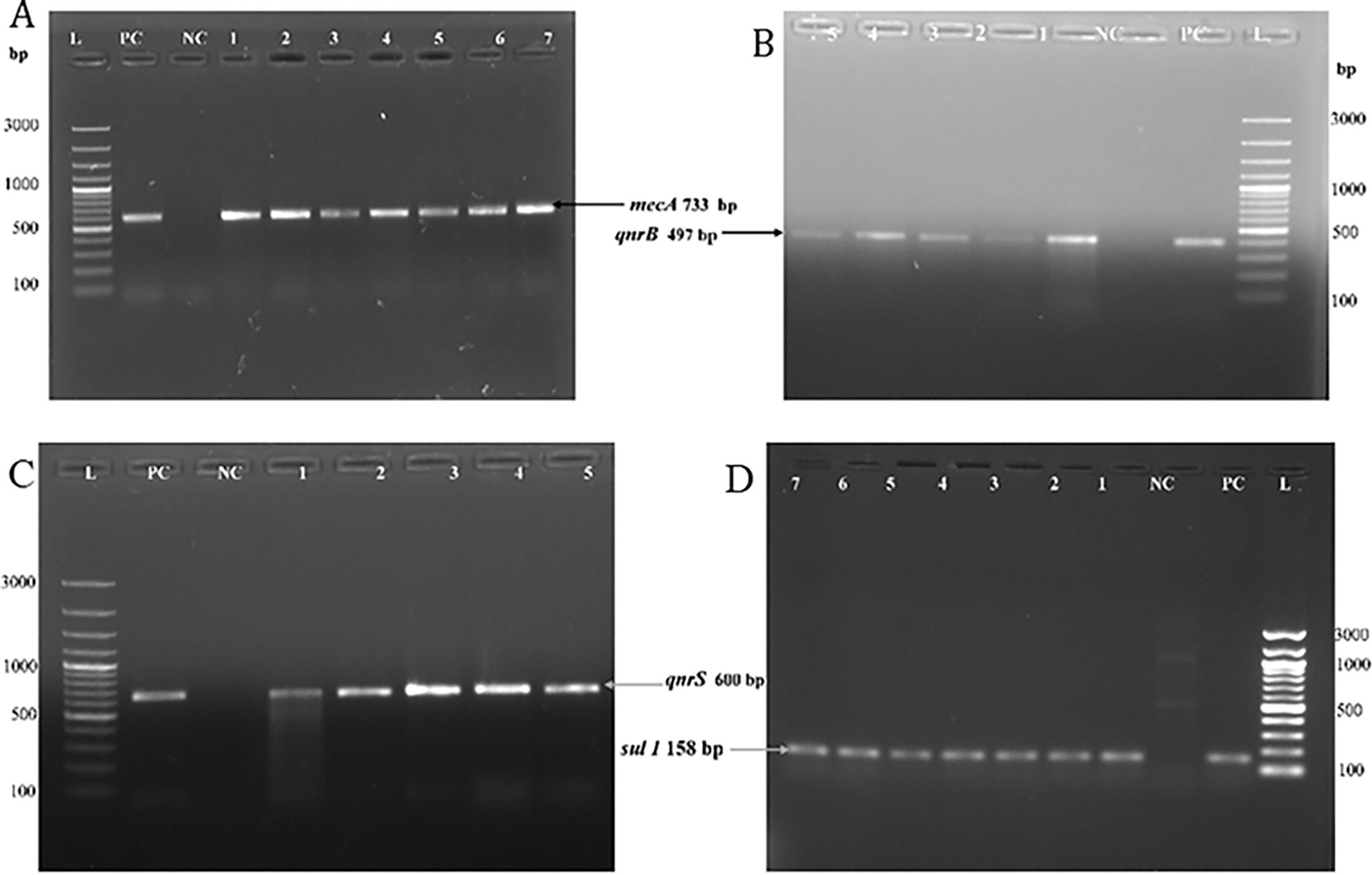
Agarose gel electrophoresis of PCR-amplified products.
Bacterial isolates harboring qnrS and qnrB AMR genes were also frequently resistant to at least one of the different generations of non-β-lactam quinolone antibiotics such as nalidixic acid, ciprofloxacin, gatifloxacin, and gemifloxacin. The mecA and sul 1 genes harboring bacterial isolates showed resistance to sulfadiazine and methicillin. The phenotypic resistance pattern in context to genotypic resistance characteristics of individual isolates is given in Table 5.
Phenotypic and Genotypic Characteristics of MDR and Extended-Spectrum Beta-Lactamase-Producing Isolates
Molecular characterization of bacterial isolates
To ascertain the phylogenetic relationship of the bacterial isolates, the gene corresponding to 16S rRNA was successfully amplified and sequenced using Sanger’s chain termination method. Basic Local Alignment Search Tool analysis of the sequences revealed that the obtained bacterial sequences showed 97–100% identity with the reported sequences of S. aureus. The phylogenetic tree of the isolates was constructed using MEGA software (version 10.2.5). A comparative analysis of 16 s rRNA gene of the strains was done by constructing the maximum likelihood consensus tree with the reference sequences. In each case, the phylogenetic tree was supported by 500 bootstrap values. The phylogenetic relationship among reference strains and isolates is shown in (Fig. 6). MALDI-TOF MS analysis also showed that bacterial isolates matched with S. aureus with a high-confidence identification of score value ranging from 2.20 to 2.48.

Phylogenetic tree based on 16S rDNA sequencing of the Staphylococcus aureus isolates KSA10 (MW644647) and MSA73 (MW644648) was constructed using maximum likelihood method in MEGA version 10.2.5. The bootstrap consensus tree inferred from 500 replicates is taken to represent the evolutionary history of the taxa analyzed.
Discussion
As the global human population is rising, the demand for animal protein is also rising, which is facilitated by the intensive industrial farming throughout the world (Henchion et al., 2014). Even so, intensive livestock farming depends on the daily use of antimicrobials to sustain health and productivity of farms (Lhermie et al., 2018), which ultimately leads to the emergence and spread of AMR (Pokharel and Karna, 2022; Tiseo et al., 2020).
Foodstuffs play a significant role in the spread of commensal as well as foodborne pathogens. It occurs during the processing of raw products as well as cross-contamination and recontamination at different stages of the food supply chain (MacDonald et al., 2015). Food contamination with antimicrobial-resistant bacteria may pose a serious threat to public health because AMR may be transferred to other clinically important bacteria (Collignon et al., 2016).
S. aureus is a non-spore-former, Gram-positive facultative anaerobic bacterium commonly found in the outdoor environment (i.e., air, water, soil, and dust) and in humans and other animals as natural microflora, predominantly present on skin, hair, and nasal passage (Anas et al., 2021; Madigan et al., 2012). Through contamination, during slaughtering, processing, storage, and marketing, S. aureus can enter into the food chain and may cause several human illnesses (Rong et al., 2017). Staphylococcal food poisoning (SFP) is known to be one of the world’s most serious foodborne diseases caused by intake of highly heat-stable enterotoxins produced in foods (Bianchi et al., 2022).
In the present study, a total of 50 staphylococci isolates were obtained from meat and meat products and were tentatively identified based on their cultural, morphological, and biochemical characteristics as per standard protocols of Cappuccino and Sherman (1995). Isolates with the highest level of resistance, as indicated by in vitro testing, MDR pattern, and the presence of AMR gene(s), were further characterized by 16S rRNA sequencing and MALDI-TOF MS analysis and identified as S. aureus. Morphological characteristics of these bacterial isolates were confirmed by scanning electron microscopy, which showed spherical-shaped cells arranged in clusters (Fig. 1), a characteristic feature of S. aureus. Bantawa et al. (2018) isolated S. aureus from raw buffalo meat. Das et al. (2019) predominately isolated and identified S. aureus from bovine carcasses. Thwala et al. (2021) reported the presence of Staphylococcus in meat and meat products. The presence of pathogenic bacteria in meat and meat products with varying incidence has also been globally reported (Mat et al., 2017; Saif et al., 2019; Omoruyi and Ajayi, 2021).
In the present study, Staphylococcus isolates showed widespread resistance among β-lactam antibiotics such as aztreonam (82%), methicillin (68%), oxacillin (54%), cefepime (36%), ceftazidime (34%), cefaclor (24%), cefotaxime (22%), ertapenem (4%), and meropenem and imipenem (2%). The growing resistance against the third- and fourth-generation cephalosporins and carbapenems in the present study is worrisome since these antibiotics are not commonly used in animal medicine. Hence, it is clear that widespread application of cephalosporins and carbapenems in clinical practice along with many other factors plays a crucial role in the dissemination of antibiotic resistance. Also, it could be related to the production of ESBL enzymes (Jaja et al., 2020). Nomura et al. (2020) reported that resistance to oxacillin in S. aureus is contributed by staphylococcal β-lactamase that functions as ESBL.
Among non-β-lactam antibiotics, the most widespread resistance was observed against nalidixic acid (80%), sulfadiazine (76%), vancomycin (24%), erythromycin (10%), chloramphenicol (6%), and kanamycin and gatifloxacin (4%), which reveals the growing problems of AMR in Staphylococcus obtained from food-producing animals. In a study conducted by Osman et al. (2017), 34.8% of staphylococci isolates were found to be resistant to methicillin, 26% to erythromycin, 4.3% to chloramphenicol, 30.5% to ciprofloxacin, and 38.7% to vancomycin. Also, they reported MDR in 69.6% (16/23) of their isolates with 15 types of drug resistance pattern (Osman et al., 2017). Similarly, Wang et al. (2017) cultured S. aureus from raw meat, in which 52.1% of the isolates exhibited resistance to erythromycin, 17.4% to ciprofloxacin, and 12.0% to chloramphenicol; moreover, 57.5% of the isolates exhibited an MDR pattern. Naas et al. (2019) conducted a study on the occurrence, characterization, and antibiogram of S. aureus in meat and meat products in Libyan retail markets and found resistance to cefotaxime (100%), erythromycin and kanamycin (50%), and methicillin (16%). Bantawa et al. (2019) reported that S. aureus cultured from different meat samples in Eastern Nepal were resistant to nalidixic acid (17%) and cefotaxime (13%). In our study, 82% of the Staphylococcus isolates showed an MDR pattern that is higher than that of the other MDR Staphylococcus isolates obtained from diverse environments and other animal-originated food items. In a study conducted by Saud et al. (2019), 21.9% of S. aureus isolates recovered from buffalo meat were reported to be MDR.
In the present study, the occurrence of bacteria with a high MARI value (>0.2) is an indication of high risk of contamination in the food chain. The higher risk is primarily due to the injudicious use and therefore greater exposure of humans, veterinary, poultry, and livestock to the antimicrobials. Jaja et al. (2020) reported that the MARI of S. aureus ranged from 0.3 to 0.8 in meat. Savariraj et al. (2019) and Mahros et al. (2021) also reported an MARI of >0. 2 in S. aureus isolated from meat and meat products.
β-lactamase production is a crucial cause of antibiotic resistance in bacteria. In the present study, in vitro iodometric assay and CDDA revealed that 48% of the Staphylococcus isolates produced β-lactamase, whereas 24% of isolates produced ESBL. Agu et al. (2020) detected β-lactamase in Staphylococcus (62.5%) isolates from meat and meat-based food items. Pugazhendhi et al. (2020) reported β-lactamase production through iodometric assay in all strains of S. aureus isolated from poultry. Schaumburg et al. (2014) reported ESBL production in Staphylococcus (3%) isolates from chicken meat. Various reports showed that S. aureus isolated from different sources had β-lactamase production and MDR (Pugazhendhi et al., 2020; Şanlıbaba, 2022).
The presence of qnrS and qnrB genes in bacterial isolates is responsible for acquiring resistance against quinolone antibiotics. In the present study, qnrS and qnrB genes were detected in 20.8% of the Staphylococcus isolates. Abdu and Mirabeau (2019) reported the qnrB gene in 9% of the clinically isolated MDR S. aureus isolates. Quinolone is a broad-spectrum antibiotic and widely used in the treatment of disease, especially urinary tract infections, with few side effects (Shahcheraghi et al., 2013). The presence of plasmid-mediated quinolone resistance (PMQR) genes qnrS and qnrB in bacterial isolates facilitates the resistance to quinolone antibiotics, such as nalidixic acid, gemifloxacin, ciprofloxacin, and gatifloxacin, and the same was phenotypically detected in bacterial isolates. These are highly significant antimicrobials and have been proven to be effective in reducing infections caused by Gram-negative bacilli in immunosuppressed patients. This resistance phenotype is unusual in food, and imports can be a source of PMQR genes for humans (Høg et al., 2019). Most of the AMR genes detected in our isolates were previously identified on mobile genetic elements and could be transferred to the pathogens.
The methicillin resistance mecA gene and sulfadiazine resistance sul 1 gene were detected in 29.2% and 25% of the staphylococci isolates, respectively. Fijałkowski et al. (2016) reported that the mecA gene was present in 4% of the staphylococci isolates from ready-to-eat meat. Bokharaei et al. (2020) found that 15.4% of S. aureus isolates from Iranian food were positive for the mecA gene. Parvin et al. (2021) found that 43.5% S. aureus isolates from frozen meat were positive for the mecA gene. Shahid et al. (2021) also detected the mecA gene in 12.20% of Staphylococcus isolates from food processing units. Haubert et al. (2017) reported sulfadiazine-resistant sul 1 and sul 2 genes along with the mecA gene in S. aureus isolates from subclinical bovine mastitis in Brazil. The presence of the mecA gene in Staphylococcus is considered to be the cause of resistance to other β-lactam antibiotics in the bacteria of same groups (Börjesson et al., 2009). A new penicillin-binding protein (PBP-2a) that is encoded by the mecA gene in S. aureus has a low affinity toward methicillin, thus allowing the growth of the cell, even in the presence of inhibitory concentrations of antibiotics. These PBPs provide resistance and decrease the ability of β-lactam antibiotics to act on biosynthesis of the bacterial cell wall (Peacock and Paterson, 2015).
Conclusion
The findings of the present study indicate that there is very high prevalence of multidrug resistance in staphylococci isolated from raw buffalo meat and street meat products. The presence of such MDR strains in raw and processed meat products has the potential to spread antibiotic resistance to other bacteria through human consumption of meat, thereby posing a serious threat to public health. The resistant behavior of Staphylococcus strains against quinolones, β-lactams, and sulfadiazine antibiotics is primarily due to the presence and expression of qnrB, qnrS, mecA, and sul 1 genes, and these genes can be easily transferred to other bacteria via horizontal gene transfer. The present findings suggested for a strict monitoring and implementation of effective biosafety and hygienic measures in the whole food chain. Appropriate selection of antibiotics, right dosages, increased use of vaccination for vaccine-preventable infectious diseases, and judicious use of antibiotics in food animals, as well as regular monitoring of antibiotic resistance and susceptibility profiling, are some of the immediate measures to be taken for the prevention of antibiotic resistance.
Footnotes
Acknowledgment
M.A. is thankful to the University Grants Commission (UGC), New Delhi, India, for non-NET fellowship.
Authors’ Contributions
M.A.: Conceptualization, methodology, investigation, and writing—original draft. S.A.L.: Methodology, formal analysis, data curation, and writing—original draft. A.M.: Conceptualization, resources, supervision, data curation, and editing. J.A.: Formal analysis, data curation, and editing. All authors have read and agreed to the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.