
Abstract
Vibrio vulnificus, both types A and B, in tilapia fish causes several diseases in humans—septicemia, gastroenteritis, and wound infection—that primarily occur due to the consumption of raw or undercooked fish. Using fresh tilapia fish samples collected from Riyadh market in Saudi Arabia, this study attempted to phenotypically detect V. vulnificus using specific media, such as thiosulphate citrate bile salt sucrose agar, CHROMagar™ Vibrio, and cellobiose polymyxin colistin agar, while Api 20NE was employed. Molecular detection was conducted using 16S rRNA gene sequencing, to characterize the clinical (genotype B) and environmental strains (genotype A) of V. vulnificus. Virulence genes, such as hemolysin A (VvhA) and ViuB which encodes vulnibactin siderophores, were employed to detect both strains, in addition to capsular polysaccharide (CPS) and allele 1 genes. Virulence-correlated (vcg) genes vcgC and vcgE were also considered. The multidrug resistance of both strains to different antibiotics was tested, while whole-cell protein profiles were used to examine their differences based on Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis. The tilapia fish contained type A and B of V. vulnificus at a ratio of 12% and 4%, respectively. Generation times of the clinical and environmental strains were 14.45 min and 12.04 min, respectively. Analysis of the 16S rRNA, VvhA, CPS, ViuB, vcgC, and vcgE in type A and B revealed that both strains comprised all virulent genes in different ratios. Raw tilapia fish contained V. vulnificus type A and B, so care in handling is essential or during cooking these kinds of fish.
Introduction
Vibrio vulnificus is a virulent, Gram-negative, halophilic, nonspore-forming, rod shaped, motile bacterium found in seafood, particularly raw oysters, from marine and estuarine environments worldwide (Vickery et al., 2007). In particular, tilapia fish have been found to be contaminated with V. vulnificus, with the handling and method of cooking tilapia, as well as its source and individual health status, considered possible risk factors (Tanveer et al., 2024). Carmona-Salido et al. (2021), were isolated zoonotic V. vulnificus strains from vibriosis outbreaks occurring on eastern Mediterranean tilapia farms between 2016 and 2019.
Raw or undercooked seafood consumption and open wounds exposed to contaminated water have been found to be the main causes of infection with V. vulnificus (Grau et al., 2008). Previous studies have also observed that the consumption of contaminated oysters or seafood results in high mortality rates due to septicemia caused by V. vulnificus (Oliver, 2005; Jones and Oliver, 2009), especially in people infection with chronic liver disease, diabetes, and/or other immunocompromising conditions (Hsueh et al., 2004).
The commonly found biotype one of V. vulnificus, found in shellfish and in the intestinal contents of fish, infects humans either through the ingestion of raw or undercooked shellfish or by wound exposure to the bacterium (Strom and Paranjpye, 2000). Meanwhile, biotype two of V. vulnificus commonly infects marine vertebrates and has been reported to cause infections in humans (Veenstra et al., 1992). Furthermore, biotype three, which has been found to cause wound infection and bacteremia in Israel due to incorrect handling of tilapia, was identified by Bisharat et al. (1999). Notably, Kim et al. (2017) observed that biotype three strains are less pathogenic than biotype one strains.
V. vulnificus harbors several virulence factors, such as vulnibactin siderophores (ViuB), hemolysin A (VvhA), capsular polysaccharides (CPS), lipopolysaccharides, exotoxins, pili, and flagella (Strom and Paranjpye, 2000; Gulig et al., 2005). In this regard, 16S rRNA, CPS, and virulence-correlated (vcg) genes have been found to be highly correlated biomarkers for V. vulnificus (Nilsson et al., 2003; Rosche et al., 2005; Chatzidaki-Livanis et al., 2006). While the isolates of biotypes vcgC and vcgE can be differentiated using vcg genes (Rosche et al., 2005), the environmental (type A) and clinical (type B) isolates of V. vulnificus can be detected using 16S rDNA sequences (Nilsson et al., 2003). Furthermore, differentiation between clinical and environmental isolates can be conducted using CPS genes (Chatzidaki-Livanis et al., 2006).
Infections related to this pathogen, which leads to primary septicemia, have a fatality rate similar to Biosafety Level 3 and 4 (BSL 3 and 4) pathogens, which cause diseases such as anthrax, bubonic plague, Ebola, and Marburg fever. In particular, V. vulnificus largely affects patients suffering from diabetes, liver disease, and immune disorders, disproportionately affecting males (∼85% of cases) and older patients (>40 years) (Baker-Austin and Oliver, 2018).
Quantitatively estimating the generation time of bacteria is integral to understanding their population dynamics. However, it is extremely difficult to determine the generation time of most bacteria in the natural environment, with only a few estimates currently available (Gibbons and Kapsimalis, 1967). To address this gap in the literature and understand its related dynamics, we conducted in vitro estimations of the generation time of both the clinical and environmental strains of V. vulnificus under optimum conditions using the nutrient-rich media of tryptic soya broth.
In Saudi Arabia, fresh tilapia fish are sold in the markets of Riyadh. This study attempts to elucidate the prevalence of V. vulnificus, considering both its clinical and environmental strains, in tilapia by focusing on its phenotypic and genotypic detection and identification using different virulence factors. Protein profiles were considered to distinguish between the clinical and environmental strains on SDS-PAGE. Furthermore, to determine the antimicrobial resistance of V. vulnificus isolated from different tilapia fish samples. The generation time was also determined to explain the variance in time taken for type A & B cells to reproduce under optimization growth conditions. Ultimately, this study raises awareness about the risks of consuming raw or undercooked fish and seafood.
Materials and Methods
Tilapia fish sample (whole fish)
Fresh fish samples (n = 50) were collected from markets located in Riyadh, Saudi Arabia, and then transferred in an icebox to the Laboratory of Food Microbiology, College of Food and Agricultural Science, King Saud University.
Media used
To isolate V. vulnificus from the tilapia fish samples (whole fish), they were first subjected to pre-enrichment in alkaline peptone water (Oxoid, Code: CM1028) for 24 h at 37°C, after which they were streaked onto thiosulfate-citrate-bile salts-sucrose (TCBS) agar (Oxoid, Code: CM0333) (Colwell, 1984) for 24 h at 37°C. Subsequently, green colonies were picked from the TCBS agar, streaked onto CHROMagar™ Vibrio, and then incubated at 37°C for 24 h. The green-blue to turquoise-colored colonies were considered V. vulnificus, while the mauve-colored colonies were identified as V. parahaemolyticus . When streaked on cellobiose polymyxin colistin (CPC) agar, V. vulnificus produced flat yellow colonies with opaque centers and translucent edges.
Identification of strains using the API 20NE system
The API 20NE (Ref. 20050, bioMérieux, France) system helps identify non-fastidious, non-enteric, Gram-negative rods. For this study, the test strips were first inoculated and incubated according to the instructions of the kits. Next, colonies of suspected Vibrio spp. isolates were suspended in sterilized saline solution (0.85% NaCl) for inoculation into eight conventional substrates, as well as in AUX medium for inoculation into 12 assimilation cupules. In addition, an external test for oxidase detection was conducted. The seven-digit profile number obtained using the API 20NE system was converted into a unique identifier using APILAB software (version 3.3.3). Notably, both API 20NE and API 20E can be found on the same website. The API 20NE results showed a probability of ≥80%, which was considered acceptable for the current study (bioMérieux).
DNA extraction and detection of V. vulnificus and V. parahaemolyticus genes
DNA was extracted from the colonies of V. vulnificus and V. parahaemolyticus grown in brain heart infusion agar (Oxoid, Code: CM1136) + 3% NaCl, and then incubated for 24 h at 37°C. Using the method proposed by Gulig et al., (2005), along with some modifications, one colony was selected, suspended in 100 µL sterilized distilled water, and then boiled for 10 mins. Subsequently, it was cooled in ice sediment at max. speed 20,800× g for 1 min. This suspension was then used as a template for the PCR assay. As shows in Table 1. gene specific-primers were designed for the 16S rRNA type A (environmental strain) suggested by Han and Ge (2010) and 16S rRNA type B (clinical strain) as (Warner and Oliver, 2008), with modified to G nucleotide instead of C by owns. VvhA gene was designed as according to Brasher et al., (1998), ViuB gene as Panicker et al., (2004), CPS allele1 as Han and Ge (2010), vcgC as modified by (Warner and Oliver, 2008) and vcgE as modified Rosche et al. (2005). While for V. parahaemolyticus toxR primer was screened as Kim et al. (2017). The amplification process for V. vulnificus and V. parahaemolyticus genes involved incubating the suspensions at 94°C for 5 min, followed by 35 cycles at 94°C for 1 min, annealing at 62°C for 30 s, and elongation at 72°C for 30 s. Subsequently, a final extension was conducted at 72°C for 10 min. The amplified PCR products were then separated on 1% agarose gel using ethidium bromide at 100 v for approximately 1 h. Finally, DNA bands were observed under ultraviolet light. The sizes of the DNAs of V. vulnificus and V. parahaemolyticus genes were compared against the DNA ladder run presented in Table 1. The positive isolates were then sequenced, while GenBank and the basic local alignment search tool (BLAST) program were employed to ensure that the proposed primers were consistent with the target species.
PCR Conditions for Detecting V. vulnificus Using Species-Specific and Virulence Genes

Confirming the identification of V. vulnificus and V. parahaemolyticus by sequencing
To confirm the identity of the amplified products, the PCR products for different genes—16S rRNA A and B, VvhA, ViuB, CPS allele1, vcgC, and vcgE were sequenced. To confirm the identify V. parahaemolyticus, the PCR products for toxR gene were also sequenced. For this purpose, the PCR products were first purified and labeled using commercial kits according to the manufacturer’s instructions (AxyPrep™ PCR clean-up kit, Axygen®, NY, USA; BigDye Terminator v3.1 cycle sequencing kit, Applied Biosystems, CA, USA; BigDye X Terminator purification kit, Applied Biosystems, CA, USA), as previously described by Al-Shabanah et al. (2013). The samples were then sequenced using an automatic ABI 3500 genetic analyzer (Applied Biosystems, USA). Nucleotide sequences of 100 bp (for each gene) were further compared with the GenBank database using the BLAST, which was developed by the National Center for Biotechnology Information, USA.
Antibiotic sensitivity test
In accordance with the guidelines of the Clinical and Laboratory Standards Institute (CLSI) (2014), disc diffusion method was implemented to detect the antimicrobial susceptibility of the isolates. Seventeen antimicrobial agents (Becton, Dickinson, and Company, Franklin Lakes, NJ, USA) were used in this study and belonged to different classes of antibiotics as shows in Table 2. Müller-Hinton agar plates were employed to test the different antibiotics after suspending the isolates in a saline solution of 0.85% NaCl. Each isolate was categorized as susceptible, intermediate, or resistant based on the diameter (mm) of the zone of inhibition around the antibiotic disk, in compliance with CLSI (2014) recommendations.
Antimicrobial Agents Tested Belonged to Different Classes of Antibiotic

Biofilm synthesis in V. vulnificus strains cultured on Congo red agar
Vibrio vulnificus isolates were tested for the production of biofilm on Congo red agar (CRA) medium, with the color that the colonies produced when cultured on CRA used for quantitative determination. V. vulnificus isolates were streaked onto brain heart infusion agar and supplemented with sucrose and Congo red stain as per the following measurements: 57 g/l BHIA (Oxoid, CM1136), 30 g/l sucrose, and 0.8 g/L CRA were aerobically incubated at 37°C for 24 h. Subsequently, biofilm produced by the V. vulnificus isolates was compared to that produced by V. parahaemolyticus ATCC 17802 (a non-biofilm-producing negative control). The isolates that formed intensive black, black, or reddish-black colonies with rough, dry, and crystalline consistency were considered normal biofilm producers. In contrast, those producing smooth red or Bordeaux red-colored colonies were classified as nonproducers of biofilm.
Wolfe test
Bacterial adherence to glass surface can be detected by adding some modifications to the method proposed by Wolfe et al. (2004). For this purpose, 1 L of the following media containing 5 g of bacto-tryptone, 3 g of yeast extract, and 3 mL of glycerol in 686 mL tap water, 300 mL distilled water + 3% NaCl were prepared and distributed into test tube (10 mL). After sterilizing the tubes and cooling them down to 45°C–50°C, two of them were inoculated with the clinical strain, while the other two tubes were inoculated with the environmental. Both strains comprised 24-h-old actively growing cultures cultivated in tryptic soy broth. One tube without any bacteria was used as the control. All tubes were incubated at 37°C for 24 h without shaking. Following a 15-min staining period with 1% (w/v) crystal violet, all the glass tubes were cleaned with distilled water before further testing. The strains that produced a purple pellicule on the cultures’ surfaces were identified as glass-biofilm positives.
Preparation of the cell extract
Overnight cultures of the identified colonies of V. vulnificus types A and B were grown in brain heart infusion (Oxoid, Code: CM1135) supplemented with 3% NaCl at 37°C. The cells were collected through centrifugation, after which 250 mg of cells was suspended in 100 μL TES buffer (50 mM tris HCl, pH 8, 1 mM EDTA, 25% sucrose). Ten microliters of lysozyme (50 mg/mL) were added to the cells suspended in the TES buffer and incubated at 37°C for 30 min. Following this, 10 microliters of 20% SDS was added and the contents were mixed until the solution became clearly visible. The contents were stored at −20°C for 1 to 2 d (Yehia, 2014). Subsequently, 25 microliter of the mixture were loaded on SDS-PAGE. Electrophoresis was performed at 25°C in a vertical tank apparatus using a constant-voltage power supply until the bromophenol blue tracking dye reached the bottom of the gel. Notably, the gel was stained with Coomassie Brilliant Blue R-250 for 18 h at room temperature. Next, gel destaining was performed by continuous agitation until obvious bands of proteins appeared. Whole-cell protein profiles of presumptive Vibrio isolates were obtained to compare types A and B based on the protein marker.
Determination of doubling times (τ) for V. vulnificus
By introducing some modifications to the methodology proposed by Dryselius et al. (2008). V. vulnificus strains were streaked on tryptic soya agar and incubated at 37°C for 24 h. One colony of each strain was picked and inoculated in 10 mL of tryptic soya broth medium and incubated at 37°C for 24 h. Five mL of overnight culture of V. vulnificus both strains (A and B) were inoculated into 500 mL flask containing 100 mL of tryptic soya broth supplementing with 3% NaCl and incubated at 37°C in water bath shaker (GFL Incubation/Inactivation Bath 1103, GmbH Landsberger Str. 245, 12623 Berlin, Germany) at 100 rpm. Bacterial cultures were sampled at zero time and then every 60 min and diluted in series, after which they were plated onto tryptic soya agar plates for colony count. Doubling times were determined for mid-exponential-phase cultures by estimating the change in colony count considering a 1-h time span by first calculating the number of generations (n) as follows:
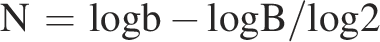
Here, b and B refer to the number of colony-forming unit at the end and at the beginning of the 1 h time interval, respectively. The doubling time (τ) was obtained by dividing the time interval (t) by the number of generations (n), which can be expressed as follows:
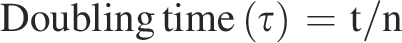
Statistical analysis
Correlations between the virulence genes (VvhA, ViuB, CPS, vcgC, and vcgE) and 16S rRNA (A and B) were analyzed using chi-square or Fisher’s exact tests. All analyses were conducted using SAS (SAS Institute Inc., Cary, NC) software packages.
Results and Discussion
Isolation of V. vulnificus in specific media
Green colonies formed on TCBS agar were believed to be composed of either V. parahaemolyticus or V. vulnificus. Meanwhile, in the case of CHROMagar™ Vibrio, mauve colonies were considered as pertaining to V. parahaemolyticus, and blue colonies were related to V. vulnificus. Furthermore, V. vulnificus appeared on CPC agar as a yellow colony (Fig. 1). Notably, most V. parahaemolyticus strains did not grow on CPC agar. Carmona-Salido et al. (2021) recovered V. vulnificus from Moribund tilapia. In recent years, with the fast development of the tilapia farming industry, various infectious diseases have been reported (Haenen et al., 2023). Janampa-Sarmiento et al. (2024) reported for the first-time isolation of pathogenic V. vulnificus from farmed Sorubim (Pseudoplatystoma sp.).

V. vulnificus and V. parahaemolyticus on different media.
Identification of V. vulnificus using API 20 NE
Based on the biochemical characterization results obtained using API 20NE (Table 3), the following suspected colonies of V. vulnificus and V. parahaemolyticus were identified: six strains were found to be V. vulnificus type A (50%), with their main profiles bearing the seven-digit number 7770645; 2 strains pertained to V. vulnificus type B (16.66%), with the two main profiles related to the seven-digit numbers 7470005 and 5071045; and the remaining four strains were V. parahaemolyticus (33.33%), with a profile pertaining to the seven-digit number 7117444.Biochemical tests such as enzymatic detection and carbohydrate fermentation tests are commonly used to confirm the presence of V. vulnificus based on its characteristic metabolic activities (Loo et al., 2022).
Identification of Bacteria Isolated from Tilapia Fish Samples Using the API 20NE Method

Verifying the identification of V. vulnificus using a PCR test
To further confirm that the colonies identified using API 20 NE pertained to V. vulnificus and V. parahaemolyticus, PCR was employed for their detection using 16S rRNA (A and B) and toxR genes, respectively. As shown in Figure 2, PCR amplicons targeting the species-specific toxR gene (368 bp) were observed for V. parahaemolyticus, while those targeting 16S rRNA (839 bp) appeared for V. vulnificus type A and B. These results verify that this study employed appropriate methods to identify these bacteria. Notably, previous studies (Kim et al., 2017) have also employed toxR-targeted PCR for the identification of V. parahaemolyticus.

Amplification of the toxR and 16S rRNA genes of V. parahaemolyticus and V. vulnificus, respectively, through PCR. Lane 1 indicates molecular weight (MOLEQULE-ON 100 BP CAT NO.: DM-M-002-500), Lane 2 refers to the control strain V. parahaemolyticus ATCC 17802, Lanes 3–7 pertain to the isolates of V. parahaemolyticus (368 bp), and Lanes 8–11 relate to the 16S rRNA gene of Vibrio vulnificus (839 bp).
Colonies of V. vulnificus under UV light
V. vulnificus on CHROM™ Vibrio medium produced colonies that were either magenta in color or exhibited a blue center with a blue fluorescence edge under UV light (365 nm). Other Vibrio spp. produced colonies whose colors ranged from colorless to olive, but they did not fluoresce, as shown in Figure 3.
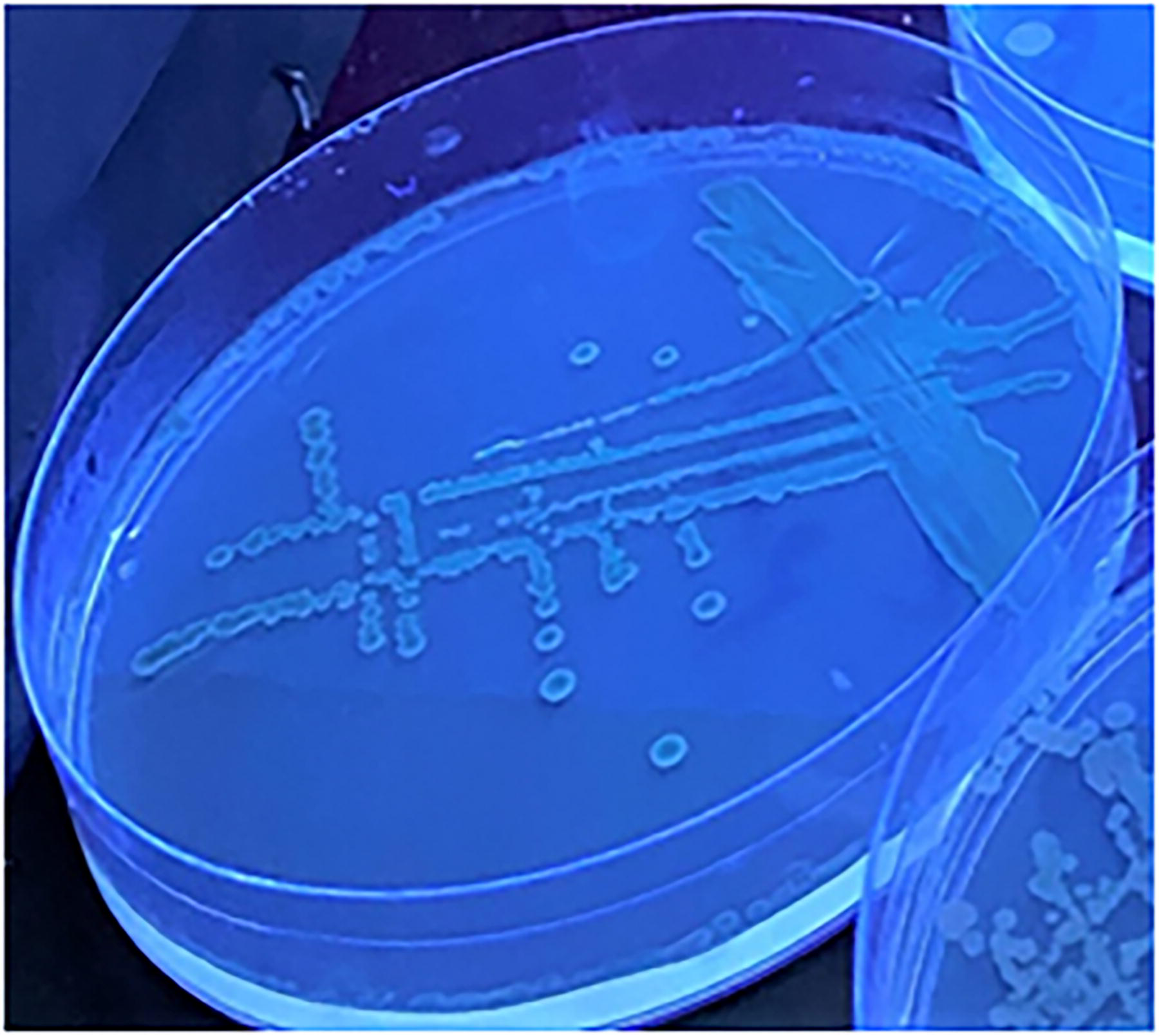
Vibrio vulnificus colonies growing on CHROM™ Vibrio under UV light, after aerobic incubation for 24 h at 37°C.
Biofilm production and adherence to glass
Microorganism communities attached to living or abiotic surfaces form biofilms, which offer extreme resistance to antibiotics, environmental conditions (e.g., pH, temperature, high pressure, high salinity, radiation), and malnutrition, thus acting as protective clothing for microorganisms (Yin et al., 2019). In this study, investigations into Vibrio’s biofilm were conducted using phenotypic methods. For this purpose, the CRA test developed by (Freeman et al., 1989) for Staphylococcus sp., based on the subculture of bacterial strains on BHIA, was supplemented with sucrose and Congo red dye. As shown in Figure 4, both strains of V. vulnificus formed a black-colored colony on CRA, indicating the production of slime, which means that both are biofilm-forming strains capable of food spoilage and endangering the health of consumers. In contrast, V. parahaemolyticus exhibited white colonies on CRA and did not produce any slime. Notably, in most cases, biofilm formation is considered harmful (Coughlan et al., 2016).

Slime production on Congo red agar media: Vibrio parahaemolyticus ATCC 17802
In the context of food sanitation, biofilm formation on abiotic surfaces is considered a concerning issue. For instance, the biofilm matrix formed by exopolysaccharide glycocalyx is extremely resistant to sanitizers, since the biofilm environment stresses the free cells (Wong et al., 2002). With regard to the food industry, foodborne pathogens that form biofilms have been found to result in food spoilage, thus compromising the health of consumers (Galiè et al., 2018) and causing food hygiene problems (Dewanti and Wong, 1995). Furthermore, in hospitals, biofilms of bacteria found on medical devices or on patient tissues result in persistent infections (Han and Ge, 2010). Some marine Vibrio species, such as V. alginolyticus (Kogure et al., 1998), V. cholerae (Hood and Winter, 2006), and V. harveyi (Karunasagar et al., 1996), have also been found to form biofilms. In this regard, a comprehensive examination of the adherence of pathogenic Vibrio to biotic surfaces could help explain their pathogenesis in the host. For instance, Lopez-Cortes et al. (1999) reported on the adhesion of pathogenic marine bacteria to seafood.
Comparison between different genes in both clinical and environmental strains
As shown in Table 4, 16S rRNA type A was found in the environmental isolates, while type B was found in the clinical isolates. Although some clinical isolates contained 16S rRNA type A and vice versa, these could neither be detected when performing DNA sequencing nor did they appear in the migration of the whole protein profile on SDS-PAGE, as explained later in the section. Meanwhile, some isolates exhibited a nontypeable type, which did not pertain to any specific amplicon of 839 bp for either 16S rRNA type A or B. Furthermore, V. vulnificus hemolysin A (VvhA) and CPS allele 1 were found in both environmental and clinical isolates, as was the case with vulnibactin siderophores encoded by viuB, which contributes to iron uptake, and the virulence-correlated genes (vcgC and vcgE).
Comparison of Virulent Factors in Clinical and Environmental Isolates of V. vulnificus

+++ commonly present gene in V. vulnificus
+++/− gene of V. vulnificus that is commonly present but sometimes excluded.
−/+ gene of V. vulnificus that is sometimes excluded but included at other times.
Along these lines, Wanida Mala et al. (2014) reported the predominance of 16S rDNA type B in both clinical (67%) and environmental (67%) isolates, while nontypeable types were detected only in the latter.
They also found that the V. vulnificus strain ATCC 27562 included 16S rDNA type A. Meanwhile, Nilsson et al. (2003) identified the virulence and sources of the types of 16S rDNA.
The hemolytic and cytotoxicity activities of the hemolysin A (VvhA) gene of V. vulnificus, which encodes VvhA, have been thoroughly examined in previous studies (Wright and Morris, 1991; Gulig et al., 2005) to establish that VvhA is a species-specific gene marker for detecting V. vulnificus (Han and Ge, 2010). Studies have also detected the ViuB gene, which is integral for iron acquisition, in both clinical and environmental strains of V. vulnificus (Litwin et al., 1996; Panicker et al., 2004). ViuB was detected in one-third and 50% of the clinical and environmental isolates of V. vulnificus, respectively. While CPS allele 1 achieved a ratio of 75% for environmental isolates and 67% for clinical isolates (Mala et al., 2014). Meanwhile, in Japan, the ViuB ratio was found to be higher in clinical isolates than in environmental isolates (Yokochi et al., 2013).
Notably, virulence-correlated gene (vcg), 16S rDNA, and CPS operon sequence variations have previously been used to differentiate between the subtypes of V. vulnificus (Nilsson et al., 2003; Rosche et al., 2005; Chatzidaki-Livanis et al., 2006). Wanida Mala et al. (2014) found a combination of both vcg types C and E present in the clinical and environmental isolates, although type E alone was not detected in the environmental isolates. Our results are inconsistent with another study of Baker-Austin and Oliver (2018), who found that 90% of clinical isolates and 93% of environmental isolates exhibited vcg types C and E, respectively. This indicates that V. vulnificus has the ability to transfer genes between its clinical and environmental strains.
DNA sequencing
Direct DNA sequencing of the V. vulnificus and V. parahaemolyticus genomic DNA extracts was performed using different F primers, such as 16S rRNA (A and B), VvhA, ViuB, CPS allele1, vcgC, vcgE, and toxR. The forward sequencing primer exhibited hypervariable regions unique to the V. vulnificus and V. parahaemolyticus strains. Furthermore, upon submitting 100 bp sequences to GenBank for BLAST alignment analysis, sequences corresponding to regions of V. vulnificus and V. parahaemolyticus were identified. The GenBank accession numbers for every gene are noted in Table 5.
DNA Sequencing and Identification according to GenBank for BLAST Alignment Analysis

SDS-PAGE
We also distinguished between the environmental and clinical strains of V. vulnificus by observing the migration total number of proteins for each strain on SDS-PAGE. As shown in Figure 6, at 21.5 and 30 kD, some differences in band arrangement between types B (clinical strain: lanes 3 and 4) and A (environmental strain: lanes 1, 2, 5–8) are evident. These differences were difficult to detect by conducting PCR using 16S rRNA types A and B genes, since the amplicons for both strains appeared to be similar in band and size at 839 bp, as shown in Figure 5.
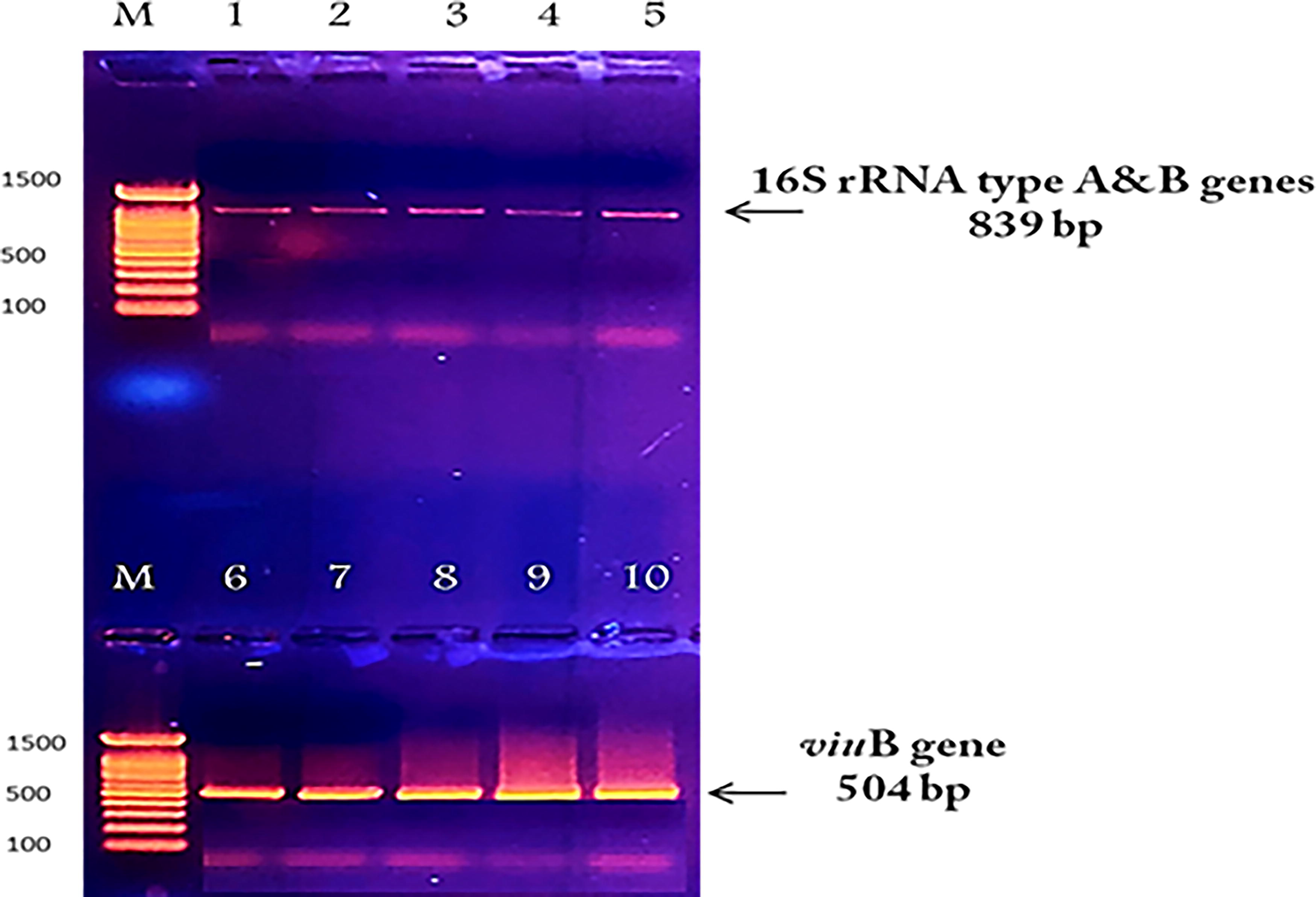
16S RNA type A was detected in environmental isolates (1–3), and type B was found in two isolates (4 and 5). viuB-encoded vulnibactin siderophore was also found in both environmental and clinical isolates (6–10).
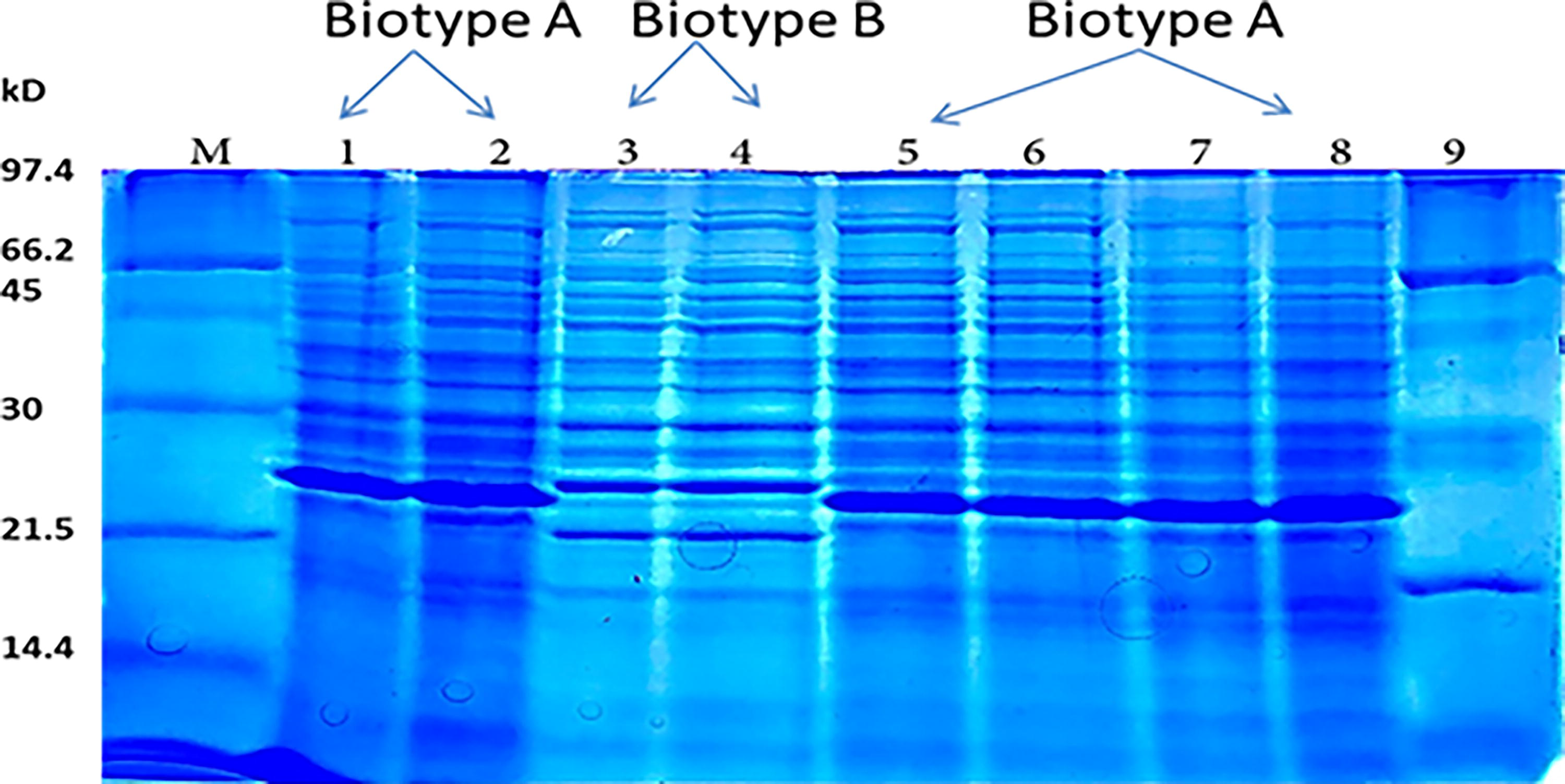
Lanes 1–2, 5–8: Total protein profile of V. vulnificus type A (environmental strain), Lanes 3–4: V. vulnificus type B (clinical strain), Lanes M and 9: Unstained protein ladder standards, low range: 14–97 kD, BIO RAD #1610303.
Antibiotic sensitivity test
Out of the 17 antibiotics investigated in this study, the clinical strain of V. vulnificus was found to be resistant to eight belonging to different classes—beta-lactam, cephalosporin, lincosamide, penicillin, glycopeptide, and tetracycline—achieving a resistance rate of 47.05%. Comparatively, its environmental strain was resistant to only six antibiotics, attaining a ratio of 35.29%, belonging to the beta-lactam, cephalosporin, lincosamide, penicillin, and glycopeptide classes (Table 6).
Antibiotic Sensitivity Test

Along these lines, Baker-Austin et al. (2009) studied the antibiotic resistance of V. vulnificus in South Carolina and Georgia (United States) and found that its clinical isolates were resistant to three or more classes of antibiotics at a ratio of 45%, while its environmental isolates were resistant to 8 or more antibiotic agents, attaining a ratio of 17.3%. The groups of drugs involved in this study include doxycycline, tetracycline, aminoglycosides, and cephalosporin, which are usually prescribed to counter V. vulnificus infection (Baker-Austin et al., 2009).
Since V. vulnificus strains have been exposed to several veterinary and clinical antibiotics (Oliver, 2015; Baker-Austin and Oliver, 2018), their increased resistance to antibiotics has been identified in different environmental niches, leading to the categorization of the antibiotics into three levels: high resistance to tobramycin and cefazolin; intermediate resistance to aztreonam, streptomycin, erythromycin, vancomycin, clindamycin, ampicillin, penicillin, and gentamycin; and low resistance to cefepime, kanamycin, oxytetracycline, tetracycline, nalidixic acid, oxalinic acid, aztreonam, streptomycin, erythromycin, vancomycin, clindamycin, ampicillin, penicillin, and gentamycin (Kurdi Al-Dulaimi and Ariffin, 2019; Pan et al., 2013; Roig et al., 2009). The increasing rate of antibiotic resistance of V. vulnificus in various environments is a result of the extensive use of multiple classes of antibiotics for clinical purposes, aquaculture, and in agricultural fields. Wastewater transports these antibiotics to water bodies, such as rivers and lakes, thereby posing a potential risk to public health (Heng et al., 2017; Hor et al., 1995; Hsueh et al., 2004). Therefore, water bodies not only contribute to increased resistance to antibiotics but also maintain reserves of V. vulnificus, as a result of which these life-threatening pathogens are transmitted into the food chain and human body, causing V. vulnificus infection (Dafale et al., 2020). Regulation of using antibiotics in aquaculture is considered to be an essential approach to prevent the development of antimicrobial-resistant strains of V. vulnificus (Tanveer et al., 2024).
Doubling time
The doubling time of the environmental and clinical types of V. vulnificus was evaluated using rich broth media of tryptic soy at 37°C every 60 min in the logarithmic phase. The results indicated that the doubling time achieved by the clinical isolates was about 14.45 min, while that attained by the environmental strain was 12.04 min (Fig. 7). Along these lines, when Dryselius et al. (2008) determined the doubling times of three Vibrio species incubated in rich media at 37°C, they observed doubling times of 12–14 min for V. parahaemolyticus, 16–20 min for V. cholerae, and 18–22 min for V. vulnificus. The difference between the doubling times estimated by Dryselius et al. (2008) and the current study may be attributed to the use of tryptic soy broth in the latter to suppement the 3% NaCl used in the former.
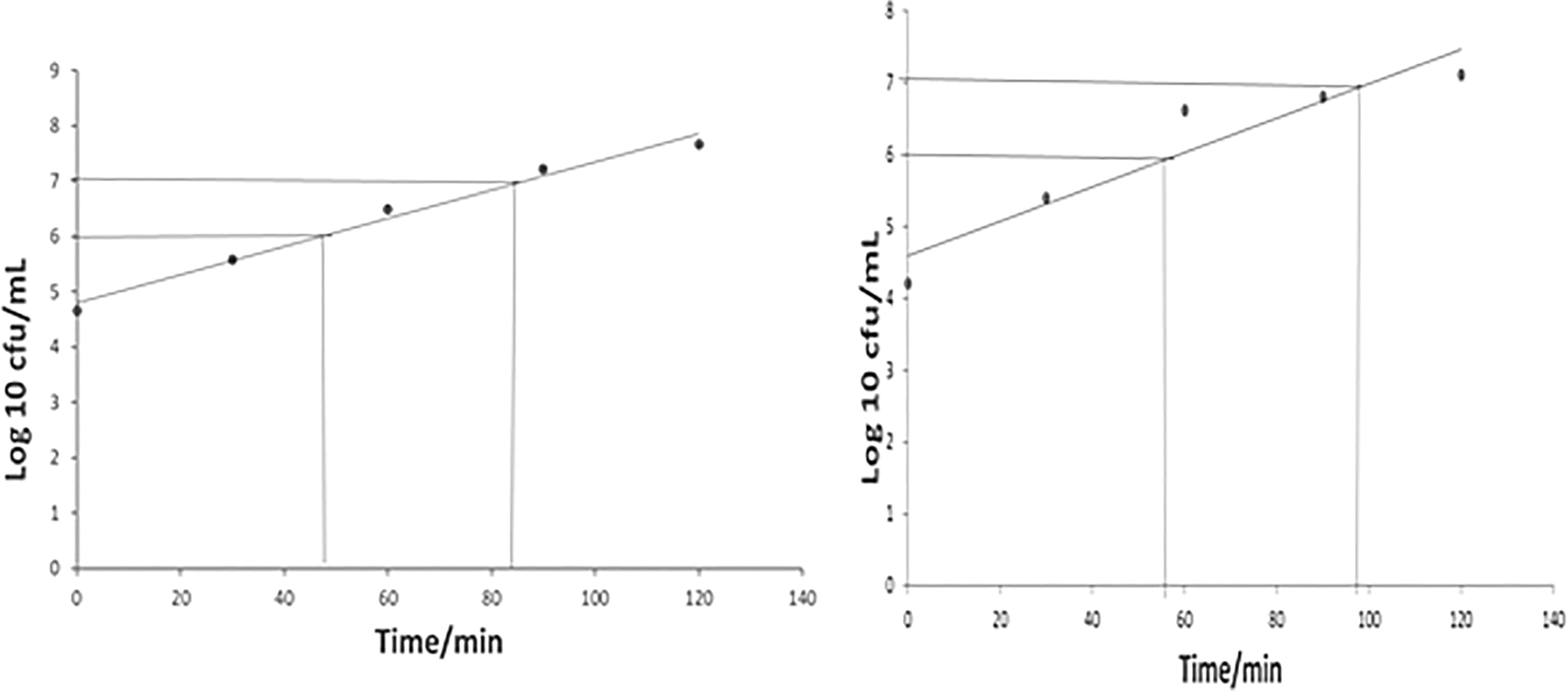
Doubling time of Vibrio vulnificus.
Conclusion
This study highlights the potential risk of contracting a variety of foodborne illnesses when consuming raw or undercooked tilapia fish. Notably, when fish are contaminated with Vibrio sp., no change in taste or smell can be identified. Moreover, various treatments, or even the addition of organic acid salts, sauces, or alcohol, do not kill Vibrio. Therefore, the consumption of undercooked or raw fish increases the risk of exposure to V. vulnificus. Fishe and shellfish are gaining popularity in Saudi Arabia due to increasing variety and availability as a result of economic growth. It is important to be conscious of the fact that cooking fish well and heating them adequately are the primary methods for controlling V. vulnificus infection when consuming these foods. The Food and Drug Administration has also confirmed that eating raw or undercooked seafood can put consumers at an unnecessary risk of infection by foodborne pathogens.
Footnotes
Acknowledgment
This study was supported by Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2025R23), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Funding Information
Princess Nourah bint Abdulrahman University Researchers Supporting Project number (PNURSP2025R23), Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia.
Data Availability
All supporting data are included within the main article and in these sites: https://www.ncbi.nlm.nih.gov/nuccore/PP832854.1?report=fastahttps://www.ncbi.nlm.nih.gov/nuccore/PP425857.1?report=fastabhttps://www.ncbi.nlm.nih.gov/nuccore/MF963048.1?report=fastahttps://www.ncbi.nlm.nih.gov/nuccore/U32676.1?report=fastahttps://www.ncbi.nlm.nih.gov/nuccore/DQ360502.1?report=fastahttps://www.ncbi.nlm.nih.gov/nuccore/OQ718810.1?report=fastahttps://www.ncbi.nlm.nih.gov/nuccore/AY626584.1?report=fasta![]()
Authors’ Contribution
H.M.Y.: Writing—original draft, visualization, validation, software, project administration, methodology, investigation, formal analysis, data curation, conceptualization. N.H.M.: Writing—review and editing, validation, S.M.K. and A.S.A.: Resources, writing—review and editing. M.F.E.: Funding acquisition. A.A. and M.M.A.-D.: Writing—review and editing.
Disclosure Statement
The authors declare that there are no competing interests associated with the article.