
Abstract
Denaturing high-performance liquid chromatography (DHPLC) has been employed as a prescreening tool to reduce the amount of DNA sequencing. It could be a simple and cost-effective screening method for mutations and polymorphisms in exons 4, 5, and 6 of the CD36 gene, which encode the protein region responsible for the removal of oxidized low-density lipoprotein. Genomic DNA was isolated from 306 Caucasian infants of Polish origin. Six single-nucleotide substitutions were detected by DHPLC and confirmed by direct sequencing. The A591T, G550A, and C572T alterations have not been described so far. Each of two nonsynonymous substitutions (Asp184Asn, Pro191Leu) was found in one subject (0.2% minor allele frequency). The results suggest that nonsynonymous alterations in the analyzed CD36 region are rare in Caucasians. DHPLC is a specific and cost-effective technique that may prove to be particularly useful for the identification of polymorphisms and mutations in the CD36 gene.
Introduction
C
More than 20 mutations in the coding sequence of the CD36 gene have been described (Hanawa et al., 2002; Imai et al., 2002; Rać et al., 2007). CD36 deficiency is divided into two phenotypic subgroups. In type I deficiency, neither platelets nor monocytes express CD36, whereas in type II deficiency (very rare in Caucasians, 0.3% of population; more frequent in Asians and Afro-Americans, 3-4% of population), CD36 is expressed in monocytes but not in platelets (Yamamoto et al., 1994). CD36 may play an important role in the pathogenesis of certain metabolic diseases. There have been reports on increased total cholesterol and LDL concentrations in blood serum in association with mutations of the CD36 gene (Yanai et al., 2000). CD36 has been reported to play an important role in atherogenicity (Huszar et al., 2000). Some authors suggest that CD36 gene polymorphisms modulate lipid metabolism and cardiovascular risk in Caucasians (Nozaki et al., 1999; Kintaka et al., 2002; Ma et al., 2004).
Direct sequencing is considered to be the most sensitive mutation detection technique. However, it is technically demanding, time consuming, and expensive. Thus, denaturing high-performance liquid chromatography (DHPLC) has been employed as a prescreening tool to reduce the amount of DNA sequencing. DHPLC is highly sensitive, rapid, and reproducible (Cotton and Bray, 2001; Holinski-Feder et al., 2001; Fasano et al., 2005; Zeng et al., 2007). This technique is based on the detection of heteroduplexes in short DNA segments by ion-pair reversed-phase HPLC. Partial heat denaturation within an acetonitrile gradient results in altered mobility of the heteroduplex relative to homoduplex DNA. DHPLC elution profiles are very characteristic for particular sequence changes and this method allows identifying frequent and previously described alterations with a very high accuracy without sequencing (Kurzawski et al., 2002).
Objectives
The objectives of the study were to develop a screening method for detection of sequence changes in the CD36 gene region encoding fatty acid and oxidized LDL-binding domain and to assess the frequency of such alterations in the Caucasian population.
Materials and Methods
Three hundred six Caucasian infants of Polish origin born in the past 10 years in hospitals of the West Pomerania province were genotyped. Genomic DNA isolated as described (Lahiri and Schnabel, 1993) from umbilical cord blood was taken from the DNA Bank of the Department of Genetics of the Pomeranian Medical University. To define the sensitivity and specificity of detection of changes previously described in a different (Japanese) population, DNA samples with known sequence changes (provided by Prof. Hirokazu Kashiwagi) were used (Kashiwagi et al., 1995, 2001).
CD36 fragments were amplified with primers flanking exons 4, 5, and 6 (Table 1). The reaction mixture of a volume of 25 μL contained 40 ng of DNA, 7.5 pmol of each starter (high purity salt free [HPSF]; MWG-Biotech AG), the polymerase chain reaction (PCR) buffer (10 mM of Tris-HCl at pH 8.9, 50 mM KCl), 1.0-2.0 mM MgSO4, 5 nmol of each of four dNTPs, and 0.3 U polymerase Optimase (Transgenomic). Amplification was performed as follows: an initial 5 min denaturation at 94°C was followed by 35 cycles of PCR with denaturation at 94°C for 30 s, annealing (at 55°C for the first 5 cycles and 52°C for the next 30 cycles) for 30 s, extension at 72°C for 45 s, with final extension at 72°C for 5 min. PCR products were checked for specificity by agarose gel electrophoresis before sequencing or DHPLC analysis. DHPLC analysis was performed using a Hewlett-Packard 1050 chromatographic system consisting of a quaternary gradient pump, an HP G13222A Vacuum Degasser, a Rheodyne 7125 manual injection valve with a 20 μL PEEK loop, an HP G1316A thermostatted column compartment, and an ultraviolet variable wavelength detector with a standard flow cell set at 260 nm. The mobile phase was preheated between the injector and the column. Separation was effected by using an HPLC column Helix DNA 50 × 3.0 mm (CP28353; Varian). Chromatographic data were processed by the HP Chemstation software. The PCR products were denatured at 95°C for 5 min and then reannealed by decreasing the temperature from 95°C to 40°C over a period of 55 min. About 6-8 μL was loaded on the column and separated using a linear acetonitrile gradient. The mobile phase consisted of a mixture of 0.1 M triethylammonium acetate (pH 7.0) with 0.1 mM ethylene diaminetetraacetic acid with (buffer B) or without (buffer A) 25% (v/v) acetonitrile. The flow rate was 0.5 mL/min. After equilibration with 5% below the start value, the linear gradient of buffer B content (9% increase over 3 min from injection time) was adjusted to set the elution time of the homoduplex peak to between 3.7 and 4.2 min. The column was flushed (95% buffer B) between 3.01 and 3.50 min. Equilibration with buffer B at 5% below the start value took 2 min, and thus the total time between injections was 5.5 min. The temperature for heteroduplex analysis was originally established using the DNA Melt software, as described by Jones et al. (1999) (available at http://insertion.stanford.edu/melt.html). The final temperature for optimal resolution of homoduplexes and heteroduplexes for each fragment was determined experimentally. The run temperatures were higher by 1.5°C for exons 4 and 5 and by 2.5°C for exon 6 than those calculated by the Melt software. Finally, heterozygous control probes, from DNA fragments harboring sequence variants (when available), were used to confirm the efficiency of detecting genetic alterations. The experimental conditions for DHPLC analyses are summarized in Table 1. The PCR products with different alterations detected by DHPLC were bidirectionally sequenced using the Applied Biosystems Dye-terminator Cycle Sequencing Ready Reaction kit according to the manufacturer's protocol. Semiautomated sequence analysis was performed using a 373A DNA fragment analyzer (Applied Biosystems).
The samples of detected heterozygotes with identical profiles were mixed by two so as to obtain their equal content of the PCR product assessed on the basis of the peak area in DHPLC. The obtained mixture was then reannealed in the above-described conditions and was loaded on the column. If the mixture profile corresponded to the profiles of single samples, it was considered the confirmation that both these samples contain the same sequence alteration. Samples of homozygotes whose retention time slightly differed from that of the others were reannealed with wild-type (WT) homozygotes, as described earlier, to detect potential variant homozygotes.
Results
The conditions described allowed the analysis of CD36 gene in three amplicons (exons 4-6). Changes detected by DHPLC included six single-nucleotide substitutions. Genotype distributions were consistent with the Hardy-Weinberg equilibrium for all sequence changes [p = 1, exact test according to Guo and Thompson (1992)]. All alterations (known or novel) listed in Table 2 were detected initially by DHPLC and then confirmed by DNA sequencing. For all of them, we obtained very good resolution of the homoduplex and heteroduplex peaks (Fig. 1-3). In all amplicons, homozygotes were characterized by the presence of one narrow homoduplex peak. Only one homozygous sample of all 918 samples was incorrectly classified as a heterozygote different from other variants detected in exon 5. The value of half-height peak width appeared particularly useful in distinguishing between homo- and heterozygotes. It was measured as an attempt to eliminate the possibility of overlooking sequence alterations with worse separation of homo- and heteroduplexes. Further, the potential differences in retention times of homoduplexes were noted. In this manner, the IVS3-6 CC variant homozygotes were found; it differed from WT IVS3-6 TT only by 0.02 s longer retention time.
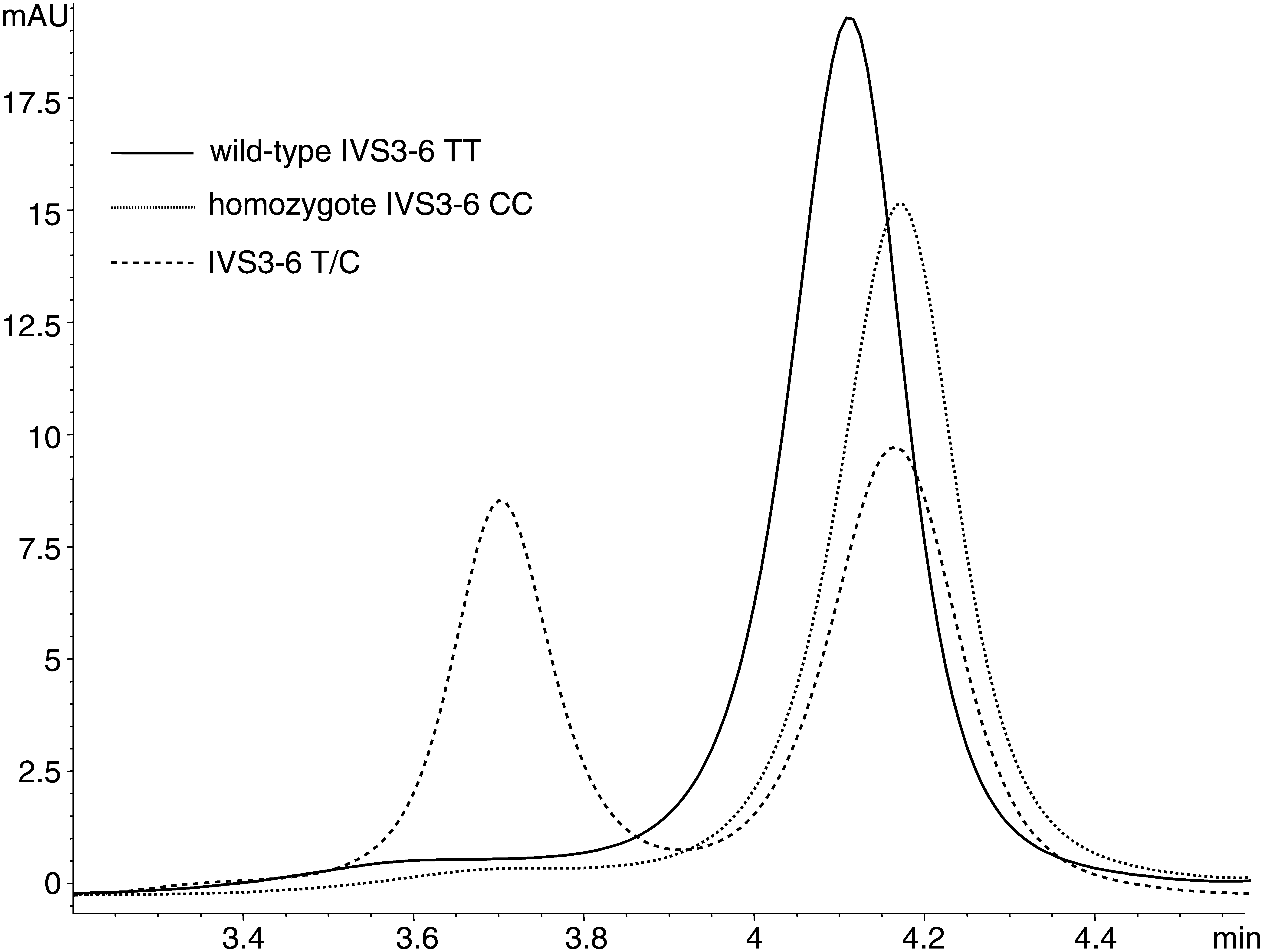
Chromatogram of sequence alterations in intron 3 detected by DHPLC in CD36 gene. DHPLC, denaturing high-performance liquid chromatography.

Chromatogram of sequence alteration in intron 4 detected by DHPLC in CD36 gene.

Chromatogram of sequence alterations in exon 6 detected by DHPLC in CD36 gene.
Chromatograms of samples containing sequence alterations found earlier in the Japanese population (Kashiwagi et al., 1995) (C478T: CT heterozygote and TT homozygote) are presented in Figure 4. The 478TT profile differed slightly from the WT homozygote profile, and after we reannealed 478TT and 478CC homozygotes, the mixture profile corresponded to that of a heterozygote (Fig. 4), which we did not find in the Polish population. However, the profile of the 478CT heterozygote sample was different, because—as shown by sequencing—this sample contained additionally IVS3-6 CT polymorphism in the analyzed amplicon.
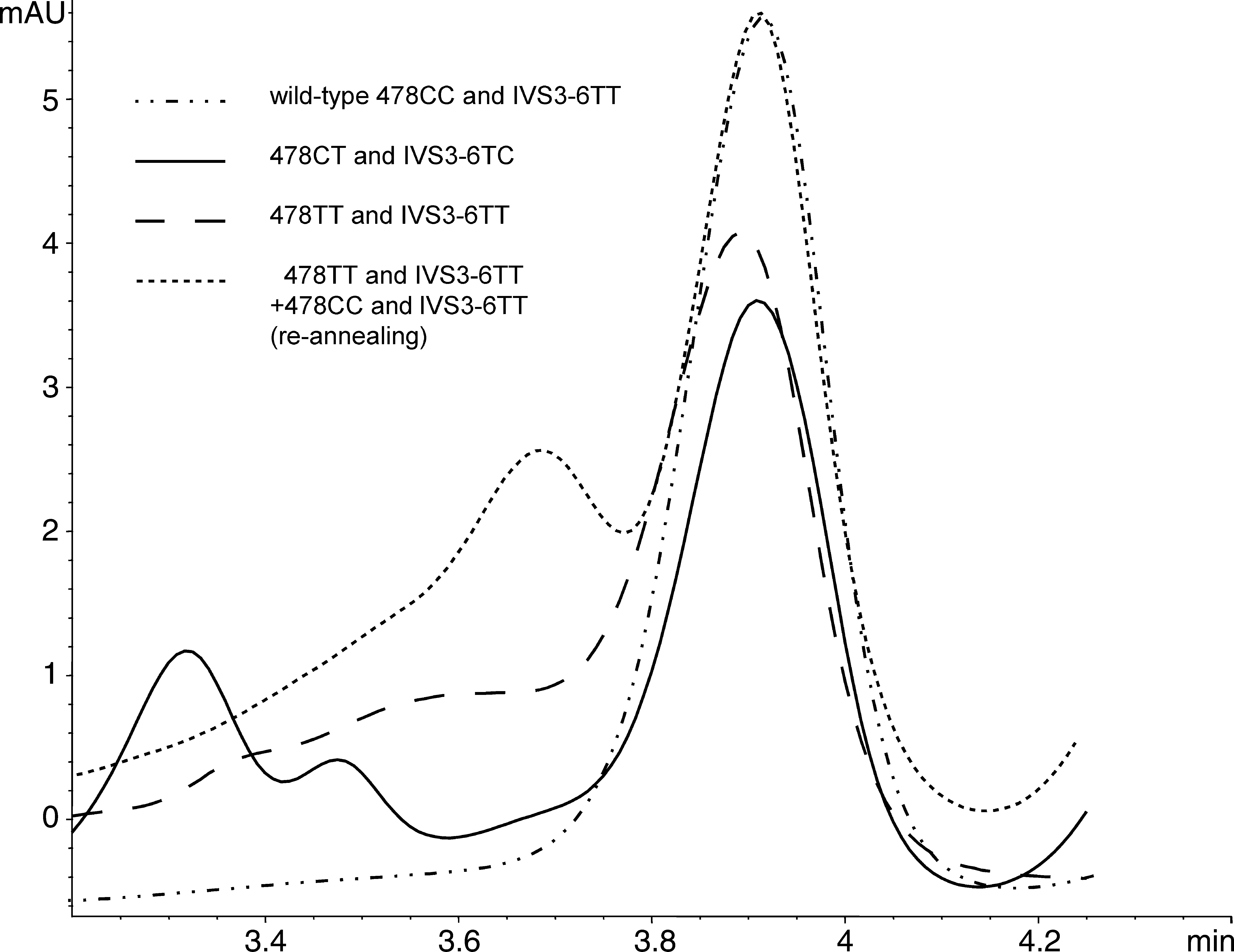
Chromatogram of sequence alterations in exon 4 and intron 3 detected by DHPLC in CD36 gene.
Discussion
Although DHPLC is an inexpensive, highly sensitive, rapid, and reproducible technique for the detection of mutations, the methodology of DHPLC analysis for the CD36 gene has not been described so far. Leprêtre et al. (2004) used DHPLC to analyze the CD36 gene but did not specify the details of the method.
Owing to the use of a high-fidelity Optimase polymerase (Transgenomic), replication error artifacts were minimized and narrow homoduplex peaks were obtained.
Sequencing of all heterozygotes detected by DHPLC would considerably increase the analysis costs. To avoid it, we performed reannealing of heterozygote samples with similar DHPLC profiles, which confirmed identity of their sequence alterations. In DHPLC analysis, variant homozygotes differ usually from WT homozygotes only by the retention time. Their differentiation may sometimes be difficult because the retention time difference could also result from technical factors (e.g., external temperature fluctuations). To explain the changed retention time, such samples were reannealed with WT homozygotes. In two cases, when we observed a longer retention time in exon 4 samples, reannealing demonstrated a heterozygous profile, and sequencing confirmed the presence of homozygous polymorphisms IVS3-6 CC (Fig. 1).
In the present study, no false-negative DHPLC results were observed, similarly to our previous study (Kurzawski et al., 2002) where DHPLC was compared with direct sequencing of MLH1 and MSH2 genes. Only one misinterpretation of two heterozygotes with closely situated changes C2040T and G2041A in the MLH1 gene was described because of similar chromatographic profiles. To prevent such error in the present study, we performed reannealing of heterozygote samples with similar DHPLC profiles. Only one of 818 homozygous samples was incorrectly classified as heterozygote because of poor quality of DNA (0.1% false positives per amplicon). Therefore, high-fidelity polymerase and good quality of genomic DNA ensure low rate of false-positive DHPLC results.
The most common sequence alteration in the exons tested was synonymous transition G573A (Pro191Pro, rs5956). The 573A allele frequency in the population tested was 2.6%, whereas the dbSNP base quotes 4.2%-4.5% for the Caucasian populations. This allele has not been found in the Asian and African populations. Another synonymous alteration A591T (Thr197Thr) was found in three neonates. The only nonsynonymous alterations were G550A (Asp184Asn) and C572T (Pro191Leu), each of them detected in only one subject. The A591T, G550A, and C572T alterations have not been described so far. The Asp184Asn and Pro191Leu alterations may potentially change protein properties, but further studies are necessary to verify this hypothesis.
In intron fragments adjacent to the tested exons, we found the presence of IVS3-6 T/C (rs3173798) and IVS4-10 G/A (rs3211892) polymorphisms. The IVS3-6C (9.6%) allele frequency was similar to that described earlier in the Caucasian populations (6.2%-11.2%); it is much more frequent in the Asian and African populations (21%-41%). The IVS4-10A allele frequency (3.9%) was slightly higher than that described in Caucasians (1.6%-2.6%); this allele is much more common in Africans (21%-36%) and has not been found in Asians. The IVS3-6 T/C polymorphism is located in a conserved splice site (Fry et al., 2009). Some authors report that the minor allele C is associated with protection against neovascular age-related macular degeneration (Kondo et al., 2009). Other authors report that CD36 intron polymorphisms may contribute to individual and population variability in blood lipids (Love-Gregory et al., 2008).
DHPLC has been used for over 10 years as an attractive alternative to direct DNA sequencing. Recently, another method based on differences in melting characteristics of homo- and heteroduplex has become popular: high-resolution melt (HRM) analysis (Vossen et al., 2009). Both DHPLC and HRM are efficient techniques for mutation scanning, with comparable sensitivity (close to 100%) and specificity, but they require high-quality DNA (Millat et al., 2009). Their performance is optimal for the detection of heterozygotes, but homozygous variants can also be identified with high sensitivity when samples are mixed with WT homozygote to generate heteroduplexes (Vossen et al., 2009). Scanning of GC-rich regions has proven problematic for both methods (Tindall et al., 2009). The limitation of HRM analysis is loss of sensitivity with amplicons containing more than two melt domains (Vossen et al., 2009), but it is faster and more cost-effective than DHPLC (Millat et al., 2009). It seems that DHPLC and HRM will co-exist for many years as standard mutation scanning techniques. No HRM method for the analysis of CD36 gene has been described so far.
In summary, DHPLC is a highly sensitive, specific, and cost-effective technique, useful not only for identification of known changes but also to detect alterations that have not been described in the CD36 gene so far. An important advantage of this method is the possibility to detect all sequence alterations within one amplicon in a single analysis. DHPLC may also be efficient in the detection of some homozygous mutations, if samples with a homozygous profile and changed retention time are subjected to reannealing with WT homozygote. Reannealing of heterozygous samples can confirm the presence of a known sequence alteration without the need for direct sequencing. The presented method, together with other methods applied so far, may prove useful in the analysis of the associations between the CD36 genotype and a number of diseases that are related to CD36 receptor function.
Footnotes
Acknowledgments
The authors thank Prof. Kashiwagi of the Department of Hematology and Oncology of the Osaka University Graduate School of Medicine for providing DNA samples. This study was supported by the Ministry of Science and Higher Education (Grant No. 2 P05D 002 30).
Disclosure Statement
No competing financial interests exist.