
Abstract
Aim: The present study was undertaken to investigate a length polymorphism of (GT)n repeats of the heme oxygenase-1 (HMOX-1) gene and its association with serum bilirubin levels in apparently healthy adults. Methods: A total of 211 individuals (normal hematology and liver function test) with bilirubin levels of 1.7 to 22.2 μM were studied. The (GT)n repeats were analyzed by PCR and subsequent sizing by capillary electrophoresis on the ABI Prism 310 Genetic Analyzer. Results: Polymorphisms of the (GT)n repeats were grouped into three classes: short (S) alleles (<20 repeats), intermediate (M) alleles (20-28 repeats), and long (L) alleles (≥29 repeats). The frequencies of the S, M, and L allele groups were 0.10, 0.49, and 0.41, respectively. Carriers of short alleles had significantly higher mean bilirubin levels (13.8 ± 5.10 μM) compared with others (9.18 ± 3.73 μM, p < 0.001). Conclusion: Short (GT)n alleles of the HMOX-1 gene promoter could be a genetic risk factor for hyperbilirubinemia.
Introduction
HMOX is a rate-limiting enzyme in heme degradation that leads to the generation of free iron, biliverdin, and carbon monoxide. Biliverdin is subsequently converted to bilirubin via the action of biliverdin reductase. HMOX-1 and HMOX-2 are the two isoforms that exist in humans. Of these, HMOX-1 is an inducible form of HMOX, and it provides cellular protection against heme and nonheme mediated oxidant injury (Maines, 1997). The human HMOX-1 gene has been mapped to chromosome 22q12, and a (GT)n repeat in the 5′-flanking region of the human HMOX-1 gene is indeed highly polymorphic (Kimpara et al., 1997) and may modulate gene transcription under thermal stress (Okinaga et al., 1996). Therefore, the expression of the HMOX-1 gene could be altered according to the number of (GT)n repeats. Indeed, Yamada et al. (2000) observed that a frequent (GT) dinucleotide repeat polymorphism in the promoter region of the HMOX-1 gene modulates the HMOX-1 response to exogenous stimuli.
Earlier studies have shown that hyperbilirubinemia is associated with (TA)n repeats and the 211 G→A polymorphism in the UGT1A1 gene promoter and coding regions, respectively (Lin et al., 2009). However, it is not very clear what role these polymorphisms play in influencing total bilirubin levels and whether polymorphisms in other genes involved in the bilirubin metabolism pathway also influence total bilirubin levels. Although short HMOX-1 alleles (<25 repeats) were found to have beneficial effects on the serum bilirubin levels and the lipid profile in patients with coronary artery disease (CAD) (Endler et al., 2004), hardly any studies have examined the effect of these repeats within the normal bilirubin range or in healthy people. Hence, the present study was undertaken to assess the association of the length of (GT)n repeats in the HMOX-1 gene promoter with serum bilirubin levels in healthy adults who have bilirubin levels within the normal/mild-increased range.
Materials and Methods
Subjects
A total of 211 unrelated, apparently healthy adults in the age group of 20 to 55 years were randomly selected. All these individuals had normal hematology (hemoglobin, RBCs, WBCs, and platelets) with normal G6PD activity and normal liver function test (albumin, alanine aminotransferase level, aspartate aminotransferase level, alkaline phosphatase, and total protein except for a slight elevation in unconjugated serum bilirubin) and were also negative for hepatitis B surface antigen and anti-hepatitis C antibody.
Laboratory analysis
Blood samples were collected after obtaining informed consent. RBC indices were determined using an automated blood cell counter (Sysmex K-1000; Kobe). Liver function tests were done on an automated biochemistry analyzer (Cobas 111; Roche). An ELISA using commercially available kits (Hepanostika HBsAg Ultra, Biomerieux bv; Hepanostika HCV Ultra, Beijing United Biomedical Co. Ltd) was used for screening for hepatitis B surface antigen and anti-hepatitis C antibody. Screening for G6PD deficiency was done by the dichlorophenol-indophenol (DPIP) dye decolorization method (Bernstein, 1962).
DNA analysis
High-molecular-weight genomic DNA was extracted from the peripheral blood leukocytes using a commercially available DNA extraction kit (Qiagen GmbH). The 5′-flanking region containing (GT)n repeats of the HMOX-1 gene was amplified by PCR with a FAM-labeled sense primer and an unlabeled antisense primer as previously described (Yamada et al., 2000). The sizes of the PCR products along with a 500 LIZ size standard were analyzed using a laser-based Gene scan application on an ABI PRISM 310 Genetic Analyzer (Applied Biosystems). The resultant data were analyzed using the Genotyper software. Normally, an individual has two different sizes of (GT)n repeats from different alleles. Each repeat number was calculated from the observed PCR product size according to the 5′ flanking region sequence of the HMOX-1 gene.
Statistical analysis
The Student's t-test was used to test for statistical significance between the groups.
Results
The number of (GT)n repeats in the HMOX-1 gene promoter varied between 11 and 39 in all the individuals studied. The distribution of the number of (GT)n repeats was found to be trimodal, with one peak located at 14 GT repeats and two other peaks located at 23 and 30 GT repeats (Fig. 1). We, therefore, grouped the alleles into three subclasses, according to the number of (GT)n repeats as described by Yamada et al. (2000). The lower component, with <20 GT repeats, was designated as Short (S) alleles; the middle component, with 20 to 28 GT repeats, was designated as Medium (M) alleles; and the upper component, with ≥29 GT repeats, was designated as Large (L) alleles. The repeat numbers 23 and 30 were found to be the most common in the studied population. The frequencies of S, M, and L allele groups were 0.10, 0.49, and 0.41, respectively.
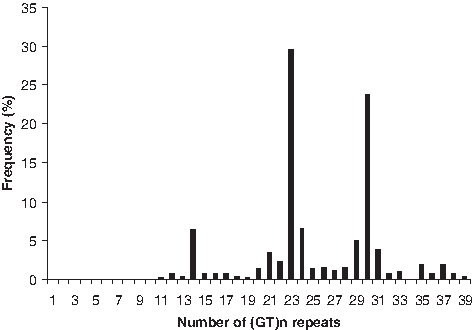
Distribution of (GT)n repeats of heme oxygenase-1 (HMOX-1) gene.
Table 1 shows the frequency distribution of the genotypes of (GT)n repeats along with mean total serum bilirubin levels. Six different genotypes, namely, S/S, S/M, S/L, M/M, M/L, and L/L were observed with a frequency of 4.7%, 12.4%, 0.9%, 25.1%, 35.1%, and 21.8%, respectively. Serum bilirubin levels in the healthy adult individuals varied from 1.7 to 22.2 μM with a mean of 9.9 ± 4.32 μM. Mean serum bilirubin levels were found to be significantly higher in S/S (16.4 ± 4.97 μM) and S/M (12.8 ± 5.01 μM) than in those with M/M (10.3 ± 4.27 μM), M/L (9.1 ± 3.60 μM), and L/L (8.1 ± 2.90 μM) genotypes (p < 0.001). Further, to look for the effect of the S allele on total bilirubin levels, six different genotypes of (GT)n repeats were divided into two groups, according to their subclasses: group I with carrier of the S allele (S/S and S/M) and group II without the S allele (M/M, M/L, and L/L). We excluded S/L alleles from group I mainly because serum bilirubin levels of S alleles and L alleles are expected to be at two extremes (which would neutralize the effect of each other) as well as due to the smaller sample size in the SL genotype. Mean serum bilirubin levels were found to be significantly higher in group I (13.8 ± 5.10 μM) as compared with group II (9.18 ± 3.73 μM, p < 0.001).
Discussion
Severe hyperbilirubinemia is a serious medical problem, especially in newborn infants. If not properly treated, it may cause kernicterus, which is fatal or could lead to neurological disorders. Hyperbilirubinemia in adults is mainly caused by abnormal liver function, hemolysis, and/or a genetic defect (Huang et al., 2005). Liver diseases, such as viral hepatitis and liver cirrhosis along with hepatocellular carcinoma, are very common and a leading cause of death. Therefore, an appropriate differential diagnosis for the cause of hyperbilirubinemia is an important issue.
Bilirubin is a natural product of heme catabolism by HMOX. In the present study, we have demonstrated that there is an association between HMOX-1 promoter polymorphism and serum bilirubin levels. The mean serum bilirubin level was significantly higher in carriers of the S allele than in those with the L/L genotype, indicating that individuals carrying this allele might have higher levels of HMOX-1 and an increased production of the endogenous antioxidant bilirubin. Since there are other factors such as TA repeats and 211 G→A polymorphisms in the UGT1A1 gene that are known to be responsible for higher bilirubin levels, it is difficult to predict the contribution of each polymorphism on the phenotype of raised bilirubin levels. Nevertheless, our unpublished data showed that 9% of individuals who are the carriers of the S allele were normal for both TA repeats and 211 G→A polymorphisms. This suggests that the short (GT)n allele of the HMOX-1 gene may alone be responsible for increased unconjugated serum bilirubin levels. The effect of the (GT)n repeat polymorphism on total bilirubin (TBIL) levels was found to be different in the populations with distinct genetic backgrounds and living conditions (Lin et al., 2009). In previous case control studies, individuals with short (GT)n alleles had significantly higher serum bilirubin concentrations within the normal range and were associated with a significant and marked reduction in CAD risk and incidence of ischemic heart disease (Hopkins et al., 1996; Djousse et al., 2001; Chen et al., 2008). On the other hand, Lüblinghoff et al. (2009) did not find any association between HMOX-1 genotypes and bilirubin concentration among patients with CAD. This probably indicates that other genes involved in bilirubin metabolism superimpose on the effects of different HMOX-1 genotypes. One candidate gene is UGT1A1, and this has been extensively studied. This enzyme is responsible for the glucuronidation of bilirubin and, thus, enhances its elimination. A variation in the promoter and coding regions in the UGT1A1 gene has been shown to be associated with higher serum bilirubin levels (Lin et al., 2009).
The functional relevance of (GT)n repeat promoter variants in the HMOX-1 gene is still under debate. Yamada et al. (2000) described a higher response of reporter gene constructs with short (GT) repeats to H202 exposure in A549 and Hep3B cell lines. Similar results were obtained by analyzing HMOX-1 mRNA and activity in lymphoblastoid cell lines from subjects homozygous for short or long repeats (Hirai et al., 2003) or by analyzing the HMOX-1 expression in mononuclear cells from patients who underwent coronary angiography (Brydun et al., 2007). Considerable evidence supports a crucial role for HMOX-1 in blocking the growth of vascular smooth muscle cells. This antiproliferative effect of HMOX-1 is mediated primarily via the release of CO, which inhibits vascular smooth muscle cell growth via multiple pathways. Pharmacologic or genetic approaches targeting HMOX-1 or CO to the blood vessel wall may represent a promising novel therapeutic approach in treating vascular proliferative disorders (Durante, 2003).
In conclusion, our findings suggest that adults who carry short (GT)n alleles of the HMOX-1 gene promoter are at a high risk of developing unconjugated hyperbilirubinemia. Considering the antioxidant and antiatherogenic properties of bilirubin, the beneficial influence on serum bilirubin levels in carriers of the S allele observed in this study might have been sufficient to exert a protective effect against the development and clinical severity of CAD. Our findings could also be helpful to clinicians when attempting a definitive diagnosis for the cause of unconjugated hyperbilirubinemia.
Disclosure Statement
No competing financial interests exist.