
Abstract
Hereditary inclusion body myopathy (HIBM) is a young-adult onset autosomal recessive disorder caused by a hypomorphic rate limiting enzyme of sialic acid biosynthesis. The enzyme is UDP-N-acetylglucosamine 2-epimerase/N-acetylmannosamine kinase, and is encoded by the GNE gene. HIBM causes slowly progressive muscle weakness and atrophy. Patients are typically diagnosed at 20-30 years of age, and most patients are incapacitated and wheelchair-confined by 30-50 years of age. Some sialic acid containing glycoproteins, including neural cell adhesion molecule (NCAM), are hyposialylated in HIBM muscle biopsy samples. We developed a method to allow detection of serum NCAM sialylation using Western blot, and tested serum samples from several patients and a HIBM mouse model. Preliminary results showed a clear difference in polysialylated and hyposialylated forms of NCAM extracted from serum, and showed NCAM is hyposialylated in HIBM serum samples. This initial finding may prove useful in reducing the need for serial muscle biopsies in HIBM treatment trials. Additional studies are underway to further validate this finding and to evaluate the specificity, reliability, and robustness of this potential serum biomarker for HIBM.
Introduction/Background
H
The major carrier of polysialic acid (PSA) in mammals is the neural cell adhesion molecule (NCAM) which binds homopolymers of alpha-2,8-linked PSA chain to form PSA-NCAM (Cremer et al., 1994). Besides its importance in neuronal development (Brusés and Rutishauser, 2001), PSA-NCAM is also thought to play a significant role in myogenesis and neuromuscular development, and in the structure and function of the neuromuscular junction (Rafuse and Landmesser, 1996; Rafuse et al., 2000). Due to its hyposialylated state in HIBM muscle biopsy samples, PSA-NCAM has been proposed as a potential biomarker for HIBM in muscle biopsy specimens (Ricci et al., 2006; Broccolini et al., 2010). Hyposialylation of HIBM skeletal muscle glycoproteins, including NCAM and Neprilysin, has been previously described (Ricci et al., 2006; Broccolini et al., 2008). Prior attempts to detect sialic alteration of serum glycoproteins in HIBM have been unsuccessful as most general serum proteins are derived from hepatocyte or B cell secreted proteins (Savelkoul et al., 2006; Huizing and Krasnewich, 2009). Here we describe the specific hyposialylation of polysialylated NCAM (PSA-NCAM) in serum of both human patients affected by HIBM and an HIBM mouse model suggesting that targeted analysis of muscle-derived proteins in serum may detect the biochemical alterations of HIBM.
Materials and Methods
The study described was performed by HIBM Research Group, a nonprofit molecular genetics laboratory specializing in performing high complexity clinical testing, regulated under the Clinical Laboratory Improvement Amendments of 1988, California Laboratory Field Services, and accredited by the College of American Pathologists.
All reagents were purchased from commercial sources and each batch validated in compliance with clinical diagnostic quality standards. Reagents and containers were sterile and nuclease free unless otherwise noted. Optical density measurements were performed using the Nanodrop ND-1000 Spectrophotometer® (Nanodrop Technologies).
Specimen
Western blot analysis of PSA-NCAM was performed on human samples of control serum (nonmyopathic), myopathic serum (non-HIBM), and HIBM serum samples. Serum from mice bearing GneM712T mutation was also tested, using surviving homozygous progenies in FVB; B6 background strain.
Protein extraction
Protein extraction was performed using the Mem-PER Eukaryotic Membrane Protein Extraction Reagent Kit (Thermo Scientific). Fifty microliters of the serum sample was mixed with 150 μL of Reagent A (detergent) and vortexed for a homogeneous suspension. Samples were incubated for 10 min at room temperature, and occasionally vortexed (e.g., 5 s every 3-4 min). The lysed cells were placed on ice. Each sample was then mixed with a 450 μL 2:1 dilution of Reagent C and Reagent B respectively (Reagents B and C must be kept at 4°C or on ice at all times). Samples were then incubated on ice for 30 min and vortexed every 5 min. Tubes were centrifuged at 10,000 g for 3 min and the supernatant was collected in new tubes and incubated for 30 min in a 37°C water bath to separate the membrane protein fraction. Samples were centrifuged at room temperature for 2 min at 10,000 g to isolate the hydrophilic fraction from the hydrophobic fraction. The hydrophilic phase (top layer) was carefully removed from the hydrophobic phase (bottom layer) and collected. Phase separations were performed as efficiently and quickly as possible to avoid disappearance of the separate phase interface between layers that occurs at room temperature.
Protein sample purification
SDS-PAGE Sample Prep Kit (Thermo Scientific) was used for protein sample purification.
Resin preparation and sample addition
Twenty-five microliters of PAGE-prep Protein Binding Resin was added to the protein sample using a cut or large-orifice pipette tip. Ten microliters of sample with protein concentration of 30-45 μg/μL was added to resin protein and vortexed. One volume of 100% DMSO equal to the sample volume (10 μL) was added. Tubes were briefly vortexed. Samples were then incubated for 5 min at room temperature and occasionally vortexed to ensure maximum protein adhesion to the resin. Samples were then centrifuged at 2000 g for 2 min. Flow-through was discarded and the collection tube was blotted on paper towel to remove remaining solution.
Wash
Samples were washed using a 300 μL mixture consisting of a 1:1 ratio of DMSO and water. Three hundred microliters of Wash Solution was added to the resin and vortexed until a homogeneous suspension was obtained. Samples were centrifuged at 2000 g for 2 min. Flow-through was discarded and collection tube was blotted on paper towel to remove the remaining solution. Wash was repeated one additional time for a total of two washes.
Elution
Spin cups were transferred to a new collection tube and 50 μL of Elution Buffer was added to the resin. Tubes were capped and briefly vortexed to obtain a homogeneous suspension. Samples were then incubated at 60°C for 5 min. After incubation tubes were capped and vortexed. Samples were centrifuged at 2000 g for 2 min. Eluate in collection tube was retained and stored.
Desialylation using neuroaminidase
Serum samples from healthy nonmyopathic individuals were desialylated by treatment with Vibrio cholerae neuroaminidase (Sigma). Twenty microliters of normal nonmyopathy serum was treated with 0.5 μL of enzyme and incubated 4 h in a 37°C water bath, along with a parallel tube without enzyme.
Western blot analysis of serum NCAM
Western blot analysis was conducted using the Mini Trans-Blot Electrophoretic Transfer Cell (Bio-Rad). Final protein concentration was measured by BradfordUltra (Expedeon), and 390 μg protein (15 μL) loaded per lane for electrophoresis separation. Purified protein sample was mixed with loading dye in a 1:1 ratio. Fifteen microliters of each sample was loaded. Loading dye was prepared as follows: Laemmli Sample Buffer (Bio-Rad) was mixed with 2-Mercaptoethanol (Bio-Rad) in a 20:1 ratio. Samples were then denatured at 95°C for 3 min using the ThermoHybaid PCR Express, model number HBPX110 (Hybaid Corporation), or the Techne Model TC-412 (Barloworld Scientific). Denatured samples were loaded on a Mini-Protean® Precast Gel (Bio-Rad) and run at 100 V for 1 h and 20 min in running buffer (1:10 diluted solution of 10× Tris/Glycine/SDS buffer [ISC BioExpress]). After the run, the gel was equilibrated for 15 min in transfer buffer (1:10 diluted solution of 10× Tris/Glycine Buffer (ISC BioExpress) with one added milliliter of 10% SDS solution (Bio-Rad)). Samples were then electrophoretically transferred using fiber pads, filter paper, and Immun-Blot™PVDF Membrane (Bio-Rad), all of which had been previously soaked in transfer buffer for at least 30 min at 4°C. The transfer was completed at 30 V overnight at 4°C. The next morning, the membrane was placed in 1:10 diluted solution of Ponceau S Stain (amresco) to visualize the bands before proceeding. The stain was washed off completely using triple distilled water or TBST (1:20 diluted solution of 20× Tris-Buffered Saline (ISC BioExpress) with one added milliliter of Tween 20 (Bio-Rad)). The membrane was then incubated in blocking buffer (5% Blotting Grade Blocker Non-Fat Dry Milk TBST solution) for 1 h at room temperature, while shaking at 25 rpm. The membrane was next incubated using 200 μg/mL NCAM (RNL-1) mouse monoclonal IgG (Santa Cruz Biotechnology) diluted in blocking buffer at 1/10,000 for 2 h at room temperature, shaking at 25 rpm. The membrane was washed in TBST for 25 min at room temperature at 25 rpm. The TBST was switched out for fresh TBST every five minutes at which point the membrane container was rotated 90°. After being washed, the membrane was incubated using 2 mg/mL goat anti-mouse IgG (abcam) at 1/10,000 and Precision Protein StrepTactin-HRP Conjugate (Bio-Rad) at 1/100,000 in blocking buffer for 1 h at room temperature at 25 rpm, and washed again as described above. The membrane was incubated in 5 mL of SuperSignal® West Dura Extended Duration Substrate (Thermo Scientific) for 5 min and subsequently dried and stored in a cassette to be developed following 30 s film exposure. Band images were measured for density using the image processing program ImageJ (NIH).
Results
Western blot analysis of serum NCAM sialylation was performed on serum samples obtained from several patients, as follows: three healthy volunteers with no myopathy, two patients with HIBM who had received no treatment, one patient with HIBM who had been consuming ManNAc of his own volition (anecdotally reported by the patient), and one patient with quadriceps sparing nonHIBM myopathy without GNE mutation and showing no hyposialylation on muscle biopsy NCAM (Broccolini et al., 2010). In all control serum samples (from unaffected individuals without muscle disease), sialylated NCAM migrated as one or more bands at approximate size of 150 kD. This signal weakens considerably when serum is treated with neuraminidase (sialidase). The density of the 150 kD bands of noncontrol human samples was measured relative to the average density of control human samples. Each human sample was compared to the standard individually. Serum samples from HIBM patients show a distinctly weaker signal at 150 kD similar to neuraminidase treated control serum. Serum samples from treated HIBM patients show a signal at 150 kD that is stronger than that of nontreated HIBM patients, but slightly weaker than control serum samples. In NonHIBM myopathy samples, the density of the 150 kD band is slightly weaker than normal control serum samples but noticeably stronger than HIBM samples (Fig. 1).

To verify that the difference in the Western blot image for HIBM serum samples is due to hyposialylation, control serum samples were desialylated (treated with sialidase). In the desialylated human controls, NCAM showed as a weak signal at 150 kD that resembled the NCAM band at 150 kD in HIBM serum samples. When compared with normal serum samples, the desialylated lanes show an extra clearly visible yet hazy band between 37 and 25 kD that is not present in normal serum samples. This hazy band is absent in HIBM serum samples and may represent a sialic acid rich glycan that has been detached as a result of the desialylation process.
Western blot analysis of serum NCAM sialylation was also performed on several groups of mice: M712T homozygous without treatment (water alone), M712T homozygous with N-acetylneuraminic acid (NeuAc) treatment, M712T homozygous with ManNAc treatment, wild-type control, and desialylated wild-type control. Wild-type serum samples show signals at 300 and 200 kD, and a weaker signal at 150 kD. The average density of the 200 kD bands of noncontrol mouse samples was measured relative to the average density of control mouse samples. The signal strength of wild-type samples is stronger than any other sample. Homozygous HIBM mice serum samples show a very weak but visible signal at 200 kD. Homozygous HIBM mice treated with NeuAc, the most common sialic acid found in mammalian cells, or ManNAc for just 1 month show a stronger signal at 200 kD than untreated HIBM mice. Both treated HIBM mice serum samples however, still show a weaker signal than seen in wild-type serum (Fig. 2). In desialylated wild-type mouse controls, the sample shows very weak signals above 88 kD, closely resembling the NCAM signal in HIBM mouse serum samples.
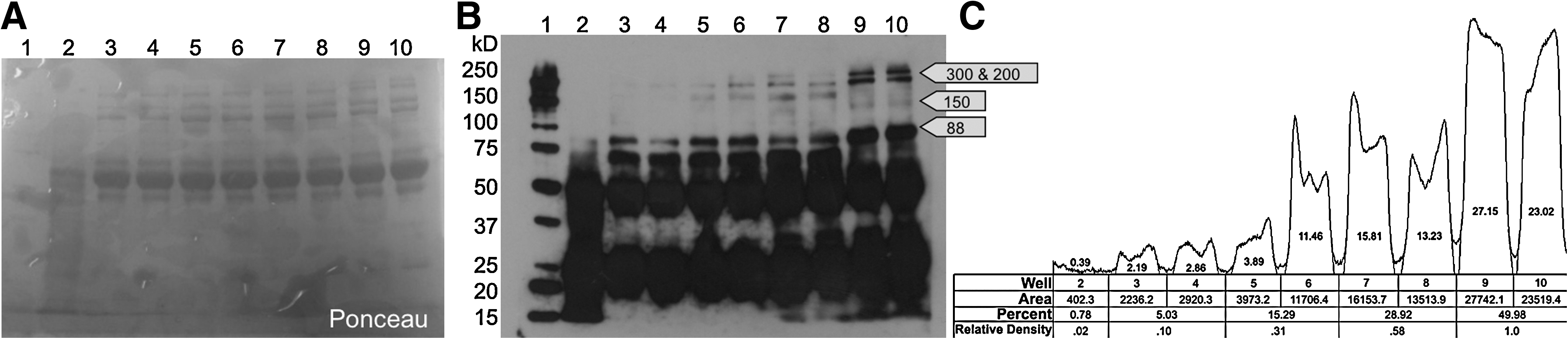
Mouse Serum NCAM Western blot.
To confirm that what looks to be hyposialylation in the desialylated normal serum samples is not a result of protein breakdown due to their incubation at 37°C, normal serum samples without desialylation enzyme were incubated along with the normal serum samples with desialylation enzyme and analyzed by Western blot. The normal serum samples that had been incubated at 37°C showed the same band makeup as normal control serum samples and showed no evidence of protein degradation. Thus, the band makeup of the normal serum samples that were incubated with desialylation enzyme was likely due to desialylation and not incubation induced protein degradation.
Discussion
We developed and tested a method to allow Western blot detection of NCAM sialylation in samples of human and mouse serum. Previous examination of PSA-NCAM in normal muscle samples showed that it migrates as a broad band at 150 kD (Ricci et al., 2006). In this study, Western blot analysis of normal control mouse serum showed PSA-NCAM migrating at 300, 200, and 150 kD, whereas normal control human serum NCAM migrated at 150 kD. In both mouse and human serum NCAM, a clear difference is seen between control bands with strong signals and HIBM serum bands that show much weaker signals and resemble desialylated control serum samples. These initial findings provide early evidence that serum biomarkers targeting PSA may be developed for HIBM. If soluble PSA-NCAM or other sialylated glycoproteins (sialoprotein) found in serum partly originate from muscle, then improved sialylation of a serum sialoprotein marker may reflect the restoration of sialylation in the muscles of an HIBM patient as they are treated; thus, potentially reducing the need for more invasive serial muscle biopsies during interventional clinical trials. ManNAc or NeuAc treatment prevents muscle disease in the HIBM mouse model (Malicdan et al., 2009). Since ManNAc or NeuAc also improve sialylation of NCAM, our findings suggest that biomarkers targeting PSA may ultimately prove useful as a surrogate outcome measure in HIBM clinical trials.
Further studies, perhaps with more specific monoclonal antibodies and an increased number of subjects, are necessary to validate the use of serum NCAM as a reliable and robust biomarker for HIBM. Other antibodies that preferentially bind to the NCAM protein backbone or PSA can be used to develop a sensitive and quantitative ELISA based assay.
Conclusion
Western blot analysis of PSA-NCAM was performed on human control serum (nonmyopathic), human myopathic serum (non-HIBM), and human HIBM serum samples. Serum from mice bearing GNEM712T mutation was also tested. There is a marked difference in the signal strength between the control samples and all other sample groups, with control samples showing the strongest signal. This difference seems to be reduced by ManNAc treatment in both patients and the HIBM mouse model. Future work includes development of such a quantitative assay and determining if improvement of PSA-NCAM sialylation level correlates with functional benefit in human HIBM patients.
Footnotes
Acknowledgment
The authors extend their gratitude to Advancement of Research for Myopathies (ARM) who supported this study.
Disclosure Statement
E.K. is an employee, stockholder and executive of Ultragenyx, a company developing a treatment for HIBM. D.D. and Y.V.-A. are collaborating with Ultragenyx, and may gain rights to exercise Ultragenyx stock options.