
Abstract
Cytokine-induced expression of suppressors of cytokine signalling (SOCS) molecules is important for the negative feedback control of STAT-dependent cytokine signalling. The aim of this study was to investigate possible association between the promoter region polymorphisms of the SOCS3 gene and metastatic colorectal carcinoma in a Turkish population. The DNA samples obtained from 103 patients and 109 healthy individuals were analyzed by polymerase chain reaction/single-strand conformation polymorphism (SSCP), and nucleotide sequence analysis. Five sets of primers designed for the SOCS3 gene were used, and we did not detect significant differences in genotype frequencies for any of these polymorphisms between the study groups. Only the S3P1 region showed polymorphism and displayed three (1,2,4, 2,3,4 and 2,4) genotypes. Interestingly, 2,3,4 genotype was observed in 3 patients, but not in controls. Moreover, the sequence analysis revealed that the nucleotides positioned at −914 and −1031 nt had the polymorphisms. Nucleotide sequence analysis of SSCP band 1 and band 3 revealed C-914A (rs12953258) and T-1031C (rs111033850) polymorphisms, respectively. The T-1031C polymorphism lies in the border of the STAT-binding site. The T-1031C polymorphism (rs111033850) is a newly identified single nucleotide polymorphism with this study, and we submitted this to the NCBI database. However, these results suggested that there is no marked association between SOCS3 gene promoter region polymorphisms and the risk of developing metastatic colorectal cancer.
Introduction
Ligand binding to cytokine receptors causes activation of the Janus kinases (JAKs) and subsequent phosphorylation of signal transducers and activators of transcription (STATs) (Shuai and Liu, 2003; Croker et al., 2008). Phosphorylated STATs dissociate from the receptor and form homo- or heterodimers. STAT dimmers, then translocate to the nucleus, where they bind specific DNA elements and alter the expression of target genes in response to cytokine stimulation (Kile and Alexander, 2001). An appropriate cellular response depends not only on receptor/cytokine interaction and the initiation of intracellular signalling, but also on the timely and effective termination or modulation of signalling. STAT3 typically acts as an oncogene (Bromberg et al., 1999). STAT3 regulates expression of VEGF and is associated with angiogenesis and tumor progression (Niu et al., 2002). The negative regulation of cytokine-induced signalling pathways involves a number of proteins. There are three different classes of negative regulators of cytokine action: the Src homology (SH)2-containing protein tyrosine phosphatase I (SHPI), the protein inhibitors of activated STATs (PIAS), and the suppressors of cytokine signalling (SOCS). The SOCS-protein family consists of 8 members; SOCS1-7 and a cytokine-inducible SH2-containing protein (CIS). Although the basal levels of the SOCS protein family are often low or undetectable in resting cell or tissues, SOCS expression is generally induced by a wide range of cytokines, hormones, and growth factors (Kile and Alexander, 2001; Krebs and Hilton, 2001; He et al., 2004). Expression of SOCS-3 is normally induced by cytokines through JAK/STAT signaling in normal cells, which in turn inhibits JAK activity and subsequent STAT3 activation. The expression of SOCS-3 protein is very short-lived with rapid turnover after stimulation with cytokines (Hilton, 1999; Siewert et al., 1999). Meanwhile, constitutive SOCS-3 expression is also observed in breast carcinoma (Raccurt et al., 2003), in breast cancer cell lines (Waters et al., 2006), cutaneous T-cell lymphoma (Brender et al., 2005), and chronic myeloid leukaemia (Sakai et al., 2002; Roman-Gomez et al., 2004). SOCS proteins could play a role in the initiation or progression of cancer. Therefore, SOCS are important negative feedback regulators of the JAK/STAT signaling pathway, and have been recently investigated for their role in the development of different cancers.
Although development of CRC is a complex process influenced by multiple factors, there is no clear understanding of the genetics of CRC formation. The human SOCS3 gene maps to chromosome 17q25.3 and consists of two exons spanning 2,734 nucleotides (www.ncbi.nlm.nih.gov/nuccore/NG_016851.1?&from=5000&to=8299&report=graph).
SOCS family proteins are characterized by an amino-terminal (N-terminal) region of variable length, a central SH2 domain, and a conserved carboxy-terminal (C-terminal) domain termed the SOCS box (Yoshimura et al., 1995; Hilton et al., 1998; Elliott, 2008). Abnormalities in the JAK/STAT signaling pathway have been reported to be associated with cancer (He et al., 2004). However, mechanisms for involvement of SOCS-3 in CRC have not been well studied. It has been demonstrated that SOCS3-positive cells were significantly increased in colonic epithelium of both inactive and active ulcerative colitis, but the proportion of SOCS3-positive cells was reduced in dysplasia and cancer (Li et al., 2010). Stat6high HT-29 human colon cancer cell line expressed low constitutive SOCS3 mRNA. DNA demethylation using 5-aza-2-deoxycytidine in HT-29 cells up-regulated mRNA expression of the SOCS3 gene (Liu et al., 2009). Until now, no report of promoter polymorphism of the SOCS3 gene in patients with CRC has been published. Therefore, the purpose of this study was to test a possible association between SOCS3 gene promoter polymorphisms and the risk of metastatic CRC development in a Turkish population.
Materials and Methods
Study populations
Our study group consisted of a total of 103 unrelated Turkish patients (age; 29.6±17.4 years) with histopathologically confirmed metastatic CRC, who underwent treatment at Gaziantep University Hospital. The control group consisted of 109 unrelated healthy Turkish subjects (age; 29.0±8.5 years) without a history of carcinoma. All of the patients and controls were recruited from the same racial, ethnic, and the same geographical area (southeastern Turkey) and environment stratification. The study was approved by the local Ethics Committee, and it was conducted in accordance with the guidelines in the Declaration of Helsinki.
Blood samples and DNA isolation
Venous blood (10 mL) was collected by venapuncture into sterile siliconized Vacutainer tubes with 2 mg/mL disodium ethylenediaminetetraacetic acid. Immediately after collection, whole blood was stored at −20°C until use. Genomic DNA was extracted from whole blood using a salting-out method and stored at −20°C (Miller et al., 1988).
Molecular genetic methods
Promoter polymorphisms of the SOCS3 gene was analyzed according to standard polymerase chain reaction (PCR) protocols. All primers were designed to amplify the specified regions using the web-based primer design program (http://workbench.sdsc.edu). Nucleotide sequences of the PCR primer sets, the sizes of the amplicons, and the annealing temperature are listed in Table 1. PCR reaction was performed in a total volume of 25 μL containing 0.1 μL genomic DNA, 2.5 μL MgCl2, 2 μL dNTPs, 0.4 μL forward primer, 0.4 μL reverse primer, 0.1 μL of Taq polymerase, 2.5 μL 10xPCR buffer, and 17 μL ddH2O. PCR conditions were as follows: after denaturation at 95°C for 5 min, 35 cycles were performed (95°C for 1 min, annealing temperatures (see Table 1) for 40 s, 72°C for 30 s), followed by extension at 72°C for 5 min to amplify the target DNA. Amplification was achieved with in-house MJ Research PTC-200 (Boston, MA). Amplicons were resolved as a single band by 1.5% agarose gel electrophoresis before single-strand conformation polymorphism (SSCP) analysis to ensure that a specific single product was amplified. All PCR products were subject to SSCP analysis performed on 7% acrylamide/bisacrylamide (49:1 ratio) gel. The SSCP gels were neutralized and silver stained to visualize the SSCP patterns and the images were transferred to the computer with a 1200 dpi scanner (Fig. 1A, B). The samples which have different electrophoretic mobilities were analyzed further by nucleotide sequence analysis (Fig. 2A, B).
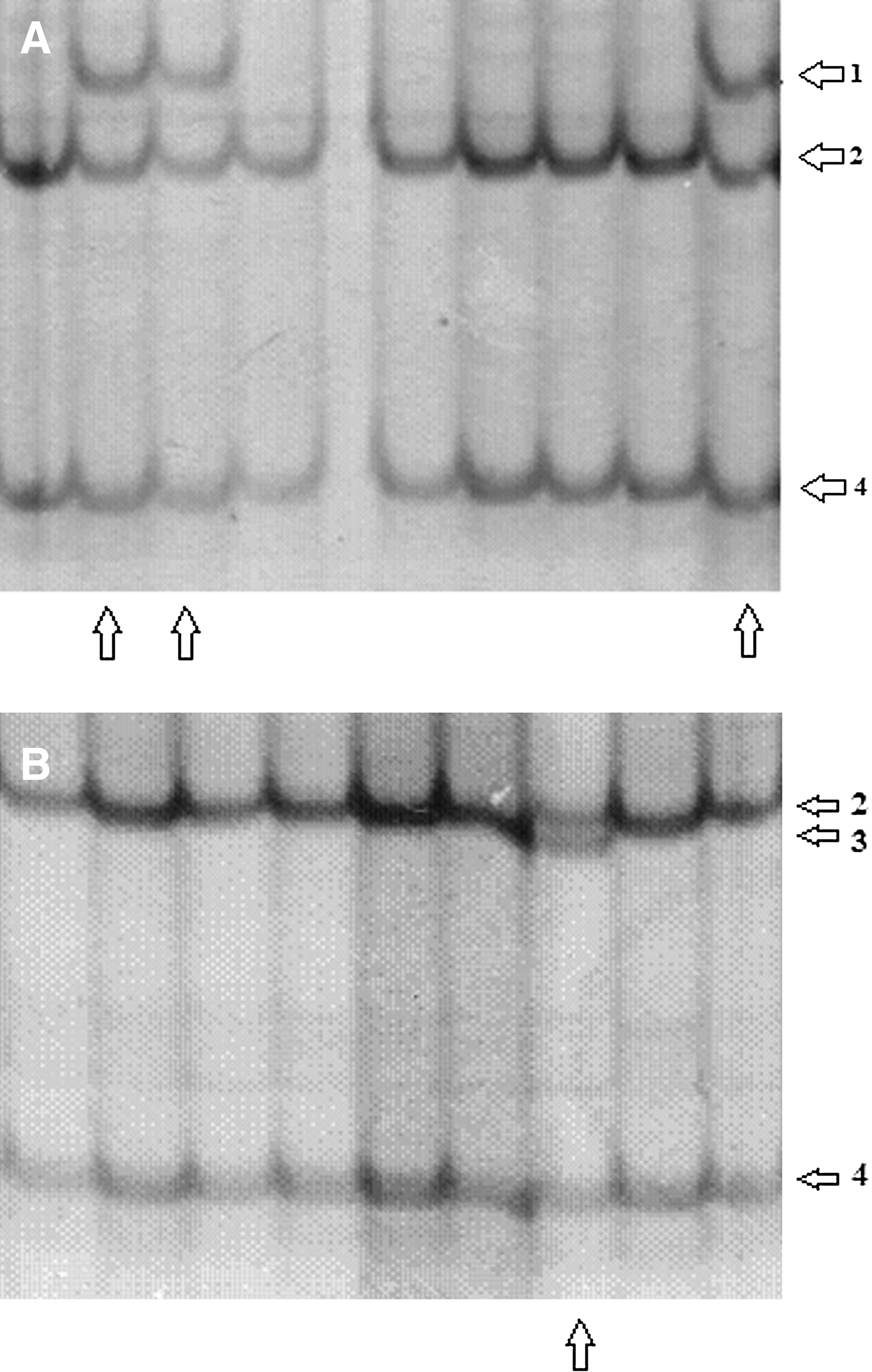
The single-strand conformation polymorphism (SSCP) acrylamide gel images of the S3P1 region of the SOCS3 gene
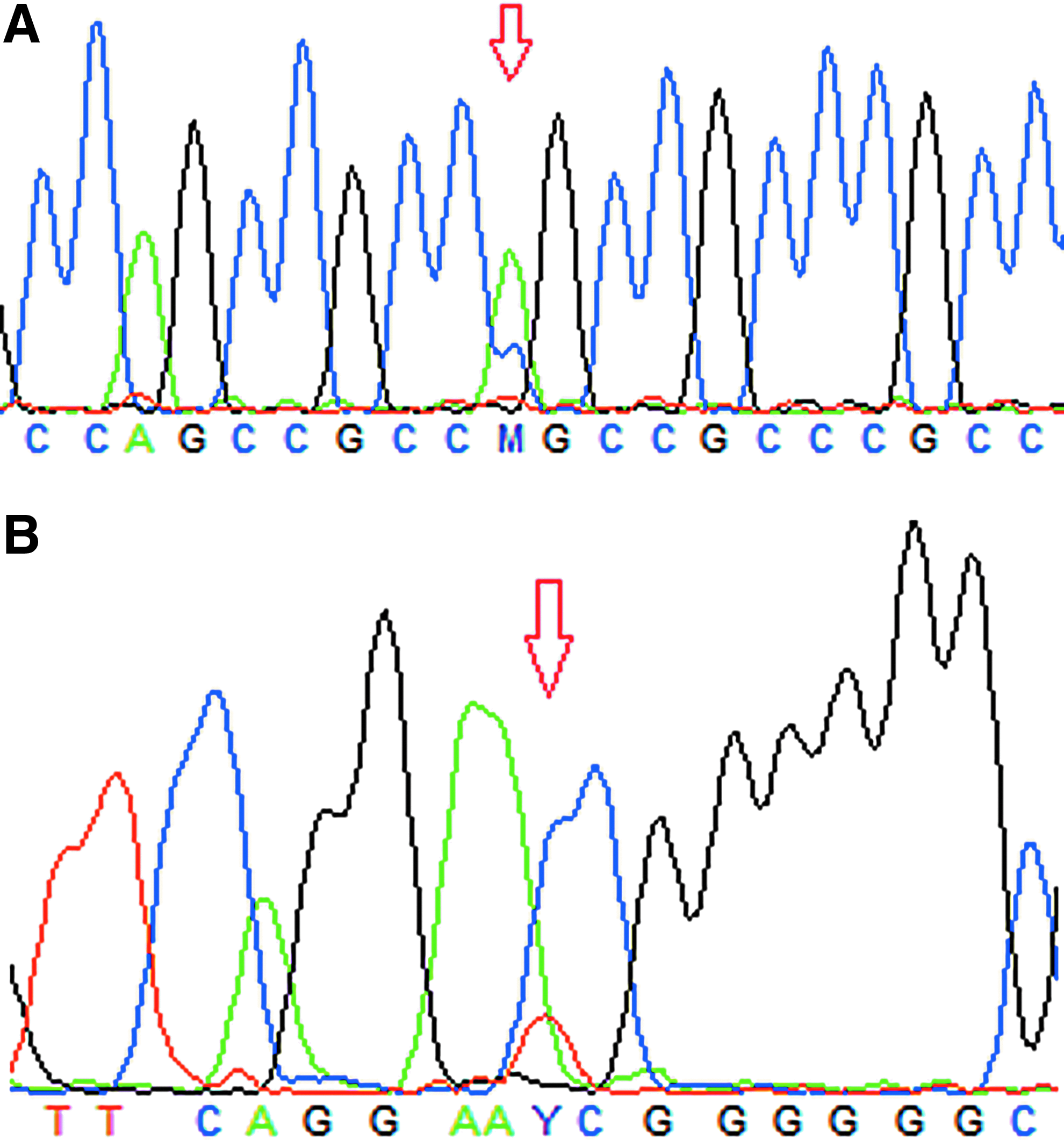
Nucleotide sequence analysis graphic of SSCP band 1 revealed C-914A polymorphism (rs12953258)
PCR, polymerase chain reaction.
Statistical analyses
Results are expressed as the mean±SD or percentage. Statistical analysis was performed using GraphPad Instat (version 3.05; GraphPad Software, San Diego, CA). The chi-square test was used for calculation of the significance of differences in genotype and allele frequencies. The unpaired Student's t-test was used for comparisons of the differences between mean values of two groups. All statistical tests and p-values were two-sided, and p<0.05 was considered statistically significant.
Results
SOCS3 gene promoter regions were PCR amplified and screened for mutations using SSCP analysis. The results revealed that the chosen parts of the promoter region of SOCS3 gene, except for the S3P1 region, were highly conserved in both patients and healthy control groups, and showed no differences. The S3P1 region is highly polymorphic and displayed three genotypes. The SSCP banding pattern gave 1,2,4, 2,3,4 and 2,4 genotypes. The distribution of these genotypes between the groups is shown in Table 2. The genotypes distribution showed that the 2,4 genotype is more common than the 1,2,4 and 2,3,4 genotypes (Table 2). These results are the first showing the distribution of the SOCS3 gene promoter polymorphisms in metastatic CRC patients and a control group. Interestingly, 2,3,4 genotype was observed in 3 metastatic CRC patients, but not in controls. There were no statistically significant differences in genotype frequencies between the groups (p>0.05). Nucleotide sequence analysis of SSCP bands 2 and 4 showed that these were wild-type. Moreover, the sequence analysis revealed that the nucleotides positioned at −914 and −1031 nt had the polymorphisms. Nucleotide sequence analysis of SSCP band 1 and band 3 revealed C-914A (rs12953258) and T-1031C (rs111033850) polymorphisms, respectively. The T-1031C polymorphism lies in the border of STAT-binding site (Fig. 3). T-1031C polymorphism (rs111033850) is a newly identified single nucleotide polymorphism (SNP) with this study, and we submitted this to NCBI database (www.ncbi.nlm.nih.gov/projects/SNP/snp_viewBatch.cgi?sbid=1050750).

Nucleotide sequence of the 5′ flanking region of the human SOCS3 gene. The STAT binding sites are boxed with gray color and the start codon ATG is defined as +1 (He et al., 2003b). The SNPs observed in our study, rs111033850 and rs12953258, are shown in boxes.
p=0.1310, Chi-square=4.065, Degrees of freedom=2.
CRC, colorectal cancer.
Discussion
In this study, we have shown that the S3P1 region of the SOCS3 gene was polymorphic, and showed three genotypes. Additionally, we observed that the 2,3,4 genotype was only present in metastatic CRC patients, and identified a new SNP, T-1031C (rs111033850), which lies in the border of STAT-binding site. To the best of our knowledge, this is the first study to examine the association of the SOCS3 gene promoter polymorphisms with the risk of metastatic CRC development. However, we could not detect any significant association between these promoter polymorphisms and patients group in this study.
STAT3 signaling participates in regulating cell growth and survival during oncogenesis. Constitutive activation of STAT proteins is frequently seen in a variety of human tumors. Cytokine stimulation activates the JAK-STAT pathway, and STAT activation itself represents an important step in the transcriptional activation of SOCS3 gene expression. Several studies have investigated the role of the SOCS protein family in the oncogenesis of several types of solid and haematological tumours indicating an important role of SOCS protein family in the tumour cellular growth and differentiation (He et al., 2003a; Oshimo et al., 2004; Sutherland et al., 2004; Watanabe et al., 2004; Weber et al., 2005). SOCS3 may also be involved in the suppression of tumour growth and metastasis of several malignancies, including lung cancer, breast cancer, hepatocellular cancer, and head and neck squamous cell carcinoma (Adams et al., 1998; He et al., 2004; Weber et al., 2005; Evans et al., 2007; Cui et al., 2008). A favorable role of SOCS3 as predictor of earlier tumour stage and better prognosis and clinical outcome in breast cancer has been reported (Sasi et al., 2010). In addition to its physiologic role in inhibiting cytokine signaling, SOCS3 may also function as an important tumor suppressor gene by blocking the transforming activity of oncogenic forms of STAT3.
Several studies related to the role of SOCS3 in the oncogenesis suggest that SOCS3 acts as a tumor suppressor. More recent studies showed that overexpression of SOCS3 markedly suppressed STAT3 expression, and inhibited STAT5 phosphorylation, resulting in decreased cell proliferation in T47D breast cancer cell lines, and decreased proliferation and anchorage-independent growth in MCF7 breast cancer cell lines (Barclay et al., 2009). An increase in RNA levels of SOCS3, and elevated expression of SOCS3 immunoreactive proteins have recently been reported in tumor tissue and reactive stroma from breast cancer patients (Raccurt et al., 2003). Recent evidence in humans suggests that the SOCS3 protein may act as a tumor suppressor in the colon (Li et al., 2010). Emerging data suggest that SOCS3 may act as a tumor suppressor in the intestine, and decreased SOCS3 expression may promote CRC. Intestinal epithelial cells-specific silencing of SOCS3 leads to a dramatic increase in tumor load in a mouse model of inflammation-associated CRC (Rigby et al., 2007). When SOCS3 mRNA levels were examined in normal mucosal biopsies of colonoscopy patients, it was found that there was no difference in median SOCS3 values between individuals with or without adenoma (Hamilton et al., 2010). Constitutive activation of STAT proteins has been reported in many cancers, including breast cancer, prostate cancer, leukemia, head and neck cancer, lymphomas, and multiple myeloma, suggesting that aberrant JAK/STAT signaling plays important roles in malignant progression by regulating cell growth and survival (He et al., 2004). Recent studies also show that STAT3 activation is increased, and SOCS3 is silenced in tumors of patients with ulcerative colitis-associated and sporadic CRC (Lin et al., 2005; Li et al., 2010). In various cancers, inhibition of STAT signaling suppresses cancer cell growth and induces apoptosis (Raccurt et al., 2003; Sutherland et al., 2004). In tumours, dysfunction of SOCS molecules can cause hyper-responsiveness to cytokines and growth factors and could contribute to the development and/or progression of malignant tumours.
One important mechanism is SOCS gene promoter methylation which may result in SOCS gene silencing and subsequent loss of negative feedback control on the JAK/STAT signaling pathway in tumour cells (Sutherland et al., 2004; Watanabe et al., 2004; Weber et al., 2005). Silencing of SOCS3 genes by methylation of the promoter has been correlated with the loss of growth control inhibition in lung carcinomas (He et al., 2003a). A similar epigenetic block apparently affects SOCS3 gene expression in head and neck carcinomas (Weber et al., 2005). A hypermethylated SOCS3 promoter was found in Barrett's adenocarcinomas, and in high and low grade intraepithelial neoplasias (Tischoff et al., 2007). Methylation of the SOCS3 promoter correlated with downregulation of SOCS3 transcripts and protein expression in these tumours (Tischoff et al., 2007). These studies indicated that aberrant silencing could be a cause for constitutive activation of JAK/STAT pathway in cancer cells. SOCS3 silencing is one of the mechanisms for constitutive STAT3 activation. Conversely, demethylation of SOCS genes promoters may result in restoration of SOCS mRNA and protein expression, and lead to a decrease in activated STAT3 levels which in turn results in induction of apoptosis and suppression of growth (Sutherland et al., 2004). On the other hand, there is also evidence that SOCS3 was not hypermethylated in ovarian and breast cancer cell lines (Sutherland et al., 2004). It is likely that SOCS activation pathways operate differently in diverse cell and tissue types.
Reduced expression of SOCS3 has been observed in various human cancers and is associated with constitutive STAT3 activation (Yoshimura, 2006). Indeed, the levels of SOCS3 were inversely correlated with STAT3 activation in regions of human livers with and without HCC (Ogata et al., 2006). Numerous studies have shown that hyperactivation of STAT3 can contribute to tumourigenesis by inducing multiple tumour-promoting genes (Yoshimura, 2006). Constitutive STAT3 activation in tumour cells contributes to an expansion of tumour cells by promoting cell proliferation, survival, angiogenesis, and tissue remodeling. However, other studies also indicated that malignant cells compared to their normal counterparts express SOCS genes constitutively (Sakai et al., 2002; Li et al., 2004; Evans et al., 2007) and forced expression of SOCS transgene often conferred resistance to IFN. Constitutive expression can potentially hamper immunotherapy by inactivating cytokine signals. SOCS3 proteins may down-regulate the biological response of melanoma cells to endogenous or exogenously administered interferons. It has been shown that overexpression of SOCS3 led to a significant reduction in IFN-induced STAT1 phosphorylation and gene expression, and siRNA inhibition of SOCS3 expression in melanoma cells enhanced their responsiveness to interferon stimulation (Lesinski et al., 2010). Clearly, many uncertainties remain on the function of SOCS in tumourigenesis.
Mutations leading to the loss of SOCS activity may give rise to cytokine hyperresponsiveness and may contribute to the development of diseases such as diabetes and cancer (Starr and Hilton, 1998). Since SOCS proteins inhibit cytokine signal transduction pathways, mutations in SOCS genes would be expected to lead to prolonged cytokine signalling. The promoter region of the human SOCS3 gene is about 1.1 kbp in length and consists of two adjacent STAT-binding sites, a G/C-rich element, and a putative TATA box (He et al., 2003b). This 5′ genomic region has high basal promoter activity and is responsive to growth factors and cytokines (He et al., 2003a, 2003b). One polymorphism (T-1031C) found in this study lies in the border of the STAT-binding site. It can be hypothesized that this polymorphism is likely to influence STAT-binding and SOCS3 activity.
In conclusion, our findings suggest that the SOCS3 gene promoter polymorphisms were not involved in the risk of developing metastatic CRC in our Turkish population. Studies in larger populations are needed to confirm these results in metastatic CRC, and further studies are required to verify these findings in different ethnic groups.
Footnotes
Author Disclosure Statement
No competing financial interests exist.