
Abstract
Aims: Human placental lactogen (hPL) acts via the prolactin receptor (PRLR) on maternal β-cells to mediate increases in β-cell mass and function during normal pregnancy. This interaction between hPL and PRLR is essential to maintain normal glucose homeostasis and to address the increased metabolic demands of pregnancy. Given the importance of the PRLR-hPL axis in pancreatic islet cell adaptation to pregnancy, we hypothesized that genetic variation in the PRLR gene could influence risk of development of gestational diabetes mellitus (GDM). DNA samples from 96 mothers affected by GDM and 96 unaffected cases were genotyped for 8 selected single nucleotide polymorphisms (SNPs) in PRLR. Results: Significant associations were identified in two SNPs analyzed. The minor alleles of PRLR SNPs rs10068521 and rs9292578 were more frequently observed in GDM cases than controls and were associated with a 2.36-fold increased risk for GDM in those carrying the minor allele. Conclusion: SNPs of the PRLR gene 5′ UTR and promoter region are associated with increased risk for GDM in a population of Chilean subjects.
Introduction
I
The human PRLR gene is encoded on chromosome 5p13.2 and consists of eleven exons. Multiple promoters allow for variable transcriptional regulation and alternative splicing that generate several different isoforms of the PRLR (Hu et al., 2002), which vary in the number of exons contained and, therefore, in the composition of the extracellular domain, intracellular domain, or both (Binart et al., 2010). The long isoform of the receptor, which is strongly expressed in pancreas, is activated by hPL to initiate the Jak2/STAT5 signaling pathway, resulting in increased insulin secretion and β-cell division (Binart et al., 2010).
Given the evidence for an important role of the PRLR in the adaptation of pancreatic islet cells to pregnancy and the complexity of PRLR products that can be generated by alternative splicing, we hypothesized that genetic variation in the PRLR gene could influence the risk to develop GDM. Consequently, identification of single nucleotide polymorphisms (SNPs) in the PRLR gene that are associated with GDM could illuminate potential genetic predisposition toward GDM and provide targets for further investigation to improve our understanding of the mechanisms controlling islet cell biology during the unique physiology of pregnancy.
Materials and Methods
Subjects
A case-control study of Chilean women was performed using samples from the clinical database and bank of biological samples from the Perinatology Research Branch (Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH, DHHS). All participants received obstetrical care at a single hospital in Santiago, Chile. All women provided written informed consent before sample collection. The use of clinical data, collection, and utilization of maternal and neonatal blood for research purposes was approved by the Institutional Review Boards of the Sótero del Río Hospital, the Eunice Kennedy Shriver National Institute of Child Health and Human Development, NIH. All samples were de-identified prior to our analysis.
The classification criteria for normal pregnant women included the following: no prior diabetes mellitus or other metabolic conditions; no obstetrical, maternal or fetal complications during pregnancy; normal plasma glucose concentrations in the first trimester; normal oral glucose tolerance test (OGTT), as defined by World Health Organization criteria, in the third trimester; and delivery at term of a healthy neonate with an appropriate birth weight for gestational age (between 10th and 90th percentile) (Mazaki-Tovi et al., 2009). Classification of GDM was based on World Health Organization (WHO) criteria of fasting plasma glucose ≥126 mg/dL (≥7.0 mM) or plasma glucose ≥140 mg/dL (≥7.8 mM), measured 2 h after a 75-g OGTT administered between 24 and 28 weeks gestational age (Alberti and Zimmet, 1998). Sample collection and DNA extraction were performed as previously described (Hill et al., 2011).
SNP genotyping
SNP analysis was performed using real-time allelic discrimination TaqMan assays (Applied Biosystems) with modifications. All PCR reactions contained 10 ng of DNA, 5.0 μL GTXpress Master Mix (2×) (Applied Biosystems), 0.25 μL TaqMan Genotyping Assay (Applied Biosystems) (40×), and water for a final volume of 10.0 μL. Real-time PCR was performed on an ABI 7500 Fast Real-Time PCR Machine (Applied Biosystems) under the following conditions: a single activation cycle of 95°C for 20 s, followed by 40 cycles of amplification (95°C for 3 s and 60°C for 30 s). For each cycle, the software determined the fluorescent signal from the VIC- or FAM-labeled probe (Applied Biosystems).
Statistical analysis
Genotype frequencies in GDM cases and controls were tested for Hardy-Weinberg equilibrium (HWE), and any difference between the observed and expected frequencies was tested for significance using the Chi-square test. In instances where the expected number of cases was smaller than five, Fisher's exact test was used. Odds ratios (ORs) and 95% confidence intervals (CIs) were calculated for associations between genotypes and GDM. Allele frequencies from the control groups were used to calculate the ORs of having gestational diabetes.
Armitage's trend test with one degree of freedom was performed to evaluate the trend across the genotypes. Any p-value less than 0.05 was considered to be significant. Statistical analyses were performed using JMP Version 9 (SAS Institute, Inc.) and the HWE test tool offered by the Institute of Human Genetics, Technische Universität München (Munich, Germany; http://ihg.gsf.de/cgi-bin/hw/hwa1.pl). Genotype frequencies were also compared with available minor allele frequencies available at the National Center for Biotechnology Information (NCBI) Single Nucleotide Polymorphism Database, dbSNP build 137, updated June 26, 2012 (www.ncbi.nlm.nih.gov/snp). Testing for linkage disequilibrium between SNPs with significant ORs for association with GDM was performed with WGAViewer, http://compute1.lsrc.duke.edu/softwares/WGAViewer/, using data from the International HapMap Consortium (Ge et al., 2008).
Results
Characteristics of the GDM cases and controls are shown in Table 1 and are consistent with known GDM risk factors and outcomes. As expected, both higher maternal body mass indexes (26.8 kg/m2 vs. 25.5 kg/m2, p=0.0433) and infant birth weights (3582 g vs. 3420 g, p=0.0041) were observed in the GDM cases compared with controls. Maternal age was higher for gestational diabetes cases than controls (32.1 years vs. 26.4 years, p<0.001), and GDM cases also experienced a higher rate of Cesarean-section delivery (32.3% vs. 13.5% in controls, p=0.0032).
Bold numbers indicate p<0.05.
GDM, gestational diabetes mellitus.
To assess the relationship between selected PRLR SNPs and GDM risk, we examined eight SNPs distributed throughout the promoter regions, introns, and untranslated regions of PRLR (Fig. 1, Table 2). For SNP rs10068521, located in the 5′ UTR of the PRLR gene (Fig. 1), the minor allele C was detected in more than twice as many GDM cases as controls (20.2% vs. 9.7%). Total minor allele content, or proportion of the number of minor alleles observed out of the total alleles present at each locus, was 10.1% in GDM cases, versus 4.8% of controls. This difference between the groups translated to a statistically significant OR of 2.36 (95%CI, 1.01-5.54; p=0.0435) to develop GDM among those carrying the minor allele C as opposed to the major allele G (Table 3).
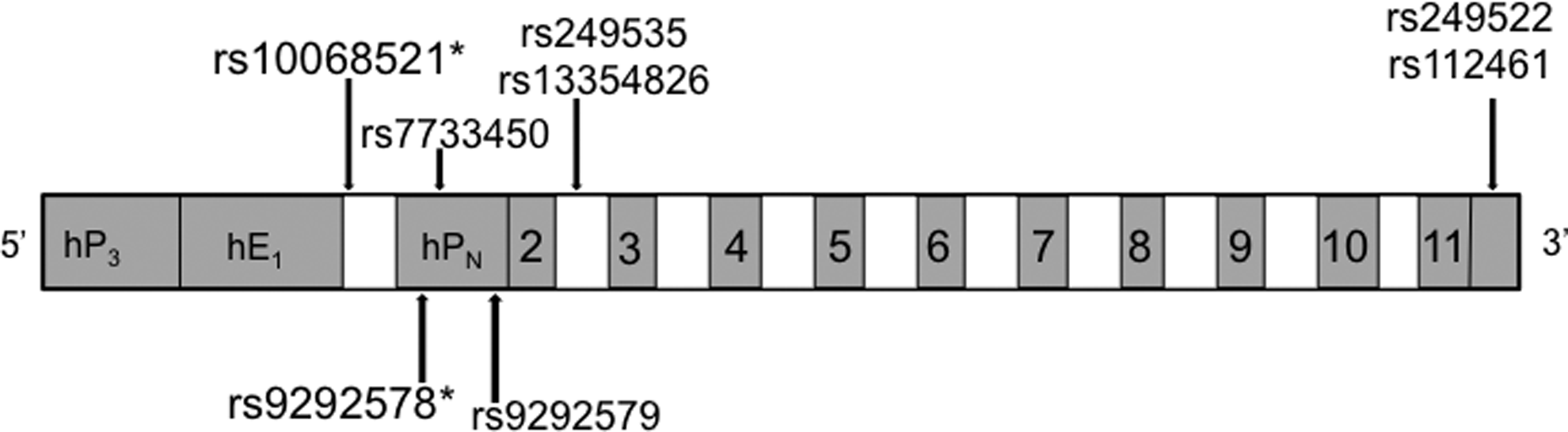
Gene structure of the PRLR gene. A schematic of the PRLR gene is shown. Numbered boxes indicate exons and open boxes indicate introns. Promoters are indicated as hP3, hE1, and hPN. Single nucleotide polymorphisms are indicated by the arrows. *p<0.05 for association with gestational diabetes mellitus. PRLR, prolactin receptor.
PRLR, prolactin receptor; SNP, single nucleotide polymorphism.
Bold numbers indicate p<0.05.
CI, confidence interval; OR, odds ratios.
For SNP rs9292578, located within the promoter HPN of the PRLR gene (Fig. 1), the minor allele A was also detected in approximately twice as many GDM cases compared with controls (19.1% vs. 9.7%). Total minor allele content was 10.6% in GDM and 4.8% in controls, which supports a statistically significant OR of 2.36 to develop GDM among those carrying the minor allele A as opposed to major allele C (p=0.0352, Table 3).
For the SNP rs9292579, also found in the promoter HPN of the PRLR gene (Fig. 1), the minor allele A was also detected in about twice as many GDM cases compared with controls (18.5% vs. 9.7%). Total minor allele content was 10.6% in GDM and 4.8% in controls. For this SNP, the difference between the groups approached but did not achieve statistical significance, with an OR of 2.12 (95%CI 0.89-5.02, p=0.0850) to develop having GDM among those carrying the minor allele A as opposed to major allele C (Table 3).
It is interesting to note that in all samples, whether cases or controls, each sample that was genotyped to contain the minor allele of rs10068521 also contained the minor allele of rs9292578, strongly suggesting that these SNPs are in linkage disequilibrium. Testing for linkage disequilibrium based on available data for populations with European, Han Chinese, Japanese in Tokyo, and Yoruba African populations results in r2 (correlation coefficient) and D′ (standardized disequilibrium coefficient) values of 1.000 and D′ values of 1.000 for all populations, indicating that these SNPs are highly likely to be linked in these groups. No associations were found for the other five SNPs analyzed, which were located in the 3′ UTR or intronic regions.
The ethnic background of our sample population from Chile is heterogeneous, and there were no specific minor allele frequencies reported for these SNPs in this particular population. However, for each of our three identified SNPs, comparison of the minor allele frequencies in our control patients correlate well with data from the international 1000 Genomes Project (1000 Genomes Project Consortium, 2010), which includes populations with ancestry from Europe, East Asia, South Asia, West Africa, and the Americas. For rs10068521 and rs9292578, the minor allele was present in 4.8% of our control cases and in 4.7% of the 1000 Genomes Project subjects. For rs9292759, the minor allele was present in 4.8% of control cases compared with 5.1% (1000 Genomes Project Consortium, 2010).
Discussion
Data from both animal models and human islet cells studied in vitro support the importance of the PRLR in glucose homeostasis during pregnancy. Rats with hyperprolactinemia induced via implantation of extra anterior pituitary glandular tissue demonstrated improved glucose tolerance despite serum insulin levels similar to control rats (Adler and Sokol, 1982). Recombinant human prolactin (rhPRL) has been shown to improve glucose-stimulated insulin secretion in cultured human pancreatic islet cells (Labriola et al., 2007) and may be protective against apoptosis in these isolated human pancreatic islets (Terra et al., 2011). Prlr+/− mice demonstrated impaired glucose disposal and insulin release during pregnancy, along with higher postprandial glucose (Huang et al., 2009). Of note, the Prlr+/+ offspring of these haploinsufficient mice had higher nonfasting glucose levels than the Prlr+/+ offspring of the wild-type mothers, suggesting that maternal genotype may still cause alterations of glucose metabolism through a still undetermined mechanism, even in genotypically normal offspring (Huang et al., 2009).
The human PRLR gene has multiple promoters, including hP3 and hPN, which allow for differential and tissue-specific transcriptional regulation, thereby producing several different isoforms of the PRLR protein (Hu et al., 2002). Exon 1 exists in several variant forms (Hu et al., 2002) and is noncoding, while exons 3 through 11 are coding exons. The human PRLR is a single transmembrane-spanning protein belonging to a superfamily of growth hormone/prolactin/cytokine receptors, and alternative splicing results in different isoforms of the PRLR (Binart et al., 2010), each with different effects on intracellular signaling, that may be stimulatory or inhibitory. The long isoform of the receptor, which is strongly expressed in pancreas, consists of exons 3-10; the hPL protein acts through this long isoform to activate the Jak2/STAT5 signaling pathway, thereby increasing insulin secretion and β-cell division (Binart et al., 2010). With regard to control of β-cell apoptosis downstream of the Jak2/STAT5 pathway, studies of rhPRL in cultured human islet cells suggest that lactogen treatment increases levels of the anti-apoptotic protein BCL2 and decreased levels of pro-apoptotic caspases (Terra et al., 2011).
There are currently three strategies useful for prevention of type 2 diabetes in women with prior GDM: maintenance of a healthy weight, use of insulin sensitizing medications, and lactation (Ramos-Roman, 2011). In large cohorts of women included in the Nurses' Health Studies and the prospective Coronary Artery Risk Development in Young Adults (CARDIA) study, longer duration of lactation was associated with a lower risk of subsequent type 2 diabetes, even several years after cessation of breastfeeding (Stuebe et al. 2005; Gunderson et al. 2010). According to the “reset hypothesis” postulated by Stuebe and Rich-Edwards, lactation promotes a return to the cardiometabolically favorable prepregnant state (Stuebe and Rich-Edwards, 2009). It is also possible that the proliferative and anti-apoptotic effects of lactogens on pancreatic β-cells could be a contributing factor to a more rapid and complete return to normal physiology.
Our results suggest that the SNPs rs10068521 and rs9292578 are significantly associated with GDM in a population of Chilean subjects. These SNPs, which appear to be in linkage disequilibrium and were, thus, transmitted together in our GDM cases, are located within noncoding regions of the PRLR gene. Based on their locations, the most likely mechanism responsible for increased GDM risk would not be due to any protein coding alterations in the PRLR itself but through effects on transcriptional regulation or efficiency. Potential mechanisms to explain the effect of the region of PRLR containing these SNPs include alteration to the nearby H34Kme3 and H3K27me3 histone modification sites, CHD2 and TBP transcription factor-binding sites, or high CpG methylation sites (UCSC ENCODE Browser/database, Human—February, 2009, GRCh37/h19 Assembly, http://genome.ucsc.edu/, Rosenbloom et al., 2013). Additionally, further studies of variants in the CSH1 gene, which encodes hPL, could clarify the contributions of each component of the hPL-PRLR interaction to development of GDM.
Relevant to our study, the Chilean population is comprised of admixture between the Amerindian (aboriginal) populations and Europeans of Spanish descent; while the overall percentage of aboriginal admixture has been estimated at 40%, there is a smaller percentage of aboriginal admixture noted in groups with higher socioeconomic status, and vice versa (Cifuentes et al., 2004). Although the genetic heterogeneity of our subjects is a potential limitation of our study, the control subjects were ascertained from the same institution and would therefore be expected to be of similar genetic background. Further examination of these SNPs in other, better-defined ethnic populations would provide greater insight into the ethnicity-specific implications of our results. Additionally, consideration of genetic markers of Caucasian admixture, as was done in studies of type 2 diabetes mellitus in American Indians (Knowler et al., 1988), may aid in correcting for the effect of these SNPs in this Chilean population or in studies of other ethnically mixed populations.
In conclusion, we report that SNPs of the PRLR gene 5′ UTR and promoter region are associated with increased risk for GDM in a population of Chilean subjects. The exact mechanics for the influence of these SNPs on PRLR transcription remains to be elucidated. Given the alarming rate of increase of T2DM worldwide, strategies to prevent or reduce T2DM risk are sorely needed from a public health perspective, especially in those ethnic populations with genetic predispositions toward T2DM development. Awareness of an individual's increased GDM risk, with its associated higher risk of subsequent T2DM, could help direct measures aimed to support breastfeeding towards those who are most likely to derive lifelong health improvements from doing so.
Footnotes
Acknowledgments
This study was supported by the A.D. Williams Fund, Grant Number 648973, of the Virginia Commonwealth University, the VCU CTSA UL1RR031990, and in part by the Division of Intramural Research, National Institute of Child Health and Human Development, National Institutes of Health.
Author Disclosure Statement
No competing financial interests exist.