
Abstract
Aim: Many case-control studies have been performed in the past to elucidate the association between the P2X7 receptor 1513 A>C (rs3751143) polymorphism and tuberculosis (TB) risk. However, their data interpretation was difficult due to scattered and inconsistent results that led to limited power. In this study, a quantitative summary assessment has been done through meta-analysis to appraise the association between the 1513 A>C polymorphism and TB susceptibility. Methodology: Systematic assessment was performed for the published studies related with the association between the P2X7 1513 A>C polymorphism and TB risk retrieved from PubMed (Medline), EMBASE search. A meta-analysis was done using a statistical program to evaluate the said association. Pooled odds ratios (ORs) and 95% confidence intervals (95% CIs) were calculated for allele contrast, homozygous, heterozygous, dominant, and recessive genetic models. Results: A total of 2710 controls and 2521 TB cases were included in this meta-analysis. Meta-analysis results showed that C allele carrier status was significantly associated with increased TB risk (C vs. A: p=0.001; OR=1.382, 95% CI=1.248-1.531). Significant risk of TB was associated with the homozygous mutant CC (CC vs. AA: p=0.001; OR=1.676, 95% CI=1.251-2.247) and heterozygous AC (AC vs. AA: p=0.001; OR=1.429, 95% CI=1.260-1.621) comparisons. Similarly, dominant (CC vs. AA+AC: p=0.008; OR=1.481, 95% CI=1.109-1.978) and recessive (CC+AC vs. AA: p=0.001; OR=1.458, 95% CI=1.292-1.645) genetic models also revealed increased risk of developing TB. Conclusion: We found that the P2X7 1513 A>C gene polymorphism is significantly associated with increased susceptibility to TB. Also, future well-designed epidemiological studies with stratified case-control and biological characterization may be beneficial to validate these findings.
Introduction
In case of TB infection, the macrophages act both as the principal reservoir of M. tuberculosis infection and as the primary effector cells regulating bacterial growth and viability (Kaufmann, 2001). It has been well recognized that ATP plays a significant role in initiating the antimycobacterial activity occurring with human macrophages, exerting its effect through the P2X7 cell surface receptor (Lammas et al., 1997). The human P2X7 (purinergic receptor P2X, ligand-gated ion channel 7, localized on chromosome 12q24) is an ATP-gated cation channel, which is highly expressed in macrophages (Rassendren et al., 1997) that are infected with Mycobacteria, resulting in the induction of apoptosis (Ferrari et al., 1999). The P2X7 response to ATP involves the calcium-dependent and phospho-lipase D-dependent effector pathway, which encourages mycobacterial death via promotion of phagosome-lysosome fusion (Fernando et al., 2007).
Variations in macrophage response to ATP have been noticed in healthy individuals, which suggest that genetic heterogeneity contributes to P2X7 expression or signaling function (Lammas et al., 1997). Nearly 10% of healthy individuals exhibit a reduced or adverse response toward ATP, a percentage similar to those who develop clinical disease after M. tuberculosis infection (Bloom and Small, 1998). This implies that there might be a pivotal association between host susceptibility to TB and loss of P2X7 function. Previous findings proved that various single-nucleotide polymorphisms in the P2X7 gene lead to loss of receptor function (Tekin et al., 2010). The most common of these is the A1513C polymorphism, where the glutamic acid at position 496 changes to alanine rendering the receptor close to nonfunctional (Saunders et al., 2003; Fernando et al., 2007).
Keeping the functional significance of this genetic variant in view, the impact of A1513C polymorphism on the susceptibility of TB has been studied in different populations, but the findings remain conflicting rather than conclusive (Table 2). Inconsistencies in earlier studies can be attributed to the sample size, ethnic diversity, and low power to detect overall effects due to individual studies. A comprehensive analysis based on different studies will provide strong evidence of the association of the P2X7 1513 A>C polymorphism with TB risk. A meta-analysis is a strong tool for analyzing cumulative data from research studies, where individual sample sizes are small and bear low statistical power (Cohn and Becker, 2003). In this study, we have presented the updated meta-analysis, including the latest data to investigate the association of the P2X7 1513 A>C polymorphism and TB risk.
Materials and Methods
Identification and eligibility of relevant studies
Systematic literature search was performed through PubMed (Medline), EMBASE web databases covering all research articles published with a combination of the following key words: “P2X7 gene AND (polymorphism OR mutation OR variant) AND tuberculosis” (last updated on February, 2013). Studies showing potential relevance for genetic association were evaluated by examining their titles and abstracts. All published studies matching with the above said eligible criteria were retrieved and included in the meta-analysis.
Inclusion and exclusion criteria
To minimize heterogeneity and facilitate the proper elucidation of results, studies included in the present meta-analysis study had to meet all the following criteria: (1) evaluation of the P2X7 A1513C A>C and TB risk, (2) use of case-control design, (3) recruitment of pathologically confirmed TB patients and TB-free controls, (4) have available genotype frequency in case and control, and (5) published in the English language. In addition, when the case-control study was included by more than one article using the same case series, we selected the study that included the largest number of individuals. The major reasons for exclusion of studies were (1) overlapping of data and (2) case-only studies and review articles.
Data extraction and quality assessment
For each publication, the methodological quality assessment and data extraction were independently abstracted in duplicate by two independent investigators using a standard protocol. Data accuracy was ensured using a data-collection form according to the inclusion criteria listed above. In case of disagreement on any item of the data collected from the retrieved studies, the problem would be fully discussed to reach a consensus. Characters abstracted from the studies included the name of first author, the year of publication, the country of origin, sources of cases and controls, the number of cases and controls, types of study, and genotype frequencies.
Statistical analysis
Pooled odds ratios (ORs) and their corresponding 95% confidence intervals (CIs) were calculated to evaluate the relation between the 1513 A>C polymorphism and TB risk. Heterogeneity assumption was examined by the chi-square-based Q-test (Wu and Li, 1999). The significance level (p-value) >0.05 for the Q-test indicated a lack of heterogeneity among the studies. Pooled ORs were calculated either by the fixed-effects model (Mantel and Haenszel, 1959) or by the random-effects model (DerSimonian and Laird, 1986). In addition, I2 statistics were employed to quantify interstudy variability and ranged between 0% and 100%, where a value of 0% indicates no observed heterogeneity and larger values indicate an increasing degree of heterogeneity (Higgins et al., 2003). The Hardy-Weinberg equilibrium (HWE) in the control group was measured via the chi-square test. Funnel plot asymmetry was estimated by the Egger's linear regression test, which is a linear regression approach to measure the funnel plot asymmetry on the natural logarithm scale of the OR. The significance of the intercept was determined by the t-test (p-value<0.05 was considered as representation of statistically significant publication bias) (Egger et al., 1997a). All statistical analysis for this meta-analysis was performed by the comprehensive meta-analysis (CMA) V2 software (Biostat). The CMA V2 has some advantages over other programs available for computing meta-analyses. A comparison of meta-analysis programs can be retrieved through http://meta-analysis.com/pages/comparisons.html.
Results
Characteristics of published studies
A total of 22 articles were retrieved through literature search from the PubMed (Medline) and the EMBASE database. All retrieved articles were examined by reading the titles, abstracts, and the full texts for the potentially relevant publications. Articles were further checked for their suitability for this meta-analysis (Fig. 1). In addition to the database search, the reference lists of the retrieved articles were also screened for other potential relevant articles. Studies comprising the P2X7 polymorphism to predict survival in TB patients or considering the P2X7 variant as an indicator for response to therapy were excluded. Studies related to investigation of the levels of P2X7 mRNA or protein expression or review articles were also excluded. Strict criteria were followed in article search, only case-control or cohort design studies having frequency of all the three genotypes were included. Following the careful screening and strict inclusion and exclusion criteria, 12 eligible original published studies were found and included in the study (Table 1). A detailed flowchart of the selection process is shown in Figure 1. Distribution of genotypes, HWE p-values in the controls, and susceptibility to TB has been tabulated in Table 2.

Flow diagram of included and excluded studies.
HB, hospital based; PB, population based; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism.
HWE, Hardy-Weinberg equilibrium; TB, tuberculosis; ETB, extrapulmonary tuberculosis; PTB, pulmonary tuberculosis.
Publication bias
The Begg's funnel plot and Egger's test were performed to evaluate the publication bias among the included studies for this meta-analysis. The appearance of the shape of funnel plots and the results of Egger's test have shown the evidence of publication bias for all comparison models (C vs. A, CC vs. AA, AC vs. AA, and CC+AC vs. AA) except dominant contrast (CC vs. AA+AC) (Table 3). The trim and fill method was employed to adjust the publication bias for recomputing the effect size (Duval and Tweedie, 2000).
Test of heterogeneity
The Q-test and I2 statistics were utilized for test of heterogeneity among the selected studies. No heterogeneity was observed in any of the models, that is, allele (C vs. A), homozygous (CC vs. AA), heterozygous (AC vs. AA), dominant (CC vs. AA+AC), and recessive (CC+AC vs. AA) genotype models, which were included for the analysis. Hence, the fixed model was employed for calculating the combined OR and 95% CI for all comparisons (Table 3).
Meta-analysis of P2X7 1513 A>C polymorphism and TB susceptibility
All 12 studies were pooled together, which resulted into 2710 controls and 2521 TB cases, and the fixed-effects model (based on heterogeneity) was used to assess the overall association between the P2X7 1513 A>C polymorphism and risk of TB. The pooled data indicated an evidence for a significant association between the P2X7 1513 A>C polymorphism and susceptibility to TB in all the models. Variant allele C demonstrated significant risk of developing TB in terms of the frequency with wild allele (A) comparison (C vs. A: p=0.001; OR=1.382, 95% CI=1.248-1.531). Also, the homozygous mutant CC (CC vs. AA: p=0.001; OR=1.676, 95% CI=1.251-2.247) and heterozygous AC (AC vs. AA: p=0.001; OR=1.429, 95% CI=1.260-1.621) genotypes revealed significantly increased risk for the occurrence of TB as compared with the wild-type homozygous AA genotype (Fig. 2). Moreover, analysis of dominant (CC vs. AA+AC: p=0.008; OR=1.481, 95% CI=1.109-1.978) and recessive (CC+AC vs. AA: p=0.001; OR=1.458, 95% CI=1.292-1.645) genetic model indicated a 1.4-fold increased risk of developing TB (Fig. 3).
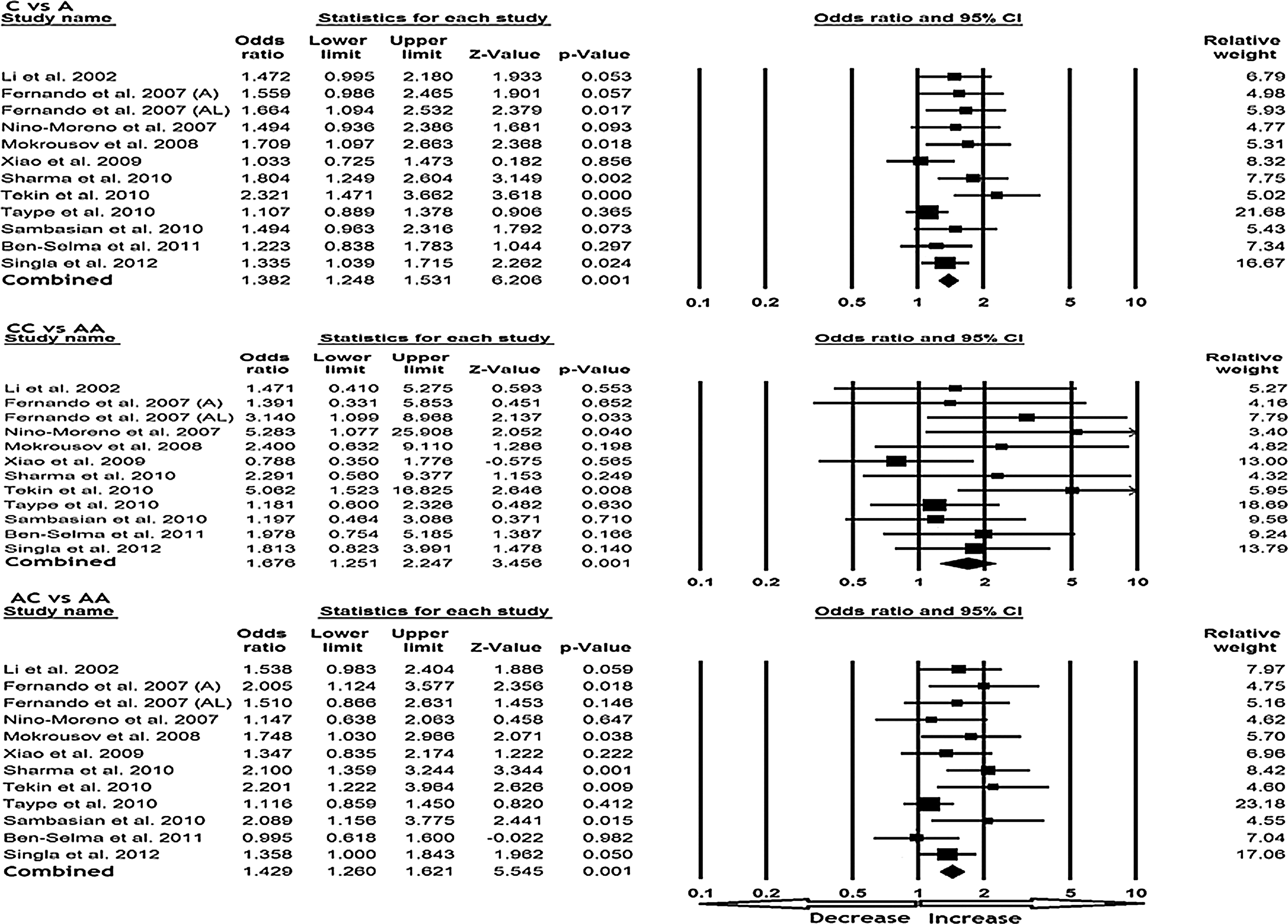
Forest plot of overall tuberculosis (TB) risk associated with the P2X7 1513 A>C polymorphism. The squares and horizontal lines correspond to the study-specific odds ratio (OR) and 95% confidence interval (CI).
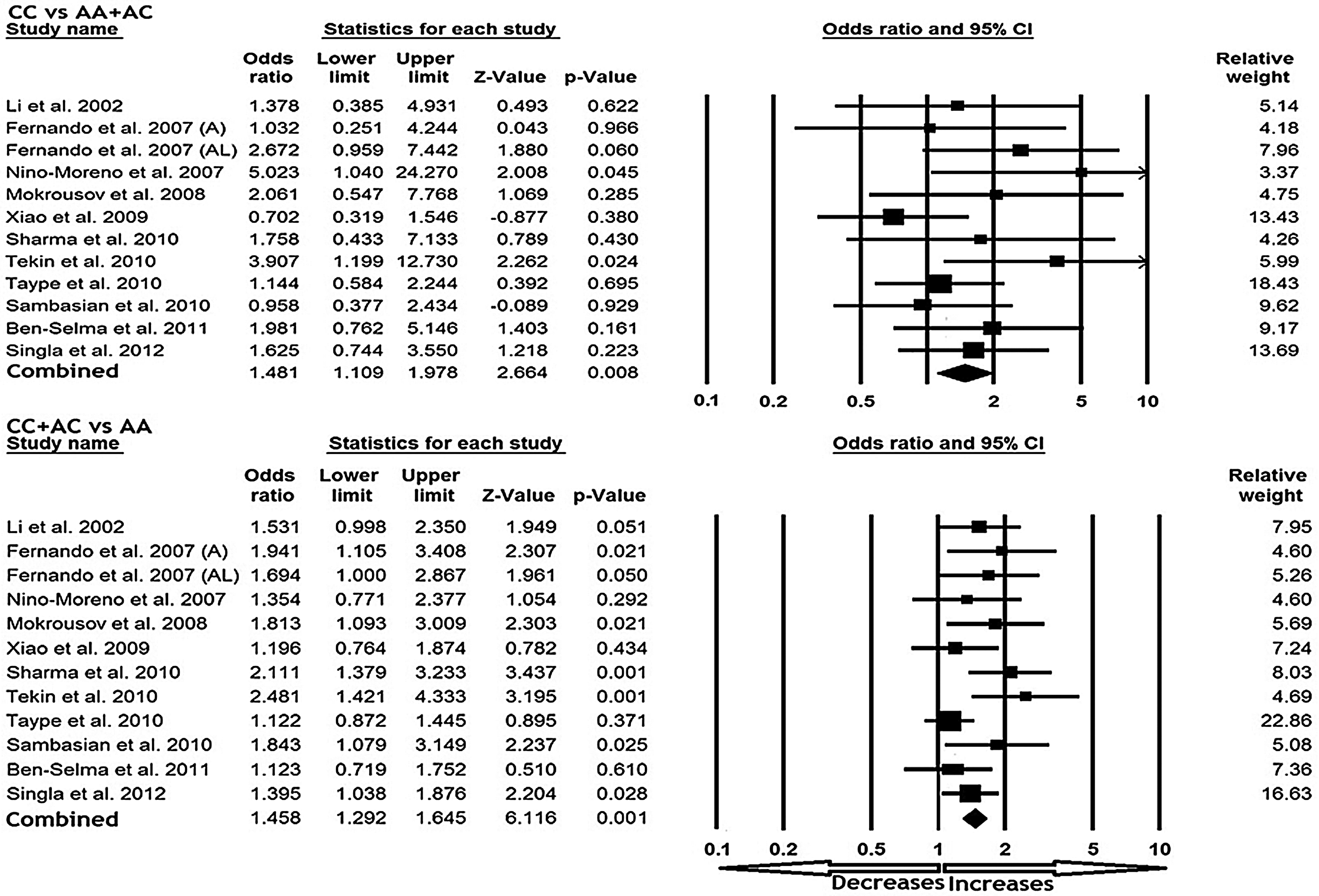
Forest plot of overall TB risk associated with the P2X7 1513 A>C polymorphism. The squares and horizontal lines correspond to the study-specific OR and 95% CI.
Discussion
Earlier studies proved that TB susceptibility is determined not only by the infectious agent and environmental factors, but also by the host genetic factor (Bellamy, 2003). As a result, a number of candidate genes have been investigated to access the probable association between modulations of TB risk across different populations. Despite stating in detail about the recruitment criteria, sample size, characteristics of participants, and genotyping methods, most of the studies lack a proper conclusion. Hence, to improve the statistical power and to determine the effect size of the P2X7 1513 A>C polymorphism, a meta-analysis has been done with updated data of 12 studies to determine a more concrete correlation between the P2X7 1513 A>C polymorphism and the occurrence of TB risk. Meta-analysis data showed that polymorphism of the P2X7 gene is significantly associated with increased TB risk in the general population. Subjects with a C allele and two variant CC alleles had 1.3- and 1.6-fold increased risk of developing TB as compared with the wild A allele and homozygous AA genotype, respectively. Similarly, heterozygous, dominant, and recessive models have shown increased risk of TB. Based upon the above data and importance of P2X7's role in the pathogenesis of TB, it is biologically plausible that the 1513 A>C polymorphism may modulate the risk of TB and could be a genetic factor for interindividual differences in susceptibility to TB disease.
Through previous reports, we know that P2X7 is a ligand-gated cation channel with a trimeric structure that is expressed in the cell membrane and activated mainly by ATP (Rassendren et al., 1997). Activation of the P2X7 receptor leads a cascade of intracellular events, opening of cation channels and pores, activation of phospholipase D (Kusner and Adams, 2000), metalloproteases (Gu and Wiley, 2006), and enhances the intracellular calcium with the resulting induction of apoptosis and death of both the host cell and the internalized bacilli via stimulation of caspases (Ferrari et al., 1999). Earlier studies have proved that ATP-induced apoptosis and subsequent bacterial killing were inhibited after the treatment of infected macrophages with the P2X7 antagonist KN62 (Lammas et al., 1997). Furthermore, it has been also noticed that knockout mice of the P2X7 gene were unable to affect the killing of mycobacteria by the addition of ATP (Fairbairn et al., 2001). The most common A>C substitution in exon 13 at position 1513′ has been shown to abolish P2X7-mediated apoptosis of macrophages and assists the growth of the pathogen, apparently predisposing the host to TB infection (Gu et al., 2001). Individuals homozygous for the 1513 C allele have complete P2X7 receptor function loss, whereas heterozygosity for this polymorphism significantly impaired function to kill intracellular parasites such as M. tuberculosis compared with individuals with the wild-type A allele (Fernando et al., 2007). Most of the studies included in this meta-analysis showed that the 1513 C allele is associated with occurrence of TB (Table 2).
In the recent past, two authors have also tried to decipher the association of the 1513 A>C polymorphism and TB susceptibility on the basis of meta-analysis, but their findings were inconsistent and indecisive rather than conclusive possibly due to a limited number of studies, low case-control studies, and lack of power (Xiao et al., 2009; Wang et al., 2011). Xiao et al. (2009), reported that the 1513 C allele was significantly associated with TB susceptibility, whereas Wang et al. (2011) failed to produce significant risk to pulmonary TB. In comparison with their studies, the present study has some advancements due to the inclusion of some newly published studies based on the 1513 A>C polymorphism, which led to a greater sample size, strict study selection, and refined data extraction that assists in derivation of a better conclusion.
The genetic susceptibility to TB is polygenic (Moller and Hoal, 2010); hence, a single genetic variant is usually inadequate to predict the risk of this deadly disease. The most important property of this gene polymorphism is that their incidence can vary substantially between different racial or ethnic populations. Before reaching a conclusion, limitations of this meta-analysis should be addressed, that is, first, studies published only in the English language were included; second, studies indexed by the selected electronic databases (PubMed, EMBASE) were included for data analysis; it is possible that some relevant articles might be published in other languages and indexed in other databases, which may be missed; third, the data were not stratified by other factors such as HIV status and TB severity due to inadequate information in the primary articles; fourth, we failed to include the gene and environmental interactions because of the lack of information in the primary studies.
In conclusion, meta-analysis is a highly cost-effective method that combines both statistically significant and nonsignificant findings from individual similar studies and develops a definitive conclusion (Egger et al., 1997b). This meta-analysis evaluated the relationship between the P2X7 1513 A>C polymorphism and TB risk and suggested that the 1513 A>C polymorphism appeared to be associated with TB susceptibility. However, future well-designed large studies might be beneficial to validate this association in different populations, including consideration of environmental factors responsible for the susceptibility to TB.
Footnotes
Author Disclosure Statement
No competing financial interests exist.