
Abstract
Germinal mosaicism should be considered when estimating the recurrence risk in families with Duchenne/Becker muscular dystrophy (D/BMD). Germinal mosaicism, however, has not been assessed in Mexican families with deletions in the DMD gene. To determine the distribution of deletions in the two hot spots and the proportion of de novo and transmitted deletions, we analyzed 153 individuals with D/BMD and a DMD partial deletion and 322 of their maternal female relatives. Predilection for the distal hot spot was observed in 112 families (73%), while gene dosage analysis of female relatives of D/BMD patients identified germinal mosaicism deletions in at least 11.6% of the patients' families, thought to result from de novo mutations. Recurrence risk due to germinal mosaicism justifies carrier detection in maternal female relatives and prenatal diagnosis in mothers of individuals with apparently de novo DMD deletions.
Introduction
D
The DMD gene has shown a wide mutational spectrum, although partial deletions represent 60-65% of all mutations. These deletions occur preferentially in two hot spots: the distal region between exons 44 and 53 (70-80%) and the proximal region between exons 2 and 20 (20-30%) (Den Dunnen et al., 1989; Koenig et al., 1989; Prior and Bridgeman, 2005; Yang et al., 2013). Haldane's theoretical model of de novo mutations in X linked diseases predicts that, in families with only one affected individual, 67% of mothers would be carriers (Haldane, 2004). The probability of the mother being a carrier has been linked to the type of mutation found in the proband. Haldane's prediction was corroborated when a duplication or point mutation was responsible for the disease, with two-third of the mothers being carriers. However, when deletions were responsible, only 40% of the mothers were found to be heterozygotes (Bakker et al., 1989; Sinha et al., 1996; Alcántara et al., 1999). Furthermore, combined analysis has shown that only 27% of mothers with a single affected son resulting from a deletion were carriers (Taylor et al., 2007).
Proximal deletions have been found more frequently in familial cases, whereas distal deletions were more frequent among single cases, a difference that may be due to variations in the chromatin structure and stability during development (Passos-Bueno et al., 1992). This would favor proximal deletions during the early stages of embryogenesis, with a consequent increase in transmission risk, with distal deletions occurring during later stages of embryogenesis, reducing the risk of recurrence and therefore being more frequent among single cases.
Germinal mosaicism is a phenomenon that must be considered during genetic counseling, since it modifies theoretical recurrence risks, particularly in cases in which an apparent de novo mutation has been identified (Bakker et al., 1987; Wood and McGillivray, 1988; van Essen et al., 1992). Estimations of recurrence risk should also consider other factors, including the risk haplotype, mutation type and location, which are included in specific algorithms (Passos-Bueno et al., 1992; Fischer et al., 2006; Helderman-van den Enden et al., 2009). For D/BMD, the recurrence risks for single cases with de novo deletions were 14% if the risk haplotype was known and 7% if it was not known (Bakker et al., 1989). These results were replicated in a larger cohort of single cases with de novo deletions, resulting in recurrence risks of 8.6% when the risk haplotype was known and 4.3% when it was not (Helderman-van den Enden et al., 2009).
Previously, we studied 76 families of nonrelated Mexican patients with D/BMD, finding that de novo deletions were present in 62.2% (23 of 37) of patients with deletions (Alcántara et al., 1999), a proportion similar to that observed in other studies (Sinha et al., 1996; Taylor et al., 2007). No cases of germinal mosaicism were found, probably due to the small number of included families or because the family structures did not allow the identification of this phenomenon. We therefore analyzed a larger number of families to determine the frequencies of de novo deletions in the DMD gene and of germinal mosaicism in Mexican families with DMD deletions.
Materials and Methods
The study included 153 families of nonrelated individuals with D/BMD, including 140 with DMD and 13 with BMD, harboring a DMD gene deletion. Deletions were identified through a 22 exon multiplex PCR (PCRm) (promoter and exons 3, 6, 8, 12, 13, 16, 17, 19, 43-45, and 47-60). To compare the distribution of the DMD deletions, the gene regions were defined as proximal and distal if the majority of deleted exons were between exons 3 and 20 and beyond exon 40, respectively (Helderman-van den Enden et al., 2009). Based on family history alone, 28 patients were classified as familial (FC) and 120 as single (SC) cases; the remaining 5 cases could not be classified because their family history was unavailable.
DMD gene dosage analysis was performed in 322 female relatives of the 153 probands, including the mother of each proband and 169 second and third degree relatives of 89 of the 153 probands (58.2%), including 43 SCs and 46 FCs. Gene dosage analysis was performed by semiquantitative multiplex PCR (PCRsqm) (Abbs and Bobrow, 1992) or real-time PCR (Traverso et al., 2006), by amplifying three exons on a case-specific basis, including two deleted exons (problem exons) and one nondeleted exon (intrasample control) for each proband. Two intersample controls, consisting of DNA samples from one healthy male and one healthy female, and a negative control (no DNA) were included in each assay. PCRsqm was used to genotype 218 samples, including 112 from patients' mothers, whereas triplicate real-time PCR was used to assess the gene dosage in samples from 104 female relatives, including 41 mothers. The relative quantification of DMD exons was determined by the ΔΔCt method (Traverso et al., 2006).
The distributions of distal and proximal deletions according to case type (FC, SC or germinal mosaics) were compared using chi-squared tests.
Results
Of the 153 individuals with deletions in the DMD gene, 30 (19.6%) had deletions in the proximal region, 112 (73.2%) had deletions in the distal region, and 11 (7.2%) had deletions in both. Exons 48, 49, and 50 were most frequently deleted, representing 21.2% of the 813 deleted exons in all 153 patients.
Gene dosage analysis in the patients' mothers showed that 61 (39.9%) were heterozygous for the corresponding deletions and were considered obligate carriers; these patients were therefore classified as FCs. DMD deletions could not be observed in the peripheral blood DNA of the remaining 92 mothers (60.1%). Molecular analysis of these mothers resulted in the reclassification of 32 patients, originally classified by their family histories as being single cases, as having familial disease and one case considered familial was also reassigned as germinal mosaic (Fig. 1). Furthermore, gene dosage analysis of other female relatives identified five cases of germinal mosaicism (Fig. 2). The distribution of the deletions by case type is shown in Table 1.
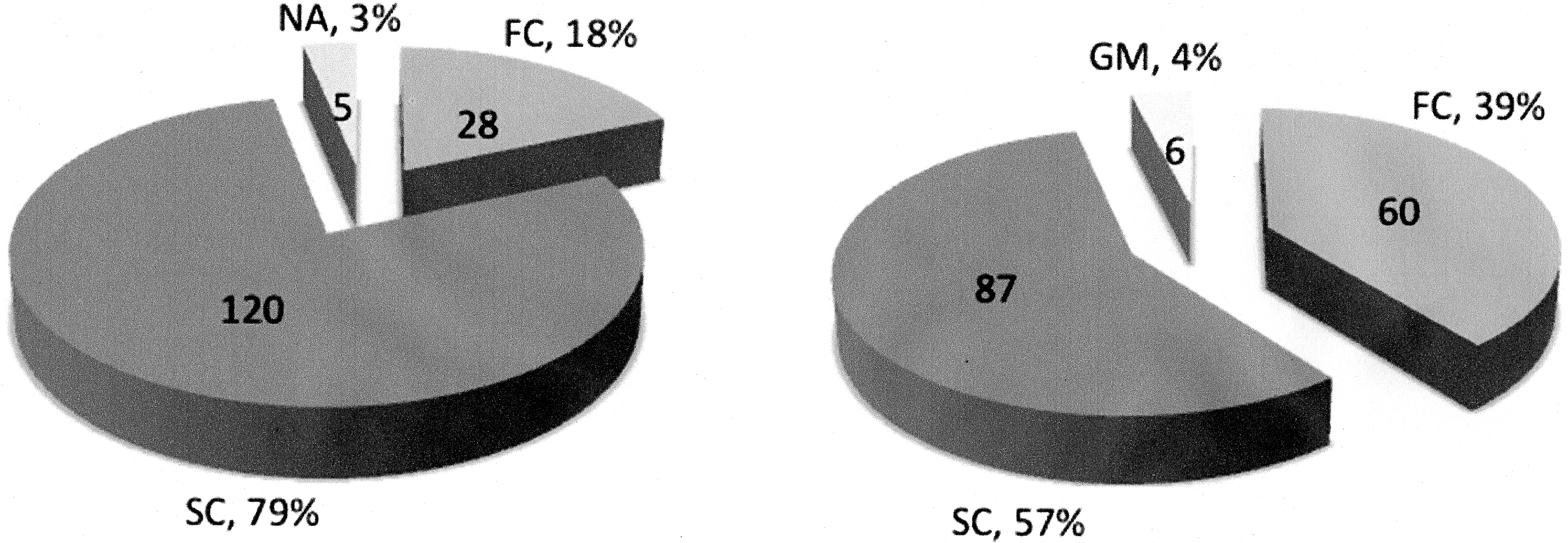
Duchenne/Becker muscular dystrophy (D/BMD) deletion cases identified as single or familial by family history alone and after gene dosage analysis of their mothers and other maternal female relatives. A molecular study allowed the classification of five families with no available family history, with four classified as single and one as familial. Six germinal mosaics were identified, one originally regarded as familial and the others as single cases. NA, family history not available; FC, familial cases; SC, single cases; GM, germinal mosaicism cases.

Genealogies of the identified individuals with germinal mosaicism.
The number and percentage of deletions in each region were determined. Chi-squared tests were performed for each type of case to determine if the frequencies of proximal and distal deletions differed significantly. Distal deletions were more frequent in afamilial cases, with χ2=45.32 when p<0.01 and for bsingle cases, with χ2=40.7 when p<0.01.
Proximal deletions were more frequent in germinal cases, but the difference was less striking, with χ2=8.37 when p<0.05.
DMD, Duchenne muscular dystrophy.
This deletion distribution analysis did not include 11 cases in which extensive deletions were found, since this type of deletion had not been considered previously (Passos-Bueno et al., 1992; Helderman-van den Enden et al., 2009). Table 2 shows that there were no differences in the distribution of deletions between FCs and SCs, in which distal deletions were predominant, while highlighting the difference in deletion distribution between these two types and mosaic cases (Fig. 3).
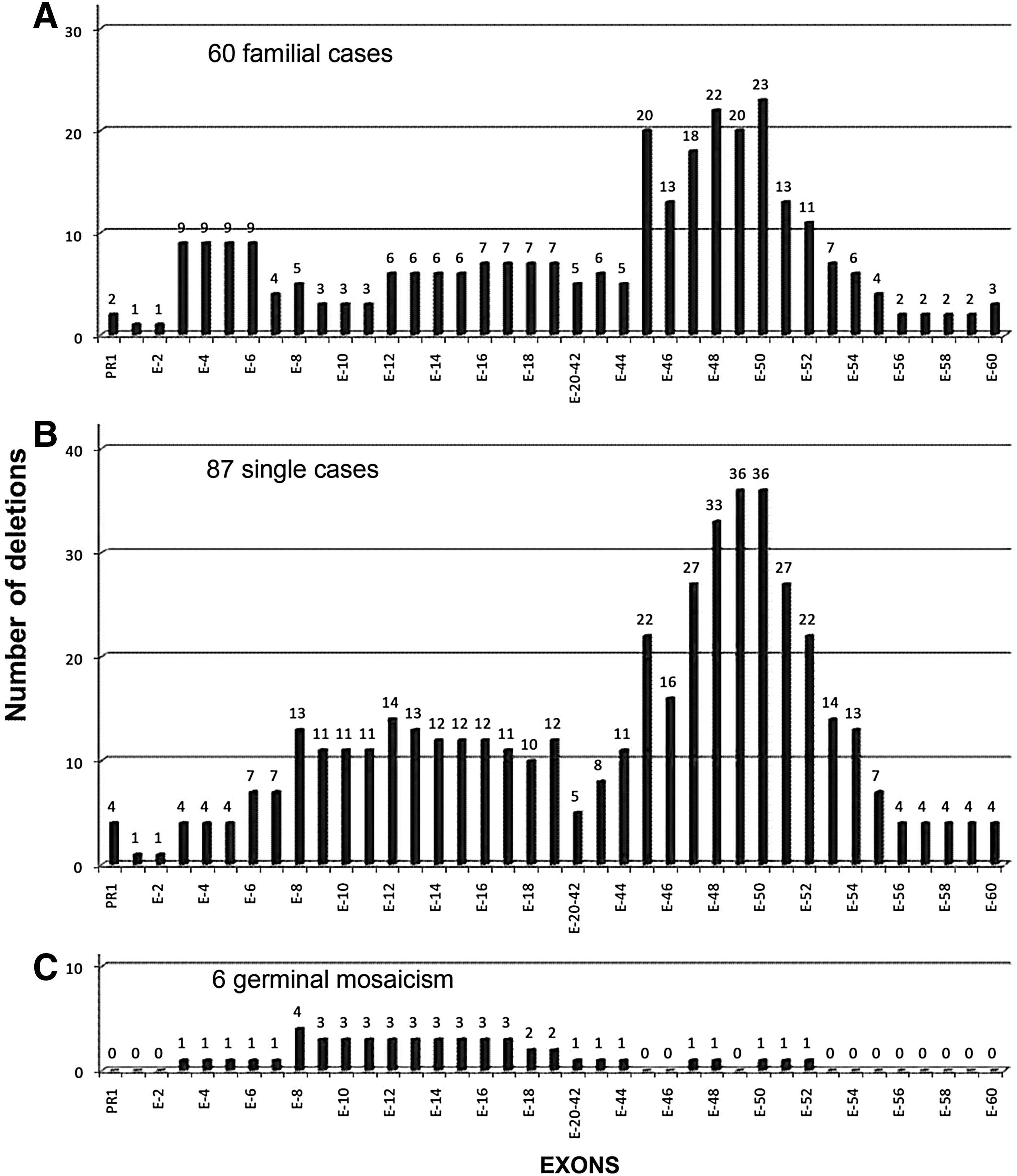
Distribution and frequency of each deleted exon studied in the
Statistically significant.
Discussion
Our preliminary study found that DMD deletions arise as de novo events in 62.2% of patients (Alcántara et al., 1999). Increasing the cohort to 153 D/BMD families with confirmed deletions resulted in a frequency of de novo deletions of 60.1%, similar to our previous result (p=0.25) and to the reports of other cohorts. The nonrandom distribution of these deletions in two hot spots is also in agreement with results in other populations, with distal deletions between exons 45 and 50 being observed more frequently, in particular, deletions affecting exons 48-50 represent 21.2% of all deleted exons (Den Dunnen et al., 1989; Love et al., 1991; Passos-Bueno et al., 1992; Shomrat et al., 1994; Florentin et al., 1995; Sinha et al., 1996; Alcántara et al., 1999; Mukherjee et al., 2003; Prior and Bridgeman, 2005; Traverso et al., 2006; Wang et al., 2008; Helderman-van den Enden et al., 2009). MLPA has been recognized as the optimal initial test for genotyping D/BMD patients, due to its ability to analyze the dosage of every exon in a single assay. Use of this method in a cohort of over 1000 D/BMD patients resulted in the identification of nearly 60% of DMD deletions, most between exons 3 and 22 and exons 45 and 54, with very few deletions found between exons 56 and 79. Moreover, the exons most frequently deleted were exons 50, 49, and 48, in that order (Yang et al., 2013). These findings suggest that our 22 exon multiplex PCR-based strategy covers practically all the exons most frequently deleted on MLPA. Moreover, our strategy can be regarded as an acceptable alternative for identifying DMD deletions, although this strategy would be unable to identify partial duplications.
No significant differences between proximal and distal distributions of deletions were observed in the FC and SC groups, with distal deletions being more frequent for both. Similar findings were observed in two other studies, one including 157 deletion cases (Grimm et al., 1994) and the other including 57 (Florentin et al., 1995). These findings differ from those of another study, which found that, among 234 deletion cases, proximal deletions were more frequent for FCs (Passos-Bueno et al., 1992). Moreover, that study reported that proximal deletions had a higher probability of being familial, a finding not supported by our results.
Extending gene dosage analysis to first, second, and third degree female relatives in 43 families identified 5 germinal mosaics, indicating that, in at least 11.6% of families originally classified as SC due to a de novo deletion, the mutation was transmitted through germinal mosaicism. This finding is in agreement with recurrence risk estimates when the risk haplotypes are known and unknown of 14% and 7%, respectively (Bakker et al., 1989), and of 20% and 10%, respectively (van Essen et al., 1992). The germinal mosaic frequency may have been underestimated since in 49 of the 92 families of SCs due to a de novo deletion, second and third degree female relatives of probands were unavailable for gene dosage analysis.
Recurrence risk may depend on deletion location. In one study of seven germinal mosaics with a deletion, six were found to be proximal. An analysis of haplotype transmission showed that the recurrence risks were 30% for proximal deletions and 4% for distal deletions (Passos-Bueno et al., 1992). Another study of 14 germinal mosaic cases with a deletion found that 8 were distal and 6 were proximal, with transmission analysis showing risks of recurrence of 15.6% for proximal and 6.4% for distal deletions (Helderman-van den Enden et al., 2009). Our extended gene dosage analysis of 58.2% (89/153) of the included families identified five germinal mosaics, with no hot spot predominance. We were unable to determine recurrence risks because we did not perform haplotype transmission analysis.
In conclusion, these findings support the well-established notion that deletions occur more frequently in the distal region of the DMD gene. Increasing the number of patients in our cohort, relative to our previous report, showed that the frequency of de novo deletions was maintained near 60%. Moreover, assessments of female relatives resulted in the identification of germinal mosaicism cases, showing no preference for either hot spot. Recurrence risk of germinal mosaicism cases in D/BMD families reveals the importance of gene dosage analysis of maternal female relatives of D/BMD single cases, even when maternal analysis of genomic DNA from peripheral blood indicates a noncarrier status for a DMD deletion. Furthermore, the frequency of this phenomenon suggests that prenatal molecular diagnoses be offered to mothers negative for the deletions found in their sons and even to other maternal female relatives of D/BMD SCs.
Footnotes
Acknowledgment
This work was supported by Research Funding from the National Institute of Pediatrics.
Author Disclosure Statement
All the authors declare that no competing financial interests exist.