
Abstract
Objectives: Some polymorphisms of the interleukin-16 (IL-16) gene are associated with various cancers. To resolve inconsistencies in published data, we performed a meta-analysis of studies of IL-16 polymorphisms and cancer risk. Materials and Methods: Seven eligible studies pooling 1678 cases and 1937 controls were quantitatively analyzed to evaluate three IL-16 polymorphisms (rs4778889, rs4072111, rs11556218) and cancer risk. Hardy-Weinberg equilibrium (HWE) for controls was evaluated by goodness-of-fit chi-squared tests. Odds ratios (ORs) and their 95% confidence intervals (CIs) were calculated for each genetic model and allelic comparison. Data were pooled using fixed- or random-effects models depending on heterogeneity across studies. Results: Our meta-analysis demonstrated that the IL-16 polymorphism rs11556218 was significantly associated with increased susceptibility to cancer in several models, including allelic contrast (OR=1.307; 95% CI, 1.108-1.541), heterozygote contrast (OR=1.650; 95% CI, 1.424-1.911), and dominant model (OR=1.605; 95% CI, 1.391-1.845). The result remained consistent after adjustment for age and gender. No significant association was found between IL-16 polymorphisms rs4778889 rs4072111 and cancer risk. Conclusions: The rs11556218 T/G polymorphism of the IL-16 gene was significantly associated with elevated cancer risk in Asian populations. Our results warrant larger, better-designed studies, including a greater ethnic variety.
Introduction
C
Interleukin (IL)-16 is a proinflammatory cytokine that has a wide array of functions in multiple immunopathobiological processes (Yellapa et al., 2012). The gene encoding for IL-16 is located on chromosome 15q 26.1-3 (Cruikshank et al., 2000). IL-16 is initially expressed as a precursor protein and then activated by caspase 3 through cleavage of the 631 amino acid proprotein to form a C-terminal 121-amino acid peptide (Center et al., 1996; Chupp et al., 1998). The cellular source for IL-16 contains a group of inflammatory cells, including cytotoxic T lymphocytes (CD8+ T cells), monocytes/macrophages, dendritic cells, mast cells, epithelial cells, and fibroblasts (Glass et al., 2006; Gao et al., 2009b; Goihl et al., 2012). IL-16 stimulates the production of a variety of proinflammatory cytokines by monocytes with related biological sequelae (Mathy et al., 2000). All of these cytokines, including IL-6, IL-1β, IL-15, and tumor necrosis factor-α, have been demonstrated to be positively related to development and progression of cancer (Gao et al., 2009b). Evidence of increased IL-16 levels in serum or plasma has also been recognized in multiple malignant tumors in both preclinical and clinical trials (Kovacs, 2001; Bellomo et al., 2007; Gao et al., 2009b).
Polymorphisms (single-nucleotide polymorphisms; SNPs) of the IL-16 gene will alter the transcription level and affect the function of IL-16, eventually exhibiting differentiated expression in cancers. Recently, a variety of studies have highlighted some polymorphisms of the IL-16 gene with regard to susceptibility to a wide range of cancers (Gao et al., 2009a, 2009b; Zhu et al., 2010; Azimzadeh et al., 2011, 2012; Li et al., 2011). Three polymorphisms (rs4778889, rs4072111, rs11556218) were substantially explored. However, published data on the association of these polymorphisms and potential cancer risk remain inconsistent, partially because of the comparatively small sample size, the heterogeneity of cancer type, and differences in anthropometric data between cases and controls. To clarify whether the association of IL-16 polymorphisms with cancer risk may contribute to cancer prevention and interventions, we performed a comprehensive literature review and meta-analysis based on eligible published data.
Materials and Methods
Search strategy
We conducted a systematic search of studies addressing the association between IL-16 polymorphisms and cancer risk. Four biomedical electronic databases were retrieved without language limitations (updated on July, 2013): PubMed (US National Library of Medicine), Embase, Web of Knowledge, and CNKI (China National Knowledge Infrastructure). The key words and subject terms involved in the search strategy were as follows: “IL-16,” “interleukin-16,” “polymorphism,” “variant,” “alleles,” “single-nucleotide polymorphisms,” “SNP,” “cancer,” “neoplasms,” “carcinoma,” “tumor.” In addition, potential additional eligible studies were sought through a manual search of citations of included studies. The full search strategy implemented is shown in Fig. 1.
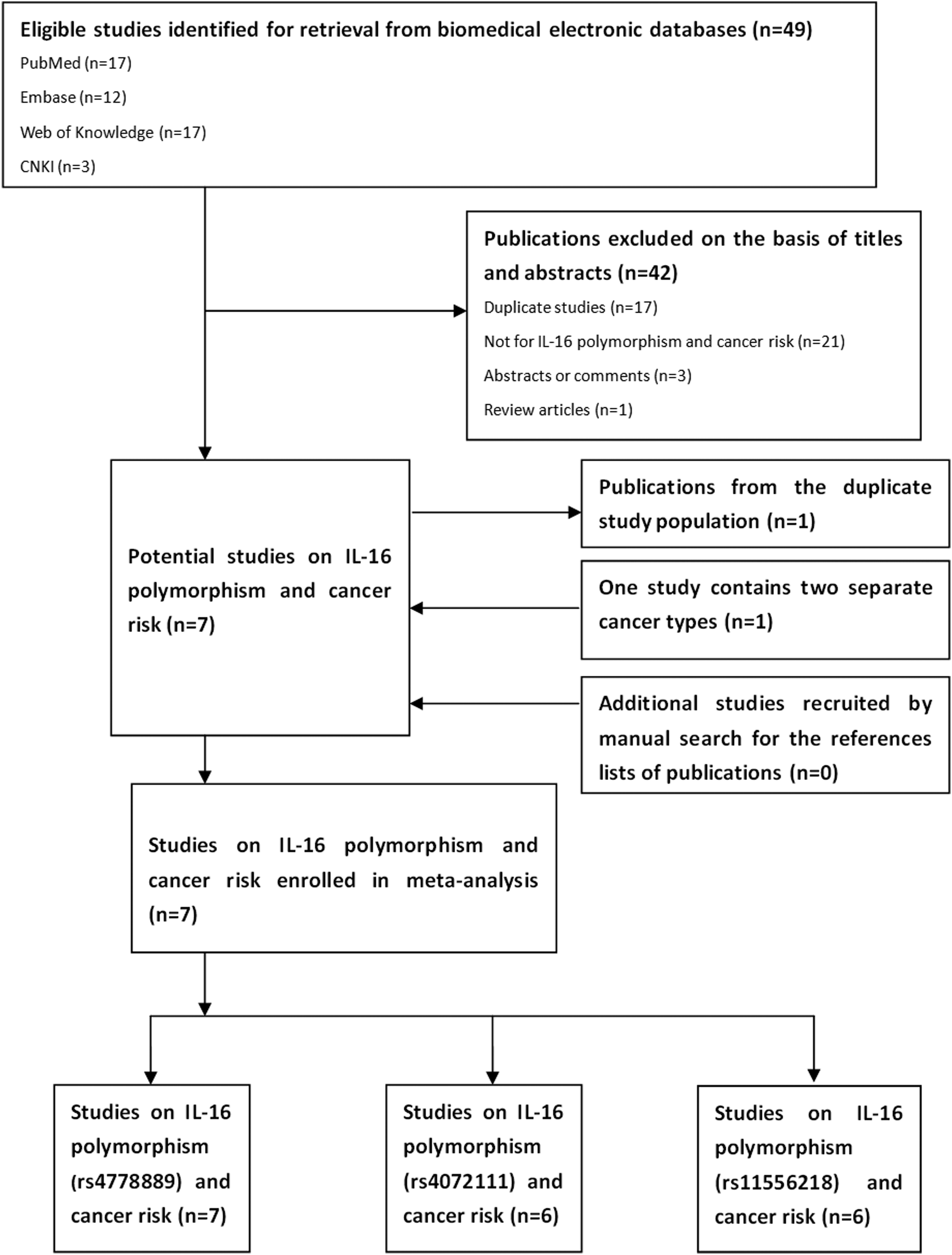
Diagrams for inclusion of studies in this meta-analysis.
Inclusion and exclusion criteria
Inclusion criteria were set to literature selections as follows: first, original case-control or cohort design articles concerning the association between IL-16 polymorphisms and cancers; second, studies should contain sufficient data on genotypes or allelic frequencies; and third, adequate data were available from which to calculate pooled odds ratios (ORs) and their corresponding 95% confidence intervals (CIs). The exclusion criteria included lack of control subjects in studies; article type of abstracts, reviews and comments; and studies not containing adequate information.
Data extraction
Data from all publications complying with the criteria described above were extracted by two authors independently. Any conflicts between evaluations were resolved by discussion until the authors reached a final agreement. For each included study, the following information was extracted: the last name of the first author, year of publication, country and ethnicity of the study population, source of control groups, genotyping method, gender and age distributions, and allele frequency in cases and controls.
Statistical analyses
Statistical analyses were undertaken using STATA version 11.1 (Stata Corp., College Station, TX) with two-sided p-values. Hardy-Weinberg equilibrium (HWE) for the controls was evaluated in each study by a chi-squared test for goodness-of-fit, and p<0.05 was regarded as evidence of inequality of genetic distributions. The minor allele frequency for the controls was also assessed to detect the least common allele occurring in reported populations. Genetic comparisons of extracted data used allelic contrast, homozygote contrast, heterozygote contrast, and dominant and recessive models. ORs and their 95% CIs were measured for each study. The statistical significance for pooled ORs was determined using the Z-test, and pZ<0.05 was considered significant. The Cochran Q statistic was applied to evaluate the heterogeneity across studies (Egger et al., 1997b). When a significant Q statistic was achieved (p<0.1), a random-effects model (the DerSimonian-Laird method) was selected to calculate the pooled ORs (DerSimonian and Laird, 1986); otherwise, a fixed-effects model (the Mantel-Haenszel method) was applied (Donald and Donner, 1987). In this study, we also introduced another measurement, I2, for more quantitative analysis of potential heterogeneity. I2, which stands for the proportion of interstudy variability attributed to heterogeneity, ranges from 0% to 100% (Higgins and Thompson, 2002). High risk of heterogeneity is assured if I2 is larger than 75%. The Egger's linear regression test was conducted to verify the bias of publication (Egger et al., 1997a), and p<0.1 represents the existence of publication bias.
Results
Characteristics of included studies
A total of 49 relevant articles were obtained using the search strategy described above. Among them, 42 were excluded based on titles and abstracts. Of those, 17 studies were duplicated; 21 were not about IL-16 polymorphisms and cancer risk; 3 studies were abstracts of committee reports or comments; 1 was a review article. Then, seven studies were left for full-text review. Further, one study was removed from duplication, another one was treated as two separate studies, for it contained two different cancer types with different case samples (Gao et al., 2009b). Finally, seven studies meeting the inclusion and exclusion criteria highlighted the association of three variants in the IL-16 gene and cancer risk (Table 1). Six were written in English, while the other one was written in Chinese. Among all the studies, seven were pooled for rs4778889 variants; six for rs4072111; and six for rs11556218. All of the studies had a case-control design with a population of Asian origin. Two were for colorectal cancer; two for nasopharyngeal carcinoma; and three for other types of cancer. All of the genotyping used PCR restriction fragment length polymorphism analysis. As shown in Tables 2, 3, and 4, genetic distributions of IL-16 gene polymorphisms are presented. HWE and minor allele frequency for controls were also calculated. Few articles for rs11556218 conformed to HWE; however, most of the articles for rs4778889 and rs4072111 showed goodness-of-fit.
HB, hospital based; PB, population based; NPC, nasopharyngeal cancer; CRC, colorectal cancer; HCC, hepatocellular carcinoma; RCC, renal cell carcinoma; GC, gastric cancer; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; NA, not available.
p<0.05 was regarded as significant.
MAF(%)<5% was regarded as inequality of genetic distributions.
HWE, Hardy-Weinberg equilibrium; χ2, chi-squared; MAF, minor allele frequency.
p<0.05 was regarded as significant.
MAF(%)<5% was regarded as inequality of genetic distributions.
p<0.05 was regarded as significant.
MAF(%)<5% was regarded as inequality of genetic distributions.
Main results of meta-analysis
The detailed results of the association between the IL-16 polymorphism rs4778889 and cancer risk are presented in Table 5. In all genetic models, the overall results of pooled data did not show any statistically significant associations. The pooled results of studies in HWE were consistent with the overall one, for no deviation of HWE was detected.
p<0.05 was regarded as significant.
R, random-effect model; F, fixed-effect model.
As listed in Table 6, no evidence was found in the association of IL-16 polymorphism rs4072111 and cancer risk. Likewise, no statistical associations were revealed in studies restricted to HWE.
p<0.05 was regarded as significant.
As shown in Table 7, evidence from six studies has demonstrated that the IL-16 polymorphism rs11556218 was significantly associated with an increased susceptibility to cancer in several genetic models, including allelic contrast (G vs. T: OR=1.307; 95% CI, 1.108-1.541; pZ=0.001; pHet=0.052), heterozygote contrast (TG vs. TT: OR=1.650; 95% CI, 1.424-1.911; pZ=0.000; pHet=0.17), and dominant model (TG+GG/TT: OR=1.605; 95% CI, 1.391-1.845; pZ=0.000; pHet=0.125). No potential associations were observed in either the homozygote contrast (GG vs. TT: OR=1.209; 95% CI, 0.893-1.638; pZ=0.219; pHet=0.223) or recessive model (GG vs. TT+TG: OR=0.930; 95% CI, 0.698-1.214; pZ=0.622; pHet=0.122). However, among all the studies concerning the rs11556218 polymorphism, only one conformed to HWE. After excluding the other studies, the results of association remained consistent. Statistically significant associations were found in allelic contrast, heterozygote contrast, and the dominant model (p<0.05).
p<0.05 was regarded as significant.
In the subgroup analysis stratified by cancer type, significant risk of colorectal cancer was found for the rs11556218 polymorphism in the heterozygote contrast (TG vs. TT: OR=1.861; 95% CI, 1.491-2.323; pZ=0.000; pHet=0.251) and the dominant model (TG+GG/TT: OR=1.754; 95% CI, 1.412-2.183; pZ=0.000; pHet=0.122). No other significant association was detected for either the rs4778889 or rs4072111 polymorphisms (p>0.05). In the subgroup analysis stratified with regard to gender (Table 8), significant results were identified in both male and female populations for the rs11556218 polymorphism in the allelic contrast or the dominant model. However, for rs4778889, allele C was identified to have a slight association compared with allele T in the female population.
In addition, among seven studies that were enrolled in this meta-analysis, six provided ORs adjusted by age and gender between cases and controls through linear regressions. To further eliminate any confounding factors that might have affected our conclusions, adjusted and unadjusted pooled ORs were calculated. Because the data contained in studies are limited, only three genetic models were compared. As shown in Table 9, the overall results of adjusted ORs are consistent with the unadjusted ones. A significant association was found for the rs11556218 polymorphism in the G versus T and TG versus TT models (p<0.05).
Ad, adjusted; Un, unadjusted.
Publication bias
For the comparatively small number of studies enrolled in this meta-analysis, it was difficult to verify the publication bias using the commonly used Begg's test and funnel plots, while Egger's regression was more sensitive in this aspect. Egger's linear regression in this meta-analysis exhibited no publication bias, and p-values of all polymorphisms were greater than 0.1, respectively (rs4778889, t=0.72, p=0.503 for TC vs. TT; rs4072111, t=1.16, p=0.312 for CT vs. CC; rs11556218, t=−0.53, p=0.626 for TG vs. TT).
Discussion
IL-16, a proinflammatory cytokine that has a wide range of biological functions, is also involved in tumor growth and progression (Yellapa et al., 2012). Certain single-nucleotide changes would result in the substitution of amino acid sequelae, thus affecting the expression of the IL-16 protein. Multiple loci of the IL-16 polymorphism have been discovered in relation to susceptibility to cancer. Among them, three polymorphisms located in the promoter or exon regions (nonsynonymous mutation: rs4072111 C/T, Ser/Pro substitution: rs11556218 T/G, Asn/Lys substitution in exon 6 area: rs4778889 T/C in the promoting area) (Nakayama et al., 2000) were the main targets of interest that were explored. The present meta-analysis was based on seven eligible studies quantitatively evaluating the three IL-16 polymorphisms with regard to the potential risk of cancer, including a pool of 1678 cases and 1937 controls, all of Asian origin. Our results from pooled data for the rs11556218 polymorphism indicated that the allele G had a 1.307-fold increased risk factor of systemic cancer when compared with those carrying the allele T. Significant results were also reached under a heterozygote model (OR=1.650; 95% CI, 1.424-1.911) and dominant model (OR=1.605; 95% CI, 1.391-1.845). This phenomenon implies that allele G may carry more pathogenic risks of the occurrence of systemic cancer.
Our results are consistent with previous studies addressing the rs11556218 polymorphism and cancer risk. Gao et al. (2009b), investigating a Chinese cohort stratified by gender with regard to the relationship between IL-16 gene polymorphisms and the risk of development of gastric and colorectal cancers, found that the rs11556218 T/G polymorphism of the IL-16 gene is significantly associated with cancer risk in both male and female populations in allelic contrast. Li et al. (2011), analyzing three polymorphisms of the IL-16 gene in 206 hepatocellular carcinoma cases and 264 healthy controls, found that both the TG genotype and G allele of the rs11556218 T/G polymorphism appear to have a higher risk of hepatocellular carcinoma than wild types. Recent publications also draw similar conclusions in nasopharyngeal carcinoma in a Chinese population and colorectal cancer in an Iranian population (Gao et al., 2009a; Azimzadeh et al., 2011).
Although we found the magnitude of association with cancer risk was relatively low, close attention should be paid to this polymorphism, for it affected a large portion (TG, 42.45%; G, 28.37%) of healthy controls in the pooled study populations. However, no association was observed under a dominant model (GG vs. TT). This conclusion was unexpected. It was assumed that the GG genotype would carry more pathogenic genes for carcinogenesis than would the TG genotype. For this part, we found low expression of the GG genotype (7.14%) in overall study populations, which may explain this discrepancy.
For the rs4778889 and rs4072111 polymorphisms, no associations were identified in the overall population by any of the genetic models. However, for rs4778889, although the association was low, allele C was identified to carry more cancer risk compared with allele T in the female subgroup.
Heterogeneity is observed across the studies of three polymorphisms in Table 4 through Table 6, and thus, we selected random-effect models to combine these pooled ORs. Studies conforming to HWE were also selected and analyzed to eliminate the bias of genetic distributions imposed on the synthesized results. The significance of the overall associations was not statistically altered after eliminating studies that deviated from HWE. To further eliminate selection bias among cases and controls, we compared a pool of adjusted and unadjusted ORs of studies in this meta-analysis. No substantial changes were discovered between the two ORs. This finding indicated that the anthropometric data (age and gender) of cases and controls were well matched in selected studies. Heterogeneity among studies was significantly changed by stratification of cancer types in this study, demonstrating that the type of cancer was a major source of heterogeneity.
Several limitations of our meta-analysis should be noticed when interpreting the results. (1) The number of studies and sample size were relatively small, which may result in reduced statistical power. (2) Genetic distributions may vary according to different ethnicities. The studies enrolled in this study were of Asian origin, mainly accumulating in China and Iran. Information with regard to other ethnicities and populations was insufficient. (3) Because of the lack of data in the original publications, we could not further stratify the results by other potential confounding factors to more fully explore the origin of heterogeneity. (4) The association of gene-gene and gene-environment interactions should be established and analyzed in future studies of the association between IL-16 polymorphisms and cancer risk.
In conclusion, despite these limitations, our meta-analysis provided a more accurate estimate of the association between IL-16 polymorphisms and cancer risk. Our results showed that the rs11556218 T/G polymorphism of the IL-16 gene was significantly associated with an elevated cancer risk in an Asian population. Our results warrant larger, better-designed studies in this field, including a greater variety of ethnicity and cancer type.
Footnotes
Author Disclosure Statement
No competing financial interests exist.