
Abstract
Diseases such as atherosclerosis and coronary artery disease demonstrate disparate population prevalence or present with variable severity in men and women. While the usual explanation points to hormonal status, the role of the Y chromosome has been implicated, but not sufficiently studied. We genotyped six markers of the male-specific region of the Y chromosome, representing the major haplogroups (YAP, G, I, J, K, and R) in 373 male participants of the “Cyprus Study” with ultrasonic data on subclinical atherosclerosis. Of the five major haplogroups identified, two (J and K) accounted for roughly 67% of the Y-chromosome variance among these Greek Cypriot men. Carriers of haplogroup K had a 2.5-fold higher age-adjusted risk for having an atherosclerotic plaque present in any of the four bifurcations scanned, compared to men with other Y-chromosome lineages (OR=2.51; 95% CI=1.18 to 5.33; p=0.017). Carriers of the YAP haplogroup had about 50% less risk for having a plaque in the femoral bifurcation versus the rest (OR=0.46; 95% CI=0.27 to 0.77; p<0.001). We show a possible contribution of the Y chromosome in atherosclerotic phenotypes in men adding to the previous findings for coronary artery disease. Additional studies are warranted as evidence suggests that the Y chromosome could serve as a biomarker for the health status of men.
Introduction
T
The human Y-chromosome phylogenetic tree is now well known, with specific biallelic markers determining the 20 major haplogroups, all descending from a common ancestor (Underhill et al., 2000; Jobling and Tyler-Smith, 2003). A recently published elegant study using this approach has provided significant evidence for the contribution of Y-chromosome genes in increasing the risk for coronary artery disease in males, with carriers of haplogroup I having about a 50% higher age-adjusted risk for coronary artery disease (Charchar et al., 2012). Additionally, older studies had found some evidence that a common biallelic polymorphism of the male-specific part of the Y chromosome was associated with blood pressure (Ellis et al., 2000; Charchar et al., 2002; Shankar et al., 2007; Ely et al., 2010) and with altered cholesterol concentrations (Charchar et al., 2004) of several types.
Prompted by the above findings, we sought to investigate the possible role of the Y chromosome in atherosclerotic disease phenotypes, which are more prevalent in men (Vaccarino et al., 2011), through the analysis of phylogenetic biallelic markers. To this end, we analyzed subjects from the Cyprus Study, a general population, middle-aged prospective cohort on cardiovascular disease and atherosclerosis. Specific aims were to examine the association between markers of the Y chromosome and subclinical atherosclerosis outcomes: (1) Intima-media thickness in the common carotid arteries (IMTcc) and (2) plaque presence (PP) both in the carotid and femoral bifurcations.
Materials and Methods
Study population
Cyprus Study cohort
The Cyprus Study is a population-based cohort study of cardiovascular disease and atherosclerosis in 1106 individuals (48.4% male) aged 40 years or older, from two areas in Cyprus with baseline data collected between 2003 and 2008. A detailed description of the Cyprus Study cohort and ultrasonic analysis has been published previously (Griffin et al., 2009; Panayiotou et al., 2010). Briefly, each carotid bifurcation was examined transversely first, and then longitudinally, to ensure optimal demonstration of the intima-media complex of both the near and far walls of the common carotid artery 1.5-2.0 cm proximal to the carotid bulb. The IMT complex of the far wall of the artery was measured at its thickest part (mean of three readings) and the mean of the measurements from both carotid arteries was used in the analysis (IMTcc). An arterial bifurcation (internal carotid or common femoral bifurcation) was classified as being affected by plaque (PP) if there was a focal thickening of greater than 1.2 mm that did not uniformly involve the whole arterial wall (Ebrahim et al., 1999).
The Ethics Committee of the Cyprus Institute of Neurology and Genetics approved the study and all participants provided written informed consent. For this analysis, baseline data of the first 373 male subjects recruited, who had complete data on IMTcc and PP (outcome), were used.
Genotyping and statistical analysis
The five major clades of the Greek Cypriot Y-chromosome phylogenetic tree (unpublished data of our team) were analyzed by PCR or PCR-RFLP, based on previously published information (Underhill et al., 2000; Consortium The Y Chromosome, 2002; Jobling and Tyler-Smith, 2003). The analyzed genetic biallelic markers, representing those five phylogenetic clades are YAP, I (M258), K (M9), G (M201), and J (M304). R (M207), a sub-haplogroup of K (M9), was also analyzed (Fig. 1).
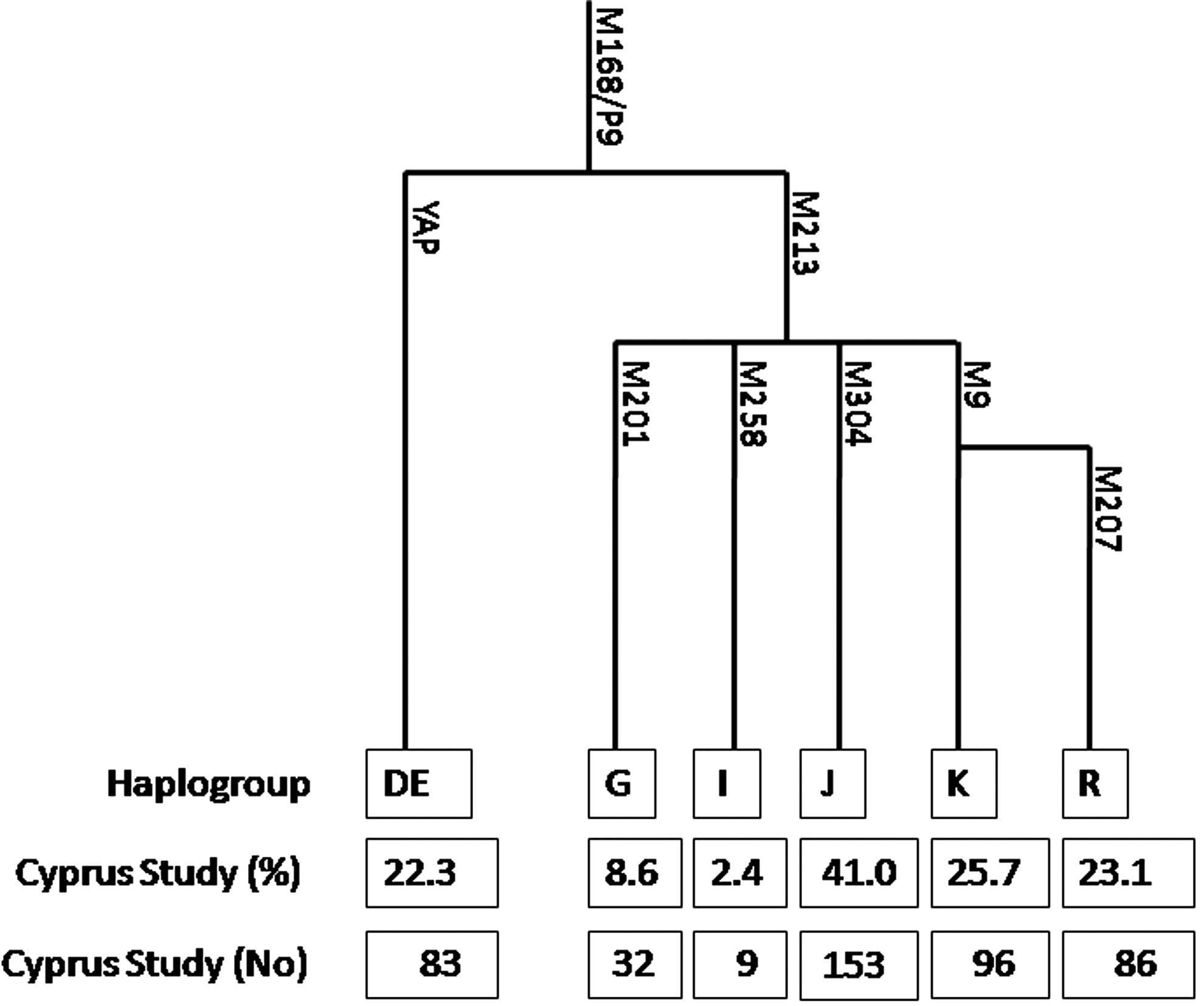
Phylogenetic tree of the Y chromosome with genotyping counts and frequency of haplogroups in the Cyprus Study cohort.
For categorical variables, p-values were calculated using the Pearson chi-square test or Fisher's exact test (two sided) as appropriate and frequencies are presented. Normally distributed continuous variables were compared with the use of t-test and non-normally distributed continuous variables with the Mann-Whitney U test. Additional age-adjusted and fully adjusted (for available demographic and clinical variables that were shown to be associated with both the outcome and the exposures) analyses were done through logistic or linear regression models and adjusted B coefficients or ORs with corresponding 95% CIs are reported. All statistical analyses were performed with STATA v.12 (StataCorp.).
Results
In the Cyprus Study population, the J (M304) haplogroup was the most frequent, found in 41.0% of the subjects, while the K (M9) and YAP accounted for an additional 48% of Y-chromosome variation (25.7% and 22.3%, respectively). The G (M201) haplogroup accounted for 8.6% and I (M258) for only 2.4% of the variation found (Fig. 1). Three individuals were found in the ancestral state for all the five major haplogroups (probably belonging in other rare haplogroups) and were excluded from statistical analysis. Baseline characteristics of included subjects according to atherosclerotic PP are shown in Table 1 and genotypic results according to atherosclerotic PP are shown in Supplementary Table S1 (Supplementary Data are available online at www.liebertpub.com/gtmb). Associations between Y chromosome haplogroups and common risk factors for cardiovascular disease, including high-sensitivity C-reactive protein and fibrinogen, are shown in Supplementary Table S2.
Mean±standard deviation is used for normally distributed values (t-test).
Median and interquartile range is used for non-normally distributed values (Mann-Whitney test).
Chi-square test (two sided) or Fisher's exact chi-square test (where appropriate) is used for categorical variables.
HDL, high-density lipoprotein; hsCRP, high-sensitivity C-reactive protein.
Atherosclerotic phenotypes
No statistically significant association was found between mean IMTcc and any of the five major Y-chromosome markers (Table 2). However, when testing for association with PP in any of the four bifurcations scanned (0 vs. ≤1), an association with the YAP (ORage adjusted=0.55; 95% CI=0.30 to 1.02; p=0.06) and K (M9) (ORage adjusted=2.51; 95% CI=1.18 to 5.33; p=0.017) haplogroups was shown. Further analysis of the K (M9) haplogroup with the R (M207) submarker confirmed the association, showing a similar effect size, with carriers of the R (M207) submarker having more than twofold increased odds for the presence of any atherosclerotic plaques compared with the rest of the subjects (ORage adjusted=2.32; 95% CI=1.06 to 5.09; p=0.036).
IMTcc, intimamedia thickness in the common carotid; PP, plaque presence in any of the four bifurcations scanned (0 vs. 1, 2, 3, 4); PPC, plaque presence in the carotid arteries (0 vs. 1, 2); PPF, plaque presence in the femoral arteries (0 vs. 1, 2).
When further analyzing the PP phenotype according to arterial site, the association between YAP and PP was shown to be driven by the presence of femoral plaques in the femoral arteries, with carriers of the YAP marker having approximately half the odds for PP compared with noncarriers (ORage adjusted=0.46; 95% CI=0.27 to 0.77; p=0.003). Further adjustment for smoking, alcohol, and fibrinogen levels (shown to be associated with the YAP marker here) did not change the association (ORfully adjusted=0.48; 95% CI=0.27 to 0.87; p=0.002) (Table 3). The association between the K (M9) and the R (M207) markers and PP remained for both the carotid and femoral arteries both before and after further adjustment for blood pressure (K: ORcarfully adjusted=2.2; 95% CI=1.23 to 3.91; p=0.008 and ORfemfully adjusted=1.89; 95% CI=1.07 to 3.2; p=0.03, respectively). Results are shown in detail in Tables 2 and 3.
Further adjustment for YAP: smoking (no/ever), alcohol (BU/week), and fibrinogen (μM).
K (M9): systolic and diastolic blood pressure (mmHg).
I (M258): creatinine (ng/mL).
J (M304): diastolic blood pressure (mmHg).
G (M201): no further adjustment.
R (M207): systolic blood pressure (mmHg).
Discussion
To the best of our knowledge, this study is the first to evaluate associations between the main Y-chromosome lineages and atherosclerotic plaques in men. The most important finding from our analysis is that haplogroup K (M9) and its sub-haplogroup R (M207) are associated with a significantly increased risk for atherosclerotic PP in men both in the carotid and femoral bifurcations.
We also show a statistically significant association between carrying the YAP marker and reduced odds for presence of atherosclerotic plaques in the femoral bifurcations, whereas no association was found between any of the Y chromosome markers and IMTcc after adjusting for age. Our findings appear to support the hypothesis set forward by Spence et al., according to which, IMT and atherosclerotic plaque represent different biological and genetically distinct phenotypes and may have different determinants—with IMT being more of an end-organ disease and atherosclerotic plaques relating more to focal disturbances of shear stress (Spence and Hegele, 2004; Spence, 2006)—as well as structural differences between femoral and carotid plaques suggesting a somewhat different pathophysiology (Dalager et al., 2007; Herisson et al., 2011). The association between the K (M9) marker and PP, both in the carotid and femoral bifurcations, was of comparable magnitude when testing for the R (M207) submarker, thus providing additional confidence in the results. Carriers of the K haplogroup had more than 2.5-fold increased odds for having an atherosclerotic plaque compared with the rest and chance was an unlikely explanation for our findings. Although this was a cross-sectional study, an advantage of using genetic markers as exposures is knowledge that the exposure (i.e., genotype assigned at birth) preceded the outcome (atherosclerosis later in life), thus being able to make some causal inferences.
A very recent study showed an association between haplogroup I (M258) and coronary artery disease in two large British cohorts (Charchar et al., 2012). In our study population, haplogroup I was found in only 2.2-2.4% of the subjects (compared with 14.5% to 17% reported in the British populations), whereas the YAP and K haplogroups were found in much higher frequencies than in the BHF-FHS and WOSCOPS populations reported, reflecting the different lineage of the British and Greek Cypriot populations. Although we cannot speculate as to the pathways through which this specific Y-chromosome haplogroup may have an effect on atherosclerosis, it is interesting that inflammatory and immune networks (with inflammation and immunity known to play a great role in atherosclerosis) have been implicated in the possible role of the Y chromosome in coronary artery disease by Charchar et al. (2012). In our study population, systolic blood pressure and alcohol consumption were found to be associated with the K and YAP haplotypes, respectively, as well as with atherosclerotic PP; in addition, smoking and circulating fibrinogen levels were weakly associated with the YAP haplotype. However, further adjustment for those variables did not significantly change these associations (Table 3).
Our findings further strengthen those of Charchar et al. (2012), despite the fact that in this study, we found associations with different haplogroups. This can be explained by the fact that different phenotypes were tested, as well as by the differences in the frequencies of the Y-chromosome markers in the British and Cypriot populations. It is possible that different haplogroups may be of different significance in the Cypriot population, that is, different haplogroups may carry the causative genetic variants. We believe that more studies are warranted for elucidating the contribution of the Y chromosome in diseases presenting with sexual dimorphism. Next generation sequencing approaches can also help in revealing causative variants in populations with different lineages. Chromosome Y may carry valuable genetic markers that can be of tremendous significance in clinical prognosis.
Footnotes
Acknowledgments
The authors express their gratitude to all colleagues who assisted with data collection as well as to all volunteers who participated in the Cyprus Study. The authors also thank Dr. Peter Underhill for useful advice regarding Y-chromosome marker genotyping.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.