
Abstract
Stress has been identified as a common trigger for psychosis. Dopamine pathways are suggested to be affected by chronic and severe stress and to play an important role in psychosis. This pilot study investigates the potential relationship of stress and psychosis in subclinical psychotic experiences. It was hypothesized that single-nucleotide polymorphisms (SNPs) previously found to be associated with psychiatric disorders would be associated with both stress and subclinical psychotic experiences. University students (N=182) were genotyped for 17 SNPs across 11 genes. Higher stress reporting was associated with rs4680 COMT, rs13211507 HLA region, and rs13107325 SLC39A8. Reports of higher subclinical psychotic experiences were associated with DRD2 SNPs rs17601612 and rs658986 and an AKT1 SNP rs2494732. Replication studies are recommended to further pursue this line of research for identification of markers of psychosis for early diagnosis and intervention.
Introduction
P
Stress has been identified as a pivotal marker for onset and more severe presentation of psychosis (Corcoran et al., 2003). Psychological stress is a state in which an individual's coping resources are exceeded resulting in potentially compromised well-being (Lazarus and Folkman, 1984). Heightened vulnerability to stress is associated with the stages of puberty and transition into adulthood because of neurobiological changes and life events (Schulenberg et al., 2004; Spear, 2009; McGorry, 2011).
Stress has also been identified as a marker for the occurrence of subclinical psychosis in a large-scale female twin study (Lataster et al., 2009). Subclinical psychotic experiences are common in the general population and phenotypically and dimensionally similar to severe psychotic experiences (Stefanis et al., 2002; Nuevo et al., 2012). They are suggested to precipitate a severe mental disorder coupled with increased environmental stressors (Cougnard et al., 2007).
Associations between stress and schizophrenia have been investigated using the vulnerability of a stress model (Yuii et al., 2007; Tsoi et al., 2008): Vulnerability is either inborn (genetic predisposition) or acquired (environmental influences) and could potentially trigger psychosis (Zubin and Spring, 1977). Neurobiological findings suggest that early neurobiological or traumatic stressors impact on the hormonal pathways through the stress cascade of the HPA (hypothalamic-pituitary-adrenal) axis, influencing the dopamine pathway and leaving individuals more prone to experiencing stress at a heightened level (Walker and DiForio, 1997; Lee and Schepp, 2009).
A dysfunction in the dopamine pathway has been associated with schizophrenia as dopamine is hypothesized to regulate emotional stability (for a review, Salgado-Pineda, et al., 2005). The dopaminergic activity is influenced by many single-nucleotide polymorphisms (SNPs), for example, within the protein kinase AKT1gene, Catechol-O-methyltransferase gene (COMT), and dopamine D2 receptor gene (DRD2). The COMT involvement in dopamine clearance in the prefrontal cortex, an area associated with cognitive impairment in schizophrenia, suggests a role in the development of schizophrenia and similar disorders (for a review, Williams et al., 2007). COMT SNPs, especially rs4680, have been repeatedly identified as associated with schizophrenia (Egan et al., 2001; Voisey et al., 2010c). DRD2, in particular the rs6277 polymorphism, has been associated with a higher risk of schizophrenia (Lawford et al., 2005; Betcheva et al., 2009; Dubertret et al., 2010; Fan et al., 2010, for a meta-analysis Monakhov et al., 2008), with an increase of receptor availability found in a sample of patients with schizophrenia (Seeman and Kapur, 2000). AKT1 has shown to influence the dopamine pathway (Beaulieu et al., 2009), and gene variants of AKT1 have also been associated with schizophrenia and other mood disorders (Emamian et al., 2004; Karege et al., 2012).
It has been proposed that dysfunction of the glutamatergic system negatively influences the dopamine pathway. Animal models and preclinical studies (Patil et al., 2007) as well as human gene studies (Voisey et al., 2010a) support this notion. For example, the Dystrobrevin binding protein 1 gene (DTNBP1, dysbindin), central in the regulation of the glutamatergic system, was found to be significantly associated with schizophrenia both at the allele and genotype level (SNP rs9370822; Voisey et al., 2010a). The short version of the CAPON gene (carboxyl-terminal PDZ ligand of neuronal nitric oxide synthase) within the glutamatergic system, is associated with both schizophrenia and bipolar disorder (Xu et al., 2005), post-traumatic stress disorder (PTSD), and severe depression in PTSD (Lawford et al., 2013). The HLA (human leukocyte antigen) region is usually associated with immune system function and a genome-wide study found associations of this region in the development of schizophrenia (Schizophrenia Consortium, 2009). A review of published associations of the HLA region with psychiatric disorders suggests that this area is of interest in further elucidating the etiology of psychiatric disorder (Corvin and Morris, 2014). In particular, this review outlined the need for the identification of causative HLA SNPs and their functions to explain the pathogenesis of schizophrenia.
Given their associations with psychosis and their roles in the broader dopamine system, these genes are strong candidates to explore the association between stress, subclinical psychotic experiences, and genetic predisposition, as had previously been proposed (Van Winkel et al., 2008). One study observed shared genetic risk for stress and schizophrenia on the COMT gene (Collip et al., 2011). Voisey et al. (2010b) found shared genetic risk for schizophrenia and PTSD on the DTNBP1 gene. Another study researched shared genetic risk for bipolar disorders and cardiovascular diseases commonly associated with stress and found moderate effect sizes (Ising and Holsboer, 2006). Recently, the Cross-Disorder Group of the Psychiatric Genomics Consortium (2013) found that specific SNPs are associated with a range of psychiatric disorders with both childhood and adulthood onset.
This exploratory study aimed to investigate if genes involved in the wider dopamine system are associated with stress and psychotic experiences in a healthy sample. It was predicted that dopamine system-related SNPs associated with psychotic disorders would be associated with higher reporting of stress and subclinical psychotic experiences.
Materials and Methods
Participants
Participants (N=182) were recruited from the Queensland University of Technology through flyers, lectures, and individual approach on campus. Selection criteria included having sufficient English skills to complete assessments, being between 17 and 25 years of age, no reported history of brain injury, and Caucasian ethnicity. The mean age of this sample was 20.48 years (SD=2.48). The majority of the participants were females (70.3%).
Participants received information about the nature of the study and data collection. Written consent was obtained before data collection began. Ethics approval for the project was obtained from the Human Research Ethics Committee of the Queensland University of Technology.
The participant questionnaire contained demographic information, measures for stress reporting, and subclinical psychotic experiences.
The Chronic Stress Scale (CSS) measures ongoing psychological stress over the past 6 months in seven categories: marital, parental, filial, financial, occupational, ecological, and physical (Norris and Uhl, 1993). The total scale has good internal reliability with an overall Cronbach's alpha of .83 (Norris and Uhl, 1993). For the current study, five subscales were chosen, modified, and renamed as per White et al. (2011), to capture the target group of this study: marital stress changed to relationships, filial stress to family, financial stress to finances, occupational stress changed to work and study stress and, without modification, physical stress. The 18 items are rated on a five-point Likert scale from 0=never to 4=very often.
The Community Assessment of Psychic Experiences (CAPE) scale consists of two parts measuring subclinical psychotic experiences and their impact (qualifying subscale) in the general population (Stefanis et al., 2002). It has sound test-retest reliability and discriminant validity (Konings et al., 2006). The scale has 42 items rated on a four-point Likert scale: 0=never to 3=nearly always. The qualifying subscale was not used for analysis.
SNP selection and genotyping
DNA was obtained by collection of a saliva sample using Oragene kits (DNA Genotek, Inc.). A total of 18 SNPs were selected from 11 genes previously associated with schizophrenia. A list of the SNPs and the associated gene is displayed in Table 1.
SNP, single-nucleotide polymorphism.
Samples were genotyped using a homogeneous MassEXTEND (hME) Sequenom assay performed by the Australian Genome Research Facility. The hME assay is based on the annealing of an oligonucleotide primer (hME primer) adjacent to the SNP of interest. The addition of a DNA polymerase along with a mixture of terminator nucleotides allows extension of the hME primer through the polymorphic site and generates allele-specific extension products, each having a unique molecular mass. The resultant masses of the extension products are then analyzed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) and a genotype is assigned in real time. Six SNPs had fails: rs4680 (1.65%), rs165774 (1%), rs6277 (0.55%), rs6589386 (6%), rs3803300 (2.2%), and rs1315259 (0.55%). All other SNPs had a pass rate of 100%.
Statistical analyses
A priori power calculations for one-way analyses of variance (ANOVAs) of three groups showed that a sample size of 159 was necessary for an acceptable power of .80 and a small effect size of 0.2 per SNP.
All analyses were performed using SPSS Statistic Package 18 for MacIntosh Computers and SPSS 19 for PCs (SPSS, 2009). Calculations for the Hardy-Weinberg equilibrium were performed using the Utility Programs for Analysis of Genetic Linkage (Ott, 1988). Odds ratios were calculated using the COMPARE2 program from the WinPepi suite of epidemiology programs (Abramson, 2004). All data were screened for entry errors, missing values, and breaches of assumptions. Little's Missing Completely At Random test was nonsignificant (χ2=0 1465.83, p=0.47) indicating that data were missing at random. Missing data were excluded list-wise. An alpha level of 0.05 was applied to determine significance unless otherwise specified.
Descriptive analysis for the CSS score distribution revealed a mean of 19.02 (SD=8.62). Normality for the CSS distribution was accepted upon visual inspection. Internal reliability for the CSS scale was acceptable (Cronbach's α=0.74).
The CAPE distribution was bimodal and a split at a median of 47 was performed to create two groups for further analysis. While this approach can reduce statistical power, other modes of analysis yielded unsatisfactory model fits. Reliability of the CAPE frequency scale was very high (Cronbach's α=0.98), as expected with the large number of items (Cortina, 1993). Moderate to high correlations between the subscales of the CAPE have previously been reported. Subclinical psychotic experiences in the general population are probably more attenuated than in clinical samples (Stefanis et al., 2002).
Results
Chronic stress scale
ANOVAs were performed individually for the effects of 18 SNPs on CSS scores. Three SNPs, rs4680 (COMT), rs13211507 (HLA region), and rs13107325 (SLC39A8), were associated with higher stress scores. Results of the ANOVAs for all 18 SNPs are shown in Table 2.
SNPs in bold reached significance.
N=182.
df, degrees of freedom.
Post hoc tests with Bonferroni and Tukey adjustment were performed for the significant SNPs. For the rs13107325 SNP, participants with genotype CC (n=144) had a significantly higher mean score on the CSS (M=19.84, SD=8.64) than those with genotype CT (n=30; M=15.23, SD=7.66; Tukey p=0.02, 95% CI [−0.57, 8.64]; Bonferroni p=0.02, 95% CI [0.48, 8.73]). There was no significant difference in means between genotypes CC and TT or CT and TT (n=2). Findings for genotype TT need to be interpreted with caution as there were only two cases. Figure 1 shows the CSS mean scores per genotype.
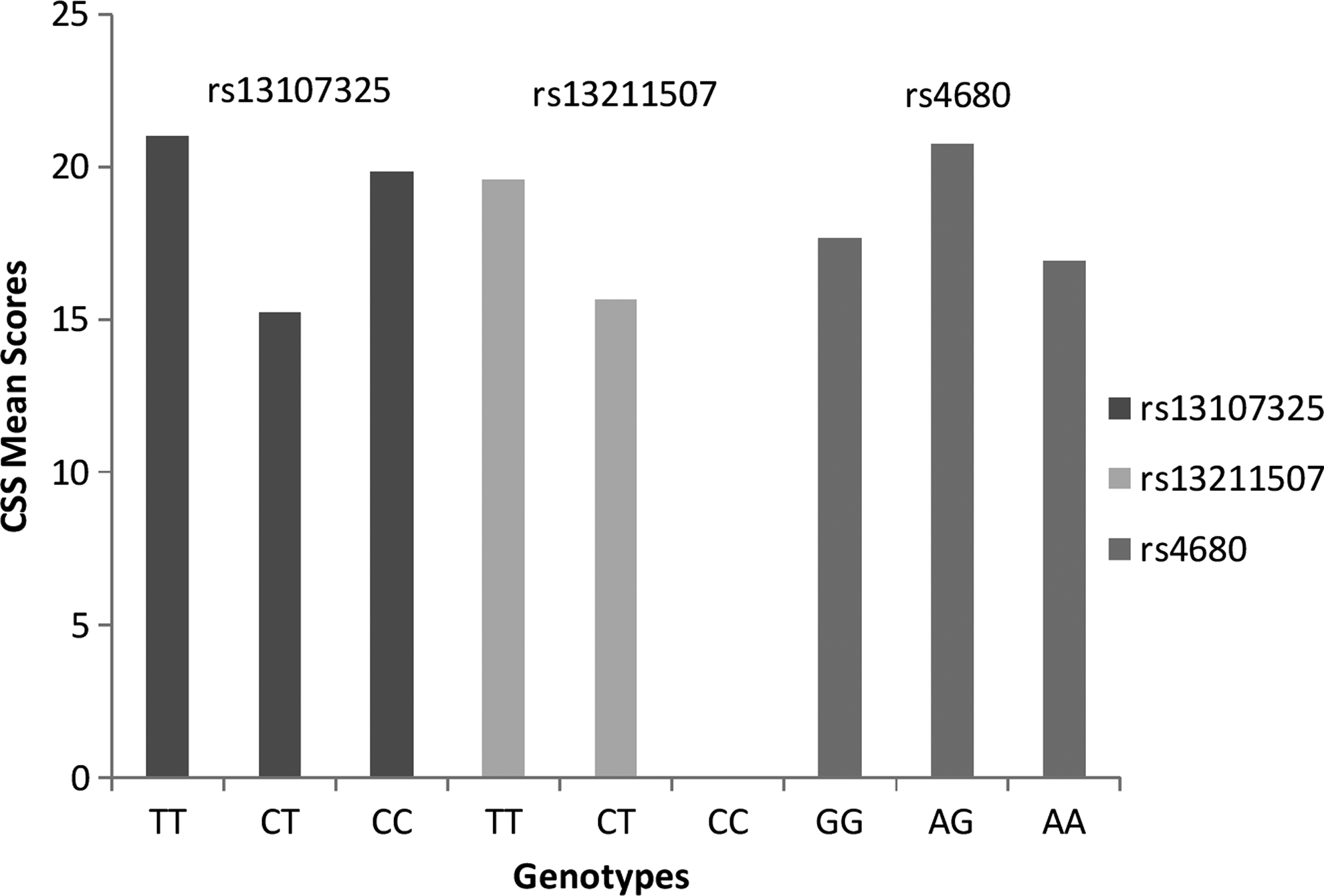
Associations of genotype and chronic stress (Chronic Stress Scale Scores) for SNPs rs13107325 SLC39A8, rs13211507 HLA region, and rs4680, COMT.
For rs13211507, participants with genotype TT (n=153; M=19.56, SD=8.73) had significantly higher CSS mean scores than genotype CT (n=25; M=15.68, SD=7.15). Figure 1 shows the CSS mean scores per genotype. No CC genotypes were reported.
For rs4680, participants with genotype AG (n=93) had significantly higher means (M=20.76, SD=8.98) than those with genotype AA (n=46, M=16.93, SD=7.24; Tukey p=0.03, 95% CI [0.22, 7.44]; Bonferroni p=0.04, 95% CI [0.14, 7.51]). There was no significant difference between genotypes AA and GG (n=36) or AG and GG (Fig. 1).
Genotype frequencies indicated that all three significant SNPs were in the Hardy-Weinberg equilibrium. None of the three SNPs survived correction for multiple testing with the Benjamini-Hochberg model (Benjamini and Hochberg, 1995).
CAPE scores
Chi-square analysis was performed for each of the 18 SNPs by the CAPE score group. Three SNPs, rs17601612 DRD2, rs2494732 AKT1, and rs6589386 DRD2, were significantly associated (p=0.009, p=0.02, p=0.03, respectively) with higher levels of psychotic experiences. Chi-square results for all 18 SNPs are shown in Table 3.
SNPs in bold reached significance.
CAPE, community assessment of psychic experiences.
Odds ratios were calculated yielding high ratios. Tables 4-6 show genotype frequencies of rs17601612, rs2494732, and rs6589386, respectively, for participants reporting higher levels compared with those reporting lower levels of psychotic experiences. For rs17601612, participants with genotype CC were five times more likely to report psychotic experiences than those with genotypes GG or CG. Odds ratios between CG and GG were not significantly different (Table 4).
For rs2494732, participants with genotype CC were four times more likely to report psychotic experiences than those with genotypes CT and TT. Odds ratios for CT and TT were not significantly different (Table 5).
For rs6589386, participants with genotype CC were three times more likely to report psychotic experiences than those with genotypes CT and TT (approaching significance, p=0.05). Participants with genotype CT were three times more likely to report psychotic experiences than participants with genotype TT (Table 6).
All three SNPs were in the Hardy-Weinberg equilibrium. After multiple testing using the Benjamini-Hochberg methodology (Benjamini and Hochberg, 1995), none of the SNPs maintained significance.
Discussion
The aim of this pilot study was to investigate the relationship between stress and subclinical psychotic experiences and genetic predisposition in a healthy sample of young adults. Stress has been identified as a strong marker of onset and maintenance of clinical psychosis (Corcoran et al., 2003). Physiologically, the dopamine system has been implied to play a role in psychosis (Salgado-Pineda et al., 2005) and to be affected by stress (Walker and DiForio, 1997; Lee and Schepp, 2009). This led to the assumption of shared genetics between psychosis and stress (Van Winkel et al., 2008).
Subclinical psychotic experiences have been found with high prevalence in the general population (Nuevo et al., 2012) with strong similarities to clinical psychosis (Stefanis et al., 2002). They are suggested to be a potential early marker for the development of clinical psychosis (Cougnard et al., 2007; Mossaheb et al., 2012). This study found partial support for these assumptions in that SNPs previously associated with schizophrenia were found to play a role both in chronic stress and subclinical psychotic experiences.
Our hypothesis focused on findings from genetic research that a variety of genes and corresponding SNPs influence the broader dopamine system, potentially increasing the risk for psychotic disorders. Shared genetics of the dopamine system between patients and healthy controls might also influence stress responses (Van Winkel et al., 2008). Our results suggest that both assumptions are worthy of further investigation. Three SNPs, rs13107325, rs13211507, and rs4680, were found to be significantly associated with CSS scores. SNPs rs17601612, rs2494732, and rs6589386 yielded significant associations with high ratings on the CAPE scale. However, none of the SNPs withstood corrections for multiple testing, likely due to the large number of SNPs analyzed in this study and the moderate sample size. An association of genes potentially involved in psychotic disorders and stress had been proposed previously (Van Winkel et al., 2008). The nonsynonymous SNP rs4680 of the COMT gene has been associated with stress (Collip et al., 2011) indicating that stress sensitivity may be a useful marker of liability to psychosis.
Involvement of rs13211507 located in the HLA region with chronic stress suggests an association with the immune system function that requires further investigation. Previous research has identified associations with intense stress resulting from a traumatic event and the immune system (Andrews and Neises, 2012; Jones and Thomsen, 2013 for reviews). This SNP and other SNPs located in chromosome 6 are associated with schizophrenia (Stefansson et al., 2009; for a review, Ozomaro et al., 2013).
We also identified association of rs13107325 located in the SLC39A8 gene with chronic stress. The SLC39A8 gene is responsible for transport of important metals that play a role in brain development and function (He et al., 2006). Deletion of SLC39A8 has been associated with mental retardation, and other SLC39 regions might be involved in psychiatric disorders such as the bipolar disorder (Ollila et al., 2009) and major depressive disorder (Muglia et al., 2010). More research is needed to understand the association of the SLC39A8 SNP rs13107325 (Carrera et al., 2012).
Genetic associations with CAPE scores showed significance for two DRD2 SNPs and one AKT1 SNP. Both genes influence the dopamine system and might be important for emotion regulation (Salgado-Pineda et al., 2005). Both genes have been extensively studied for their role in psychotic disorders (Egan et al., 2001 for AKT1; Lawford et al., 2005 for DRD2). In a cohort of participants with a lifetime diagnosis of psychosis, AKT1 SNP rs2494732 showed strong association with psychosis rather than bipolar disorder (Karege et al., 2012). The association of these genes with subclinical psychotic experiences suggests that genetic predisposition heightens the risk for people with specific genotypes (Stefanis et al., 2002). The high odds ratios reported suggest genetic predisposition to be an important factor in developing subclinical psychotic experiences.
These findings should be interpreted with caution. This pilot study was based on a convenience sample with a strong bias toward female participation with modest participant numbers. Additional data collection will be needed to obtain greater statistical power and robustness. Because of the large number of SNPs tested and the modest sample size, none of these findings withstood multiple testing. The study did not investigate potential mediator and moderator variables, such as gender and substance use, due to the modest sample size and limited power. Investigating potential interactions could be important to better understand the data.
In general, findings of this pilot study form an encouraging basis for further research into the relationship of stress, genetic predisposition, and subclinical psychotic experiences. Genetic predisposition led to increased experience of subclinical psychotic experiences and higher stress ratings. If shared genetic risk between psychotic disorders, subclinical psychotic experiences, and stress is confirmed, high-risk individuals with a family history of psychotic disorders and patients suffering from stress-influenced disorders may particularly benefit through early diagnosis (Gask et al., 2008; Mossaheb et al., 2012), prevention, and early stress management (Betensky et al., 2008).
Footnotes
Acknowledgments
This work was financially supported by the Queensland State Government, the Nicol Foundation, and the Institute of Health and Biomedical Innovation, QUT. JV is a Queensland Government Smart State Fellow.
Author Disclosure Statement
All authors declare that they have no conflicts of interest.