
Abstract
Objectives: Tetralogy of Fallot (TOF) is one of the most common forms of congenital heart disease. In this study, we aimed at investigating the associations between genetic variations of vascular endothelial growth factor (VEGF) gene and the risk of TOF in a Chinese Han population. Our findings may contribute to a deeper understanding of TOF pathogenesis and better diagnostic and therapeutic suggestions. Methods: A total of 165 TOF patients and 240 controls from a Chinese Han population in Shenyang and Harbin were recruited in the current study. Nine single-nucleotide polymorphisms (SNPs) (−2578C/A, −460T/C, −1154G/A, −634G/C, 534C/T, +398G/A, +963C/T, 752C/T, 913G/A) were genotyped by the MALDI-TOF MassARRAY system. Individual SNPs as well as their haplotypes were analyzed for their associations with TOF risk, using odds ratios and the 95% confidence interval under codominant and dominant models. Results: In the single SNP analyses, the mutant homozygous genotypes of −2578C/A (rs699947) and +963C/T (rs3025039) were related with an increased risk of TOF. In addition, carriers with the mutant A allele of −1154G/A (rs1570360) were supposed to have a significantly elevated TOF risk. Similarly, compared with the wild homozygote GG carriers, the GC carrier of −634G/C (rs2010963) revealed a significant relationship with susceptibility of TOF, but not for the mutant homozygote CC carriers. However, no significant association was found for the other five SNPs. Meanwhile, haplotype analysis revealed that CCA and ATA in block 1 (−2578C/A, −460T/C, and −1154G/A) and TTG and TCA in block 3 (+963C/T, 752C/T, and 913G/A) were significantly related with an increased TOF risk compared with the most common haplotypes. Conclusion: In summary, our results suggested that VEGF variants (−2578C/A, −1154G/A, −634G/C, +963G/A) were involved in the susceptibility of TOF. However, validation of our study needs further study in various ethnics to reveal the functional relationship between VEGF polymorphisms and TOF risk, which may contribute to diagnosis and therapy of TOF.
Introduction
C
The VEGF gene, which is located on chromosome 6p12, plays an important role in angiogenesis (Brogan et al., 1999). Among the common angiogenic factors, VEGF is the only mitogen that acts on endothelial cells. It promotes the elongation, network formation, and branching of endothelial cells (Helmlinger et al., 2000), and then, endothelial cell proliferation, migration, and survival contribute to angiogenesis. When the heart is still avascular, VEGE facilitates the proper heart morphogenesis (Dor et al., 2001). Several previous studies reported that genetic variations in the VEGF gene significantly associated with several human diseases, including CHD (Wang et al., 2014), coronary artery disease (CAD) (Gu et al., 2013), congenital amyotrophic lateral sclerosis (Terry et al., 2004), age-related macular degeneration (Richardson et al., 2007), and rheumatoid arthritis (RA) (Zhang et al., 2013). Considering the critical role of VEGF in angiogenesis, it is speculated that the common single-nucleotide polymorphisms (SNPs) of VEGF gene may be related to TOF susceptibility. In a family-based study in the United States, three VEGF polymorphisms, that is, rs699947, rs1570360, and rs2010963, were associated with the pathogenesis of TOF, which indicated that low expression of VEGF increased the risk of TOF (Lambrechts et al., 2005). However, to date, no previous research concerning the relationship between SNPs within the VEGF gene and TOF risk has been conducted in an Asian population. To verify our hypothesis, we conducted a case-control study in a Chinese Han population. Our findings may contribute to a deeper understanding of TOF pathogenesis and better diagnostic and therapeutic suggestions.
Materials and Methods
Characteristics and source of participants
A total of 165 TOF cases and 240 healthy controls were recruited in the current study. Patients newly diagnosed with histopathologically confirmed TOF were consecutively recruited from the Department of Pediatrics at Shengjing Hospital of China Medical University and the Department of Cardiology at the Fourth Hospital of Harbin Medical University between 2013 and 2014. Patients with a history of cancer and previous radiotherapy and chemotherapy were excluded. In addition, diagnoses were validated by trained abstractors and electronic medical and pathological records of all TOF patients were analyzed to confirm that there was no undiagnosed mystic tumor when recruited. The healthy controls were patients without TOF, matched for age and gender, and recruited from the same hospitals during the same period. Most of the controls were admitted to hospital for the treatment of common cold. All the cases and controls were Chinese from Shenyang and Harbin without a genetic relationship. Perioperative characteristics and complications of TOF patients were also collected. All the clinical characteristics of participants are shown in Table 1. Approval of the study protocol was obtained from the Ethics Committee of the Shengjing Hospital of China Medical University and the Fourth Hospital of Harbin Medical University. All participants (represented by their mothers) provided written informed consent to participate in the study.
ASD, atrial septal defect; CPB, cardiopulmonary bypass; ICU, intensive care unit; PDA, patent ductus arteriosus; SD, standard deviation; TOF, Tetralogy of Fallot; VSD, ventricular septal defect.
Tag SNP selection
Tag SNPs in the VEGF gene were identified using genotype data of Han Chinese in Beijing (CHB) from the International HapMap Project (HapMap Data Rel 24/phaseII Nov08, on NCBI B36 assembly, dbSNP b126). A total of 18,207 bp VEGF gene region was covered, encompassing the whole VEGF gene region from the 5′ untranslated regions (5′ UTR) and 3′ UTR, as shown in Figure 1. Retrieved data were analyzed with HaploView 4.2 software, with filters being strong linkage disequilibrium (LD) tagging criteria of r2>0.8, minor allele frequency (MAF) >5%, and D′=1. Finally, nine SNPs were derived according to the selection principle. These nine SNPs are shown as Figure 1, including −2578C/A (rs699947), −460T/C (rs833061), −1154G/A (rs1570360), −634G/C (rs2010963), 534C/T (rs25648), +398G/A (rs833068), +963C/T (rs3025039), 752C/T (rs3020040), and 913G/A (rs10434). The selected SNPs consisted of three SNPs in the promoter region (−2578C/A, −460T/C, −1154G/A), one SNP in 5′ UTR (−634G/C), one SNP in extron1 (534C/T), 1 intronic SNP (+398G/A), and three SNPs in 3′ UTR (+963C/T, 752C/T, 913G/A).
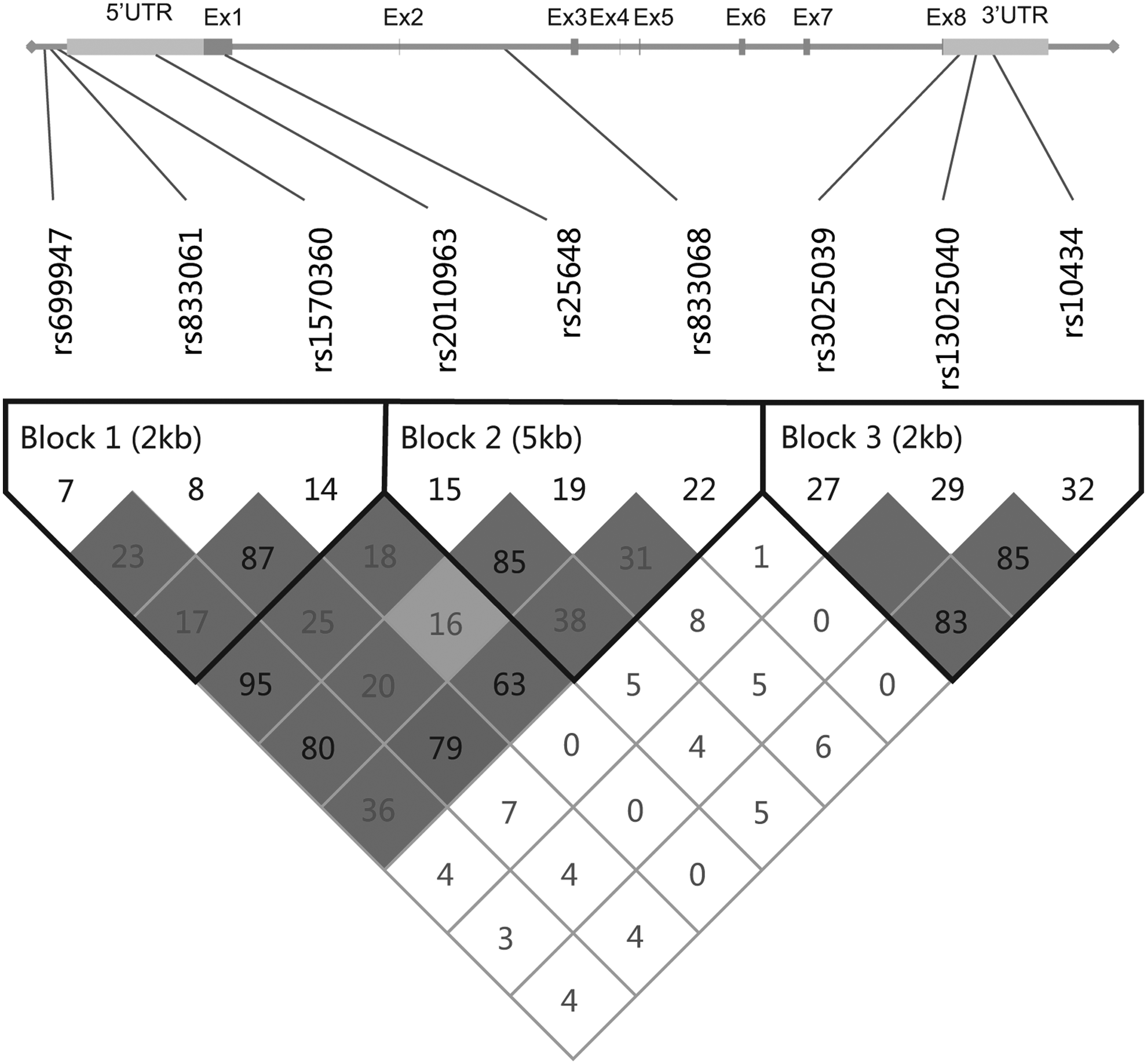
Graphical presentation of the selected nine single-nucleotide polymorphisms (SNPs) of VEGF and linkage disequilibrium (LD) plot of VEGF. Three haplotype blocks were defined using HaploView 4.2 with the setting of confidence intervals. VEGF, vascular endothelial growth factor.
DNA extraction and genotyping
Ten microliters of whole blood sample was collected from each subject using vacutainer tubes and then transferred to test tubes with ethylenediaminetetraacetic acid (EDTA). These samples were stored in −4°C refrigerator for further study. The QIAamp DNA Blood Mini Kit (QIAGEN) was applied for DNA extraction. Primers of selected SNPs were designed by MassARRAY Assay Designer 3.1 (Sequenom, Inc.) for polymerase chain reaction (PCR) amplification. In brief, the reaction was performed in 50 μL of reaction volume containing 3 μL extracted DNA, 1.5 mM MgCl2, 1 μL of 15 mM DNTP, 5 μL PCR buffer (10×), and 2 U Taq DNA polymerase. The amplification started with 94°C for 5 min, followed by 40 cycles of 94°C for 20 s, 56°C for 30 s, 72°C for 45 s, and a final polishing step of 72°C for 6 min. After amplification, the unincorporated PCR products need to be purified. We performed it using Shrimp Alkaline Phosphatase (SAP) Methods. 0.2 U of SAP and 0.2 U Exonuclease III were put in the PCR product purification system, at a condition of 37°C for 22 min followed by 75°C for 6 min. Finally, the extension products were put in a reaction mixture containing 0.4 μL SAP, purified in 37°C for 50 min, and then terminated in 75°C for 10 min. Genotyping was conducted by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using the MassARRAY method.
Statistical analyses
Deviation from the Hardy-Weinberg equilibrium (HWE) was assessed by Pearson's χ2 test among healthy controls for all SNPs. Logistic regression was conducted to estimate odds ratios (ORs) and their 95% confidence intervals (CIs) of SNPs adjusted for age and gender. A two-sided p-value less than 0.05 was considered to be statistically significant. MAF of the reference group (shown in Table 2) was the ancestral allele of these nine SNPs according to the MAF in Han Chinese in Beijing from HapMap database. Codominant and dominant genetic models were used to evaluate the associations between tag SNPs and TOF susceptibility. The degree of LD among markers, which was used to divide haplotype blocks, was analyzed by the 4-gamete rule of HaploView 4.2. No recombination was discovered and the LD blocks were established only if three haplotypes were found with a frequency more than 0.01 when compared with consecutive markers. Adjusted ORs and 95% CIs were also used to analyze the haplotypes compared with the common haplotype.
Values in bold indicate that p<0.05.
MAF in Han Chinese in Beijing from HapMap database.
MAF in controls of the current study.
CI, confidence interval; HWP, p-value of Hardy-Weinberg equilibrium; MAF, minor allele frequency; OR, odds ratio adjusted for age and gender; SNP, single-nucleotide polymorphism; VEGF, vascular endothelial growth factor.
Results
Characteristics of the study population
The characteristics and clinical data of cases and controls are summarized in Table 1. The TOF cases and healthy controls appeared to be matched adequately on age and gender, as suggested by χ2 tests (p=0.896 and 0.473, respectively). The mean age of the subjects was 1.55 years for TOF patients and 1.57 for healthy controls. About 65.5% and 61.7% of patients and controls were male. TOF patients could be categorized into four groups according to the type of complications: isolated TOF, TOF with ASD (atrial septal defect), TOF with PDA (patent ductus arteriosus), and TOF with ASD and PDA. Of all the patients, isolated TOF patients accounted for 71.5%, 17.6% patients were TOF with ASD, 7.9% were TOF with PDA, and others were TOF with ASD and PDA (3.0%). Perioperative characteristics of TOF patients were also recorded as following: the mean cardiopulmonary bypass time was 2.16±1.27 h, median ventilation time was 22±5.66 h, and median length of intensive care unit stay was 49±7.17 h.
Analysis of independent effect of single SNP
Nine distinct SNPs, that is, −2578C/A, −460T/C, −1154G/A, −634G/C, 534C/T, +398G/A, +963C/T, 752C/T, and 913G/A, were retrieved from the HapMap databases. The identification number and relative position of the tag-SNPs are shown in Figure 1. Table 2 shows the information of the allelic frequencies of the tag-SNPs. All the SNP frequencies of the controls complied with the HWE. The MAF in controls of the current study was in the range of the reference for the Han Chinese population (as shown in Table 2). Distributions of the minor allele of −2578C/A and −1154G/A, situated in the promoter region, were found to be significantly different between the controls and the cases, with OR of 1.42 (95% CI=1.05-1.92, p=0.024) and OR of 1.74 (95% CI=1.28-2.37, p<0.001), respectively. The minor C allele of −634G/C could elevate the risk of TOF significantly with OR of 1.36 (95% CI=1.02-1.82, p=0.035). Meanwhile, the frequency of +963T was higher in the group of TOF patients when compared with the healthy controls (OR=1.43, 95% CI=1.01-2.01, p=0.042). However, there were no differences of allelic distribution frequencies for the other five SNPs between cases and controls.
Logistic regression analyses of associations between VEGF SNP genotypes and TOF risk are presented in Table 3. Four SNPs in the codominant model (−2578C/A, −1154G/A, −634G/C, +963C/T) and two SNPs (−1154G/A, −634G/C) in the dominant model revealed significant differences compared with the healthy controls. Compared with the wild homozygote genotype CC, carriers with the mutant AA genotype of −2578C/A showed an increased risk of TOF (OR=2.39, 95% CI=1.13-5.06, p=0.020). Mutation of −1154G/A could elevate the risk of TOF in the codominant model (GA vs. GG: OR=2.15, 95% CI=1.41-3.28, p<0.001; AA vs. GG: OR=2.29, 95% CI=1.09-4.80, p=0.026) and in the dominant model (GA+GG vs. AA: OR=2.17, 95% CI=1.45-3.26, p<0.001). The GC carrier of −634G/C (rs2010963) revealed a significant relationship with susceptibility of TOF (GC vs. GG: OR=1.83, 95% CI=1.18-2.84, p=0.007), but not for the mutant homozygote CC carriers, when compared with the wild homozygote GG carriers. In addition, the risk effect for related genotypes of +963C/T was also observed under the codominant model (TT vs. CC: OR=3.26, 95% CI=1.18-8.96, p=0.017). However, no genotype frequencies of the other five SNPs altered significantly compared with the controls as the allelic distribution frequencies.
Values in bold indicate that p<0.05.
Haplotype analysis for the association between VEGF SNPs and TOF risk
LD analysis between the cases and controls revealed that the nine SNPs were split into three different haplotype blocks. As shown in Table 4, block 1 was situated in the region of promoter with a length of 2 kb, containing three SNPs (1-3: −2578C/A, −460T/C, −1154G/A). Block 2 covered a section of 5′ UTR (4: −634G/C) and the coding regions of exon1 (5: 534C/T) and intron2 (6: +398G/A) and encompassed 5 kb. Block 3 included the ending part, located in the region of 3′ UTR, and three SNPs (7-9: +963C/T, 752C/T, 913G/A) belong to this block. The SNPs related with TOF risk reside in block 1 (−460T/C, −1154G/A) and block 3 (752C/T, 913G/A). Haplotype-specific analysis showed significant frequency differences on the haplotype CCA and haplotype ATA in block 1, with p of 0.038 and 0.001 respectively, when compared with the common haplotype CTG. The haplotypes CCA and ATA were associated with an increased TOF risk (OR=1.75, 95% CI=1.03-2.97, p=0.038; OR=5.90, 95% CI=1.82-19.08, p=0.001, respectively). Similarly, due to higher frequencies in TOF patients, a risk effect was found in haplotype TTG and TCA in block 3 (OR=3.50, 95% CI=1.06-11.62, p=0.030; OR=4.05, 95% CI=1.05-15.57, p=0.028, respectively). However, no association was found in block 2.
Values in bold indicate that p<0.05.
Order of polymorphisms is −2578C/A (rs699947), −460T/C (rs833061), −1154G/A (rs1570360) in block1, −634G/C (rs2010963), 534C/T (rs25648), +398G/A (rs833068) in block2, +963C/T (rs3025039), 752C/T (rs3025040), and 913G/A (rs10434) in block 3.
All other haplotypes that had a frequency <1% in either cases or controls are not listed.
Adjusted for age and sex.
LD, linkage disequilibrium.
Discussion
In the current study, we aimed at investigating the associations between SNPs in the VEGF gene and TOF susceptibility in the Han Chinese population. A case-control study of 165 TOF patients and 240 controls was performed. In the present study, all the nine selected SNPs, including two SNPs (−2578C/A, −1154G/A) located in the promoter region and two SNPs (−634G/C and +963C/T) located in the region of 5′ UTR and 3′ UTR, respectively, showed significant associations with the risk of TOF. Meanwhile, haplotype-specific analysis revealed that CCA and ATA in block 1 (−2578C/A, −460T/C, and −1154G/A) and TTG and TCA in block 3 (+963C/T, 752C/T, and 913G/A) were significantly related with TOF risk compared with the most common haplotypes.
Angiogenesis is a prerequisite for adequate oxygen and metabolite supply to tissues. Therefore, it plays an essential role in tumor growth and progression. Among various enzymes involved in angiogenesis, VEGF acts as the most specific and crucial stimulus to induce migration, proliferation, and survival of endothelial cells. A large body of previous literature incriminates VEGFA as the most potential mediator of angiogenesis in TOF. Abnormal angiogenesis is a major process leading to TOF, while a complex multistep process involving endothelial cell activation, migration, proliferation, tube formation, and maturation contributes to angiogenesis. It is speculated that the binding of VEGF and VEGFR2 triggered several downstream pathways, such as the Ras-MAPK pathway (Kim et al., 2002), DAG-PKC pathway (Verrier et al., 2004), NOS pathway (Lu et al., 2011), FAK/Paxillin pathway (Schaller, 2004), P38 pathway (Wu et al., 2006), and AKT pathway (Li et al., 2014), which are involved in cell proliferation, migration, survival, and also contributed to angiogenesis, which eventually play a vital role in TOF, as shown in Figure 2. Another downstream signal pathway, the VEGF-NOTCH pathway, also plays an important role in angiogenesis; yet, no previous study had indicated the direct effect on TOF risk (van den Akker et al., 2012). However, the potential mechanism of the above signal pathways, shown in Figure 2, needs further validations. Interestingly, only an exceptional prior study indicated that the VEGF genetic variation does not significantly contribute to congenital cardiovascular malformation susceptibility (Griffin et al., 2009). However, emerging studies tend to believe that VEGF polymorphisms are associated with CHD formation or TOF risk (Lee and Hyun, 2009; Wessels and Willems, 2010; Silversides et al., 2012).
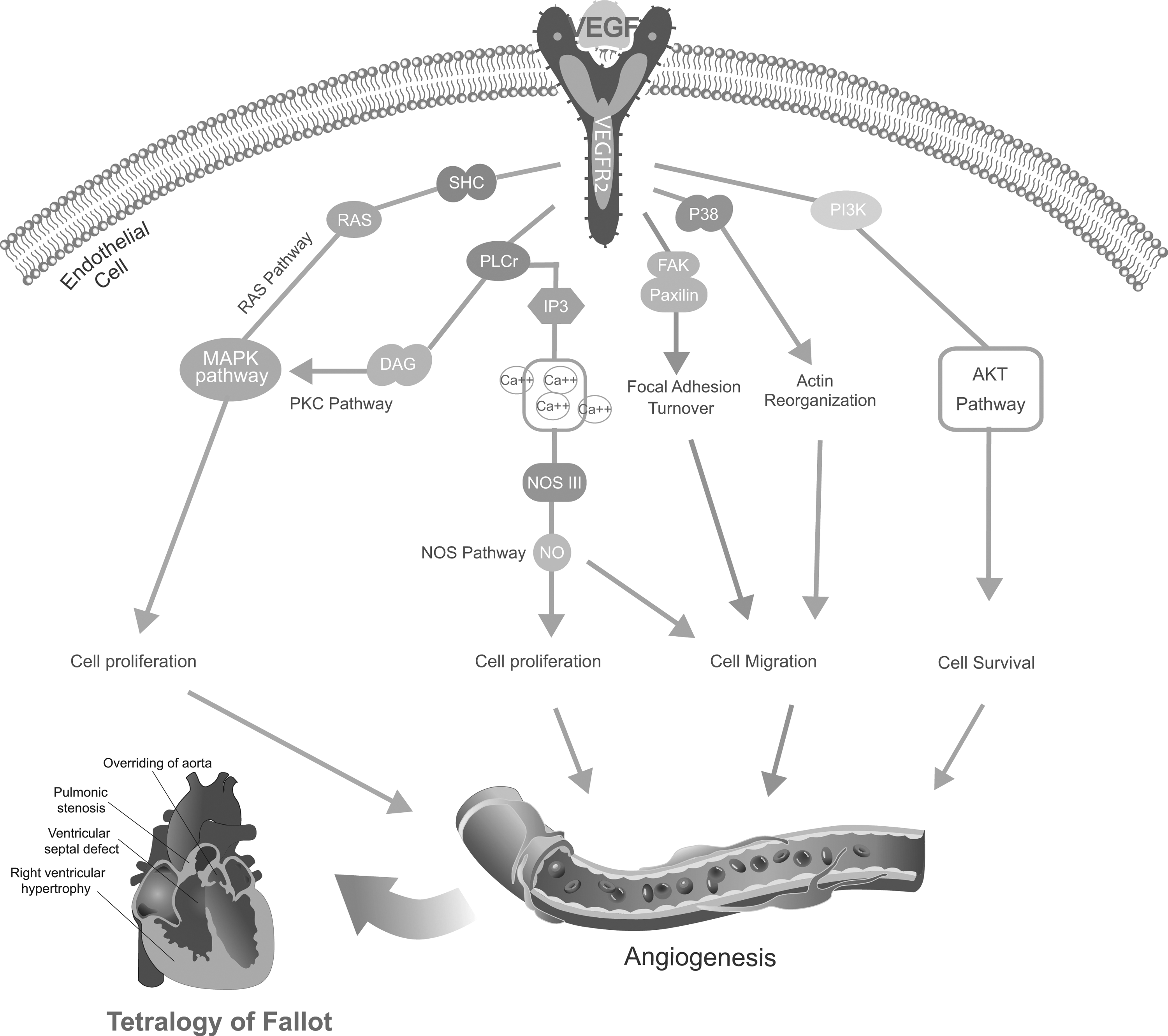
Potential schematic overview of the interactions between angiogenesis and Tetralogy of Fallot (TOF) risk. RAS-MAPK pathway, DAG pathway, and NOS pathway lead to cell proliferation. The NOS pathway also contributes to cell migration as well as FAK/Paxillin pathway and P38 pathway. Meanwhile, AKT pathway contributes to angiogenesis through PI3K.
As for the genetic variation of VEGF, in our study, the four eligible SNPs in VEGF, −2578C/A and −1154G/A are located in the promoter, −634G/C is situated in the region of 5′UTR, while +963G/A is located on 3′UTR, which means that these gene segments will not be transcribed in gene expression. However, functions of the untranslated regions of eukaryotic mRNAs are complicated. It is indispensable to correct transcription. The region of promoter initiates the regulation of transcription. 5′UTR is mainly responsible for transcription, which affects the transcription of each stage, including the stability of mRNA, folding, and interaction of the ribosome (Anthonisen et al., 2001). The functions of 3′UTR include regulating mRNA stability and degradation rate, controlling the efficient use of DNA templates, and helping to identify the special codon. At the same time, 3′UTR also regulates the transcription time, site, and products of specific mRNA (Jupe et al., 1996; de Moor et al., 2005; Shin et al., 2007). Therefore, genetic variants in these regions may lead to overexpression or low expression of VEGF. Researchers found that variations of VEGF had association with many diseases, such as breast cancer, CAD, and RA (Cui et al., 2013; Zhang et al., 2013; Rahoui et al., 2014). The upstream region of VEGF was highly polymorphism, and many SNPs were found across the region of promoter and 5′UTR (Brogan et al., 1999; Awata et al., 2002). Szeto et al. (2004) found that the expression levels of VEGF mRNA were significantly lower in patients carrying the minor A allele of −2578C/A, while Cui et al. (2013) discovered that patients carrying the AA genotype and A allele of −2578C/A had more chances of CAD; this was consistent with our result that variation of −2578C/A could advance the incidence of TOF. Similarly, the A allele of −1154G/A could decrease the transcription of VEGF and affect the level of VEGF and advance the risk of TOF (Lambrechts et al., 2003). Meanwhile, functional studies have demonstrated that mutation of −634G/C was related with a lower level of VEGF and reducing transcription (Lambrechts et al., 2003). It is possible that patients with −634*C could inhibit angiogenesis and induce the etiology of TOF. In addition, +963G/A, which is the SNP of 3′UTR, played an important role in regulating expression of VEGF, and variation may also be the inducement of TOF risk. Since this case-control study involved people from the hospital, selection bias is inevitable. Besides, our study was based on a relatively small sample size. Moreover, we did not conduct stratification analyses between genetic variation and clinical characteristics, which may affect the conclusion. Therefore, further research needs to be conducted to confirm our findings.
In summary, our findings demonstrated that four SNPs (−2578C/A, −1154G/A, −634G/C, +963G/A) of VEGF were significant associated with increased TOF risk. Haplotype analysis in block 1 and block 3 also revealed significant association with TOF risk compared with the most common haplotypes. However, further study needs to validate the functional relationship between VEGF polymorphism and TOF susceptibility in various ethnics, which may contribute to the diagnosis and therapy of TOF.
Footnotes
Acknowledgment
This work was supported by the Natural Science Foundation of Heilongjiang Province (H201412).
Author Disclosure Statement
No competing financial interests exist.