
Abstract
Background: Macrophage apoptosis is a host innate defense mechanism against tuberculosis (TB). Aim: In this study, we aimed to investigate the role of microRNA-223 (miR-223) in macrophage apoptosis of TB. Methods: We analyzed apoptosis in peripheral blood macrophages of active TB patients, infected human macrophages (TDMs and MDMs) with the Mycobacterium tuberculosis (Mtb) strain H37Rv, and observed the expression of miR-223 to investigate the relationship between miR-223 and macrophage apoptosis induced by Mtb. Results: The apoptosis rate of peripheral blood macrophages decreased in active TB patients compared with healthy controls, and miR-223 expression increased significantly in macrophages after H37Rv infection. Transfection of human macrophages (TDMs and MDMs) with miR-223 inhibited macrophage apoptosis. We also demonstrated that miR-223 directly suppressed forkhead box O3 (FOXO3), and FOXO3 played a critical role as a mediator of the biological effects of miR-223 in macrophage apoptosis. The overexpression of FOXO3 remarkably reversed the apoptosis inhibitory effect of miR-223. Conclusion: Our data provide new clues for the essential role of miR-223 in the regulation of anti-Mtb-directed immune responses, which relies on the regulation of FOXO3 expression.
Introduction
T
Patients infected with Mtb develop a latent TB infection, which is a major source of active TB. The causative agent of TB, Mtb, is unanimously recognized as one of the most successful human pathogens because of its ability to persist and survive in the macrophages of immunocompetent individuals (Hmama et al., 2015). Mtb encounters the host innate immune defense, such as alveolar macrophages, during infection. Mtb first gains entry into resident alveolar macrophages following inhalation of infectious aerosols. Macrophages patrolling the distal airways avidly engulf inhaled bacteria using a variety of phagocytic receptors (Lee et al., 2009). Mtb circumvents the macrophage-killing machinery by blocking the fusion of mycobacterial phagosomes with lysosome, and Mtb replicates within macrophages (Rohde et al., 2007). Recent studies reported that virulent Mtb strains inhibit macrophage apoptosis and induce necrosis to spread the infection, but attenuated Mtb strains induce apoptosis, which suggests that macrophage apoptosis is a host innate defense mechanism against TB (Divangahi et al., 2009; Behar et al., 2010; Liu et al., 2013).
MicroRNAs are a class of genes that transcribe small, single-stranded, 19- to 24-nucleotide noncoding RNAs. MicroRNAs have become the focus of much research recently. MicroRNAs participate in a variety of biological processes, including cellular differentiation, apoptosis, metabolism, and proliferation, by silencing messenger RNAs (mRNAs) through binding to the 3′-untranslated region (3′-UTR) of target genes (Farh et al., 2005). The present study explored whether microRNAs play a role in the regulation of macrophages apoptosis in TB patients. Our results suggest that aberrant microRNA-223 (miR-223) expression is associated with macrophage apoptosis, which indicates that miR-223 plays an inhibitory role in macrophage apoptosis by targeting forkhead box O3 (FOXO3).
Materials and Methods
Human subjects
Thirty patients with active pulmonary TB and 30 healthy controls were recruited from the First Affiliated Hospital of Xinxiang Medical College, Xinxiang, China between 2012 and 2013 (Table 1). The diagnoses of pulmonary TB were all verified using acid-fast staining of sputum smear/bacterial culture, chest X-ray examination, clinical symptoms, and responses to anti-TB chemotherapy. The Ethics Committee of the First Affiliated Hospital of Xinxiang Medical College approved the study protocols, and informed written consent was obtained from all participants.
TB, tuberculosis; ESR, erythrocyte sedimentation rate; MDR, multidrug-resistant; XDR, extensively drug-resistant; BCG, bacillus Calmette-Guêrin; HIV, human immunodeficiency virus.
Cell culture and transfection
Human macrophages were derived from peripheral blood monocytes (MDMs) as described previously (Guerfali et al., 2008). The human myelomonocytic leukemia cell line THP-1 was obtained from the American Type Culture Collection (ATCC) and grown in Roswell Park Memorial Institute (RPMI) supplemented with 10% fetal bovine serum in humidified air at 37°C and 5% CO2. THP-1 cells were differentiated to macrophages (TDMs) using 20 ng/mL PMA as previously described (Daigneault et al., 2010). The miR-223 mimics and control miRNA mimics were purchased from Dharmacon (Austin, TX). Cells were transfected with the miRNA mimic (30 nM) or FOXO3 recombinant plasmid (200 ng) using DharmaFECT 1 (Dharmacon, Lafayette, CO) according to the manufacturer's instructions. All assays were performed 48 h after transfection.
Mycobacterial culture and infection
Mtb H37Rv was obtained from the ATCC and grown to log phase at 37°C in Difco Middlebrook 7H9 Medium (Difco Laboratories, Detroit, MI) supplemented with 0.05% Tween 80, 0.2% glycerol, and 10% oleic acid, albumin, dextrose, and catalase (OADC; Becton Dickinson Biosciences, Inc., Franklin Lakes, NJ). After culture of the mycobacteria under aeration, the culture was diluted to a concentration of 1.0 McFarland standard (108 bacilli/mL) and stored at −80°C. For infection experiments, human macrophages were infected with Mtb at multiplicity of infection (MOI) of 5:1 (5 bacteria/1 cell) for 3 h. Cells were then washed three times with complete RPMI medium to remove extracellular bacilli and cultured in humidified air at 37°C and 5% CO2 for 12 h. Relevant experiments were performed 12 h after infection.
Purification of primary human monocytes and flow cytometric analysis
Peripheral blood samples were drawn from patients with active TB and controls at 8:00 a.m. Peripheral blood mononuclear cells (PBMCs) were purified using density gradient centrifugation in Ficoll-Paque (GE Healthcare Biosciences, Pittsburgh, PA) within 6 h of blood collection. Anti-human CD3 and CD33 magnet beads (Miltenyi Biotec, Inc., Auburn, CA) were used to separate primary human monocytes. PBMCs from TB patients and controls were stained using fluorescein isothiocyanate (FITC)-labeled anti-human CD14 mAb (clone HCD14; BioLegend, San Diego, CA) and phycoerythrin (PE)-labeled anti-human CD206 mAb (clone 15-2; BioLegend) for 30 min at 4°C for surface staining. A proper homeotypic antibody was used to determine the background level of staining. At least 100,000 events were collected and analyzed using Beckman CXP software on a FC-500 Flow Cytometer (Beckman Coulter, Brea, CA). CD14+ and CD206+ content was analyzed using flow cytometry.
Apoptosis assay
The Annexin V-FITC Apoptosis Kit (BD Pharmingen™, San Diego, CA) was used to evaluate apoptosis. Briefly, cells were washed with cold phosphate-buffered saline and resuspended in 100 μL binding buffer at a density of 1 × 106 cells/mL. Annexin V-FITC and PI buffers were incubated with the cells for 15 min at 4°C in the dark immediately after resuspension. Annexin V-FITC and PI signals were detected using flow cytometry (Becton Dickinson Biosciences).
RNA extraction and microRNA expression assay
Total RNA was isolated using a mirVana™ miRNA Isolation Kit (Applied Biosystems, Carlsbad, CA), and cDNA synthesis was performed using a TaqMan® microRNA Reverse Transcription Kit (Applied Biosystems) according to the manufacturer's instructions. Expression of mature microRNAs was assayed using a TaqMan microRNA assay (Ambion, Carlsbad, CA) that was specific for the amplification and identification of miR-223. Real-time polymerase chain reaction (RT-PCR) was performed using iQ5™ instrument (Bio-Rad, Hercules, CA) with the following conditions: 50°C for 2 min, 95°C for 10 min, followed by 40 cycles at 95°C for 10 s and 56°C for 30 s. The relative quantification of miR-223 was calculated using the 2−ΔΔCt method. The relative amount of microRNAs was normalized using U6 as an internal control and measured relative to a calibrator sample as the external control.
Western blot assays
Cells were lysed on ice using RIPA buffer (100 mM 2-ME, 50 mM Tris-HCl [pH 6.8], 2% w/v sodium dodecyl sulfate [SDS] and 10% glycerol). Protein concentrations were measured using the bicinchoninic acid (BCA) method, and 30 μg of protein was separated using 10% SDS-polyacrylamide gel electrophoresis. Separated proteins were transferred onto polyvinylidene fluoride membranes (Millipore, Brussels, Belgium) for analysis. A human FOXO3 was used as the primary antibody (final dilution 1:500; Santa Cruz Biotechnology, Dallas, TX). A GAPDH antibody (final dilution 1:1000; Cell Signaling Technology, Boston, MA) was used as a control. Bands were detected using an Enhanced Chemiluminescence Kit (GE Healthcare Biosciences) and visualized using the ChemiDoc XRS system (Bio-Rad).
Plasmid construction
A DNA fragment of the 3′-UTR region of FOXO3, which contains the potential target sequence of miR-223, was amplified using PCR. The primers used contained the following restriction sites: FOXO3 3′UTR forward, 5′-ACTAGTAGGATCACTGAGGAA-3′ and reverse, 5′-AAGCTTTGGTACAACATTTTC-3′. The following FOXO3 3′-UTR mutation oligos were synthesized: Mut FOXO3 3′UTR forward, 5′-AGCAGACCCTCGGATTGACACAAGACCTAC-3′ and reverse, 5′-GTAGGTCTTGTGTCAATCCGAGGGTCTGCT-3′. The PCR product from the FOXO3 3′UTR was cloned into the SpeI and HindIII restriction sites downstream of the luciferase open reading frame in the pMIR-REPORT vector (Ambion).
FOXO3 without the 3′UTR was amplified using PCR with the following primers: forward, 5′-ATGGCAGAGGCACCGGCTTCC-3′ and reverse, 5′-TCAGCCTGGCACCCAGCTCTGA-3′. The resulting PCR amplicons of FOXO3 were cloned into the T vector (Promega, Madison, WI). Sequences confirmed the correct clones.
Luciferase assays
Human macrophages (MDMs and TDMs) were seeded into a 24-well plate (1.5 × 105 per well) and cotransfected with miRNA mimics (50 nM) and a reporter recombinant plasmid (100 ng) through electroporation (Amaxa Nucleofection system) according to the manufacturer's instructions. Luciferase activity was analyzed 24 h after cotransfection using a luminometer (EG&G Berthold; PerkinElmer Life Sciences, Boston, MA), and values were normalized to the protein concentration of the total cell lysate.
Statistical analyses
Values are shown as the mean ± SD for at least three repeated individual experiments per group. Significant differences were analyzed using Student's t-test or Mann-Whitney U test for comparisons between two groups. Differences were considered statistically significant at p < 0.05.
Results
Mtb suppresses apoptosis and induces miR-223 expression in human macrophages
Peripheral blood was obtained from 30 patients with active TB and 30 healthy controls, and macrophage frequencies were determined using flow cytometric analyses of CD206 and CD14 expression. Patients with active TB exhibited a remarkably increased frequency of macrophages compared with healthy controls (Fig. 1A). However, flow cytometry indicated that the percentage of apoptotic macrophages was significantly reduced in patients with active TB (Fig. 1B). We investigated differentially expressed microRNAs in patients with active TB to determine whether microRNA modulate macrophage apoptosis. Our microRNA expression assay showed that miR-223 in peripheral blood macrophages was significantly upregulated in patients with active TB compared with healthy controls (Fig. 1C).
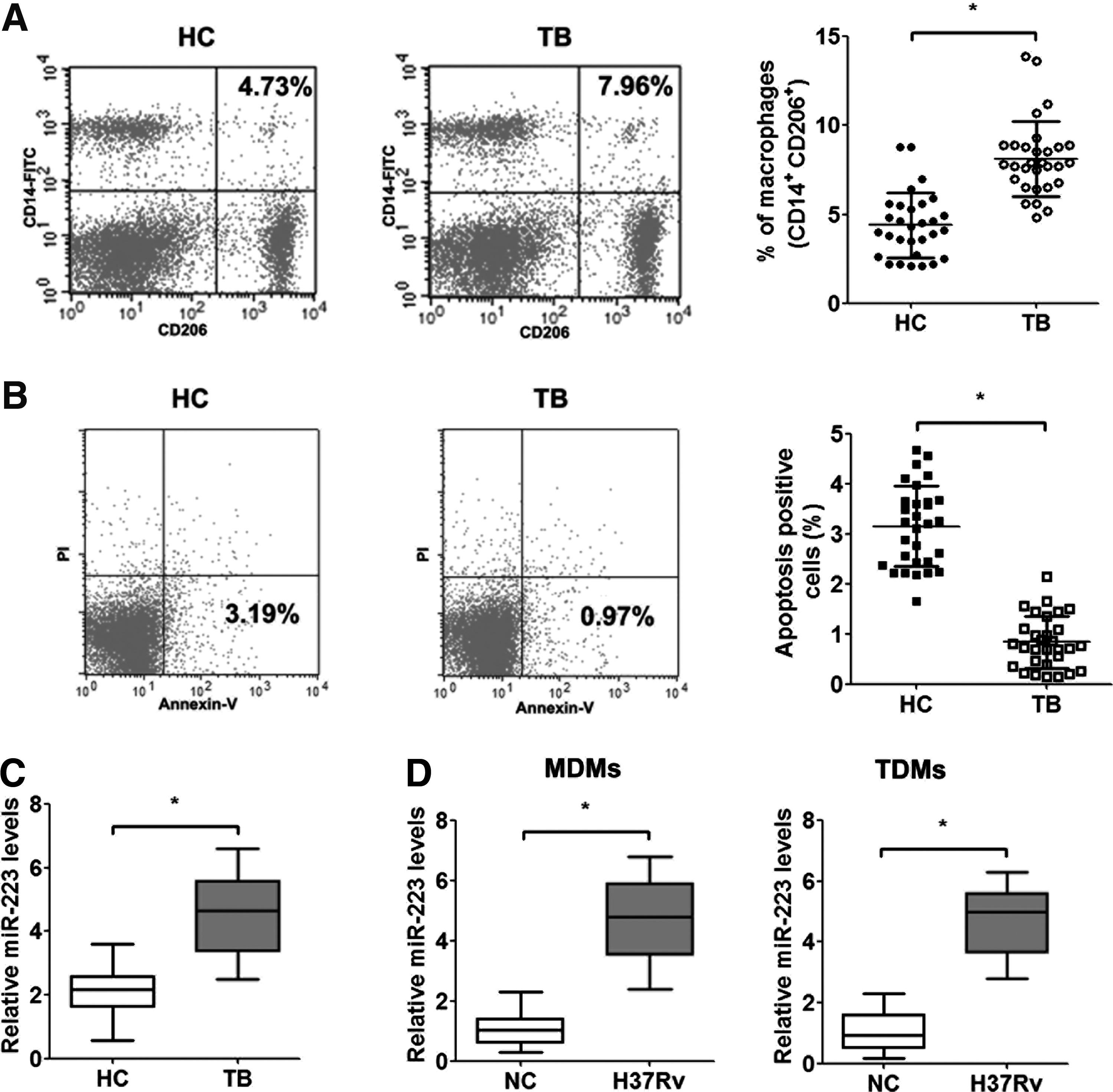
Impact of Mtb on apoptosis and miR-223 expression in human macrophages.
Moreover, we used the virulent Mtb strain H37Rv to infect human macrophages (MDMs and TDMs) to assess the effect of Mtb infection on macrophage survival and found that miR-223 expression was significantly elevated after infection (Fig. 1D). The results suggested that Mtb infection induced the expression of miR-223 in vivo and in vitro.
miR-223 inhibits human macrophage apoptosis
MiR-223 was overexpressed in human macrophages (MDMs and TDMs) using transfection with miR-223 mimics to further elucidate the role of miR-223 in human macrophage apoptosis. RT-PCR results demonstrated that transfection with miR-223 mimics drastically increased miR-223 levels in MDMs and TDMs (Fig. 2A, C). The overexpression of miR-223 in macrophages significantly inhibited the rate of macrophage apoptosis compared with the control group (Fig. 2B, D). These data suggested that miR-223 inhibited human macrophage apoptosis.
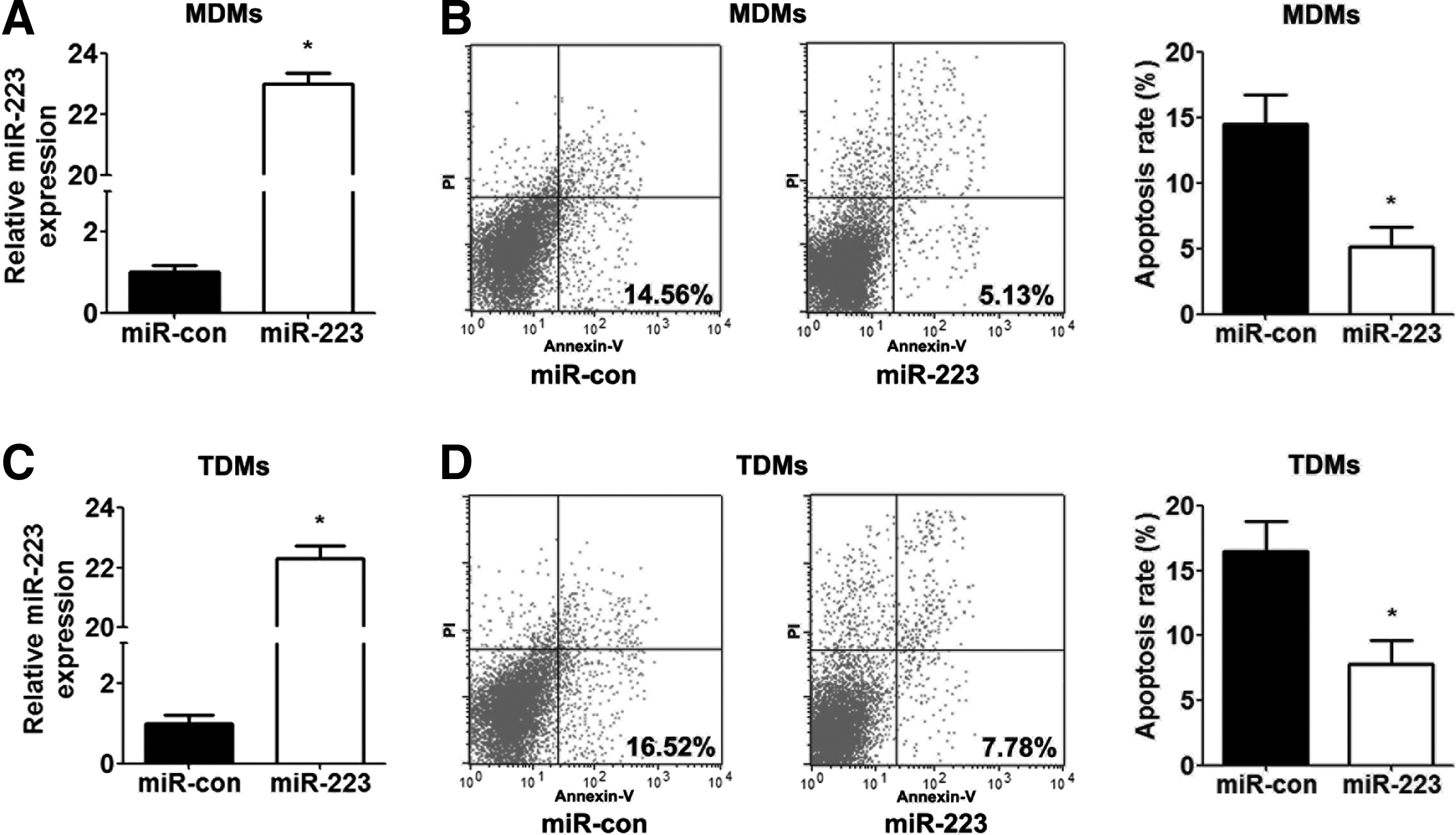
The effect of miR-223 on human macrophage apoptosis. MDMs and TDMs were transfected with control mimics or miR-223 for 48 h.
FOXO3 is a direct target of miR-223 in human macrophages
Systematic bioinformatics analyses suggested that FOXO3 is a target gene of miR-223, and its 3′UTR contains potential binding sites for miR-223. One recent study reported that FOXO3 is a target of miR-223 posttranscriptional repression (Wu et al., 2013). We transfected MDMs and TDMs with miR-223 mimics to demonstrate whether miR-223 regulated FOXO3 in macrophages. Western blot analyses demonstrated that miR-223 mimics significantly reduced the expression of FOXO3 protein in macrophages (Fig. 3A). We analyzed the potential seed sequence for miR-223 in the 3′UTR region of FOXO3 and cloned the wild-type and mutant FOXO3 3′UTR fragments into a luciferase reporter gene system to verify whether FOXO3 was a direct target of miR-223 (Fig. 3B). Wild-type or mutant FOXO3 3′-UTR constructs were cotransfected with miR-223 or control mimics into MDMs and TDMs, and luciferase activity was measured. The overexpression of miR-223 in human macrophages significantly reduced luciferase activity for the wild-type constructs, but no alterations in luciferase activity were detected with the mutant FOXO3 3′-UTR luciferase reporter plasmid (Fig. 3C, D). These results indicated that miR-223 inhibited macrophage apoptosis through the direct downregulation of FOXO3.

miR-223 negatively regulates FOXO3 expression in human macrophage. MDMs and TDMs were transfected with control mimics or miR-223 for 48 h.
FOXO3 overexpression reverses the apoptotic inhibitory effect of miR-223
We transfected MDMs and TDMs overexpressing miR-223 with a FOXO3 recombinant plasmid that lacked the respective 3′UTR or an empty vector to determine whether FOXO3 mediated miR-223 repression of macrophage apoptosis. Western blot analyses demonstrated an increased expression of FOXO3 proteins in macrophages cotransfected with this FOXO3 construct compared with macrophages transfected with the empty vector (Fig. 4A). Cell apoptosis assays demonstrated that the ectopic expression of FOXO3 abolished miR-223-induced apoptosis in macrophages (Fig. 4B, C).
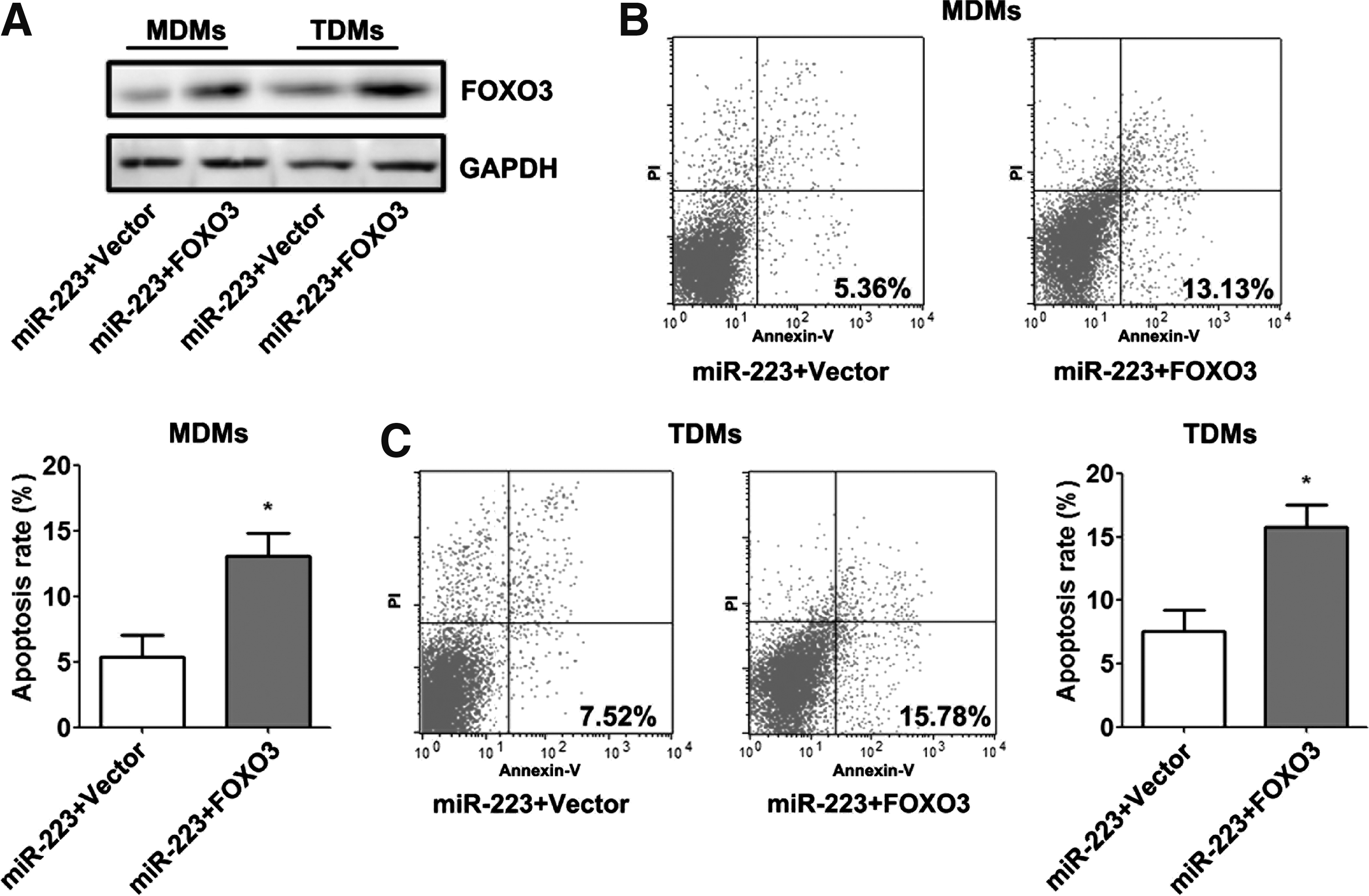
Ectopic expression of FOXO3 reverses the apoptotic inhibitory properties of miR-223. MDMs and TDMs were cotransfected with control mimics or miR-223 mimics and the empty vector or FOXO3 recombinant plasmid for 48 h.
Discussion
The ability of virulent Mtb to survive within the intracellular environment of host mononuclear phagocytes has been a major focus of TB research (Toossi et al., 2012). The capacity of Mtb to suppress normal phagosome maturation and acidification was the first identified mechanism of this facilitated intracellular survival for Mtb (Rook and Hernandez-Pando, 1996). However, recent studies demonstrated that virulent Mtb also inhibited phagocyte apoptosis to promote its own survival (Spira et al., 2003; Rios-Barrera et al., 2006). Keane et al. (1997) first reported that the infection of human alveolar macrophages was sufficient to induce classical, extrinsic apoptosis in 1997. Subsequent studies confirmed the inverse correlation of Mtb virulence and the induction of classical apoptosis at low MOI (Keane et al., 2000). The apoptosis of Mtb-infected macrophages was hypothesized as a defensive response. This hypothesis was further supported by the discovery that virulent Mtb strains actively suppress apoptosis by interfering with tumor necrosis factor alpha signaling and upregulating the expression of antiapoptotic Mcl-1 in a TB model (Balcewicz-Sablinska et al., 1998; Sly et al., 2003; Spira et al., 2003).
The current results are consistent with previous studies that demonstrated that peripheral blood macrophages from patients with active TB included a significantly lower percentage of apoptotic cells than healthy controls. These data indicate that virulent Mtb infection can increase the percentage of macrophages and suppress macrophage apoptosis in the peripheral blood of active TB patients, which may play a role in host immunity against TB and the augmentation of circulating macrophages in TB patients.
Accumulating studies identified microRNAs as effective biomarkers for use in TB prevention, diagnosis, and treatment, including markers that discriminate between healthy individuals and people with latent infection (Harapan et al., 2013; Iannaccone et al., 2014; Mehta and Liu, 2014; Zhang et al., 2014). Studies in the past decade noted that miR-223 plays a crucial role in the regulation of tumor characteristics (Wei et al., 2014), epithelial-mesenchymal transition (Ma et al., 2015), obesity (Kilic et al., 2015), differentiation of mesenchymal stem cells (Guan et al., 2015), and immature neurons (Harraz et al., 2014). Recently, miR-223 was identified as one of the most upregulated microRNAs in the peripheral blood of patients with active TB, and miR-223 is recognized as a regulator that controls susceptibility to TB through regulation of lung neutrophil recruitment (Dorhoi et al., 2013). This study explored the quantity of macrophages in active TB patients in relation to miR-223 expression level. We found that miR-223 in peripheral blood macrophages of active TB patients and miR-223 in human macrophages infected by H37Rv increased significantly, which suggested that mycobacterial infection elevated miR-223 levels.
FOXO3 belongs to the FOXO subfamily of forkhead transcription factors, which contains evolutionarily conserved transcriptional activators that are characterized by a highly conserved forkhead domain with a DNA-binding motif (Greer and Brunet, 2005). A previous report demonstrated that FOXO3 mediated apoptosis in BCG-infected macrophages, which suggests that FOXO3 expression may be one of the many critical events that occur in macrophage apoptosis (Haoues et al., 2014). Emerging evidence shows that miR-223 negatively regulates FOXO3 by directly targeting the 3′UTR of FOXO3 mRNA (Wu et al., 2013). The current study validated FOXO3 as a direct target gene for miR-223 through the prediction of molecular information and demonstrated that miR-223 negatively regulated FOXO3 expression by directly targeting the 3′UTR of FOXO3 mRNA in human macrophages. We also demonstrated that FOXO3 overexpression reversed the inhibitory effect of miR-223 on macrophage apoptosis, which suggests that miR-223 inhibited macrophage apoptosis by suppressing FOXO3 expression.
In summary, our study demonstrated that the percentage of macrophages in the peripheral blood of patients with active TB increased as a result of apoptosis suppression, which was induced by miR-223 expression. The inhibitory effect of miR-223 on macrophage apoptosis occurred through the regulation of FOXO3 gene expression. These novel findings may be used as a prognostic marker and therapeutic target in TB patients.
Footnotes
Acknowledgments
The authors are grateful to the staff at the First Hospital Affiliated to the Xinxiang Medical College and the participating health institutions. Their special thanks go to the TB patients and all of the volunteers who consented to participate in the survey.
Author Disclosure Statement
No competing financial interests exist.