
Abstract
Aims: This study aimed to compare the intron 4 sequence of the RHD and RHCE genes from Han Chinese, Tibetans, and Mongols, and explore its polymorphisms. Materials and Methods: To investigate the distinction in the RHD and RHCE intron 4, polymerase chain reaction (PCR) was performed by a set of primers: Intron4F and Intron4R. Primer Intron4F for a sequence located in exon 4 and primer Intron4R for a sequence located in exon 5, respectively. RHD and RHCE intron 4 of all the samples from 26 cases of random unrelated Hans (13 RhD-positive donors and 13 RhD-negative donors), 25 cases of random unrelated Tibetans (18 RhD-positive donors and 7 RhD-negative donors), and 4 cases of random unrelated Mongols (1 RhD-positive donor and 3 RhD-negative donors) were amplified with PCR. The PCR products were then sequenced. Results: A 576-bp product was detected in all the Han, Tibetan, and Mongol RhD-positive donors, whereas a 1228-bp product was detected in RhD-negative donors. The sequences of RHCE gene intron 4 were identical to each other in all Han, Tibetan, and Mongol RhD-negative donors, including 335 bp of Alu element, with a whole length of 1078 bp. By contrast, a 426-bp product was detected in all Han, Tibetan, and Mongol RhD-positive donors. Compared with the RHCE gene, a 652-bp deletion was noted in the RHD gene of Chinese, including the whole Alu element. The results were similar to the findings of Caucasians, whereas the lengths of RHD gene deletion fragments of Japanese and French were 649 and 654 or 651 bp, respectively. Conclusions: The RHCE gene intron 4 of Han Chinese, Tibetans, and Mongols differs from the RHD gene intron 4 in the presence of a 652-bp fragment. The RHCE gene intron 4 in Chinese has its own structural characteristics and differs among various ethnicities and regions.
Introduction
R
Materials and Methods
Blood samples
A total of 26 cases of random unrelated Hans (13 RhD-positive donors and 13 RhD-negative donors) were provided by the Blood Bank of Guangdong Provincial Hospital of Traditional Chinese Medicine. A total of 25 cases of random unrelated Tibetans (18 RhD-positive donors and 7 RhD-negative donors) were provided by the Tibet Autonomous Region Blood Center. Four cases of random unrelated Mongols (one RhD-positive donor and three RhD-negative donors) were provided by the Blood Center of Shanxi Province. All the donors resided in their native lands and did not intermarry with other ethnic groups in at least three generations. Whole blood was drawn from the cubital vein after informed consent was obtained. Ethylenediaminetetraacetic acid blood samples were collected and genomic DNA was extracted (Generation Capture Column Kit and DNA Extraction Kit; Qiagen, Inc., Hilden, Germany).
RhD antigen detection
Serotyping for the RhD antigen was carried out by the agglutination test with monoclonal anti-RhD (IgM) (Shanghai Blood Biological Pharmaceutical Co., Ltd., Shanghai, China), Coombs test with anti-RhD (IgG) and polyclonal antiglobulin reagent (Shanghai Blood Biological Pharmaceutical Co., Ltd.), and absorption/elution test with anti-RhD (IgG) (Shanghai Blood Biological Pharmaceutical Co., Ltd.) and ether (Luoyang Chemical Reagent Co., Ltd., Luoyang, China).
Amplification of RHD and RHCE intron 4
As described by Okuda et al. (1997), the amplification strategy of RHD and RHCE intron 4 is shown in Figure 1.
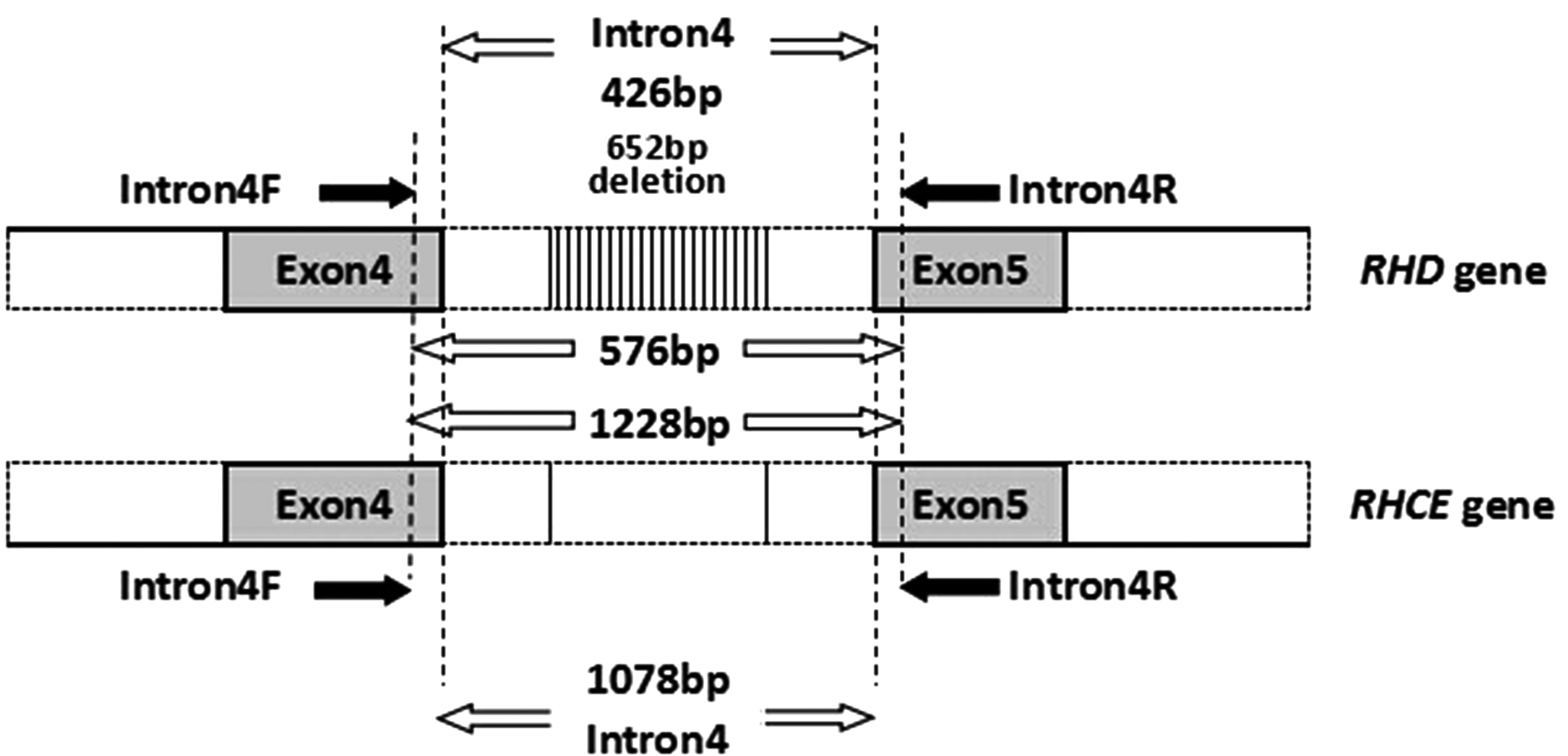
Primers Intron4F/Intron4R amplified a 1228-bp fragment of the RHCE gene and a 576-bp fragment of the RHD gene in intron 4.
To investigate intron 4 in the RHD and RHCE genes, polymerase chain reaction (PCR) amplification was performed by a set of primers: Intron4F (5′-TGTTCGCAGCCTATTTTGGG-3′) and Intron4R (5′-TGACACTTGGCCAGAACATC-3′). Primer Intron4F for a sequence located in exon 4 and primer Intron4R for a sequence located in exon 5, respectively. The final concentration of the PCR mixture consisted of 1× buffer (Mg2+-free), 1.5 mM MgCl2, 320 mM dNTPs, 2 U Taq DNA polymerase (Promega Co., Ltd., Shanghai, China), 50-200 ng of genomic DNA, and 0.24 mM of both primers Intron4F and Intron4R in a total volume of 50 μL. PCR was conducted as follows: denaturation at 95°C for 5 min; 35 cycles of 40 s at 95°C, 40 s at 55°C, and 2 min at 72°C; and final cycle at 72°C for 5 min. The reaction products were electrophoresed on 1% agarose gel containing 0.5 μg/mL ethidium bromide.
Sequencing
RHD and RHCE intron 4 were sequenced by dideoxy chain termination using an Applied Biosystems 3730 DNA analyzer (Applied Biosystems, Inc., Foster City, CA) and using the intron 4 amplification primers.
Results
Determination of Rh phenotypes
In this study, Rh phenotyping was performed by agglutination test, Coombs test, and absorption/elution test. RhD-negative phenotypes were determined through antiglobulin and adsorption/elution tests. We analyzed 26 Han Chinese, 25 Tibetan, and 4 Mongol donors. The RhD-positive phenotypes consisted of 13 Hans, 18 Tibetans, and 1 Mongol, whereas the RhD-negative phenotype donors consisted of 13 Hans, 7 Tibetans, and 3 Mongols.
Detection of RHD gene intron 4
All 32 RHD-positive samples showed a 576-bp PCR product; however, the 1228-bp PCR product did not appear due to PCR inhibition (Fig. 2).

To evaluate the results of special amplification by polymerase chain reaction, the products were electrophoresed on 1% agarose gel and stained with ethidium bromide. Intron 4 was amplified in both the RHD and RHCE genes with a size of 576 (lanes 1-6) and 1228 bp (lanes 7-10), respectively. M shows the marker.
Detection of RHCE gene intron 4
In all the 23 RhD-negative samples, a 1228-bp PCR product appeared, rather than a 576-bp PCR product (Fig. 2).
Sequencing of the RHD and RHCE genes
Sequence alignment showed that the sequence of RHCE gene intron 4 was completely consistent in the Han, Tibetan, and Mongol RhD-negative samples. The whole length of RhD-negative RHCE gene intron 4 is 1078 bp, and it contains an Alu sequence with a size of 335 bp as previously described (Kohany et al., 2006). The sequences of RHD gene intron 4 of Han, Tibetan, and Mongol RhD-positive samples were also consistent, with a size of 426 bp, including the Alu fragment; the size of RHD gene intron 4 was 652-bp smaller than that of the RHCE gene in the three populations.
Sequence contrast of RHD and RHCE intron 4 among Chinese and other ethnic groups
According to the report of Okuda et al. (1997), the sequence of RHCE gene intron 4 among Chinese was found to be consistent with that among Caucasians, but different from that among Japanese in five bases. However, we noted five nucleotide differences: three insertions (A at nucleotide 413, G at nucleotide 418, and A at nucleotide 497) and two substitutions (C→T at nucleotide 644, G→A at nucleotide 802) in intron 4 of the RHCE gene (Fig. 3). The sequences of Chinese PCR products were different from those of French PCR products. The size of the deletion sequence in intron 4 of the RHD gene was 652 and 654 or 651 bp in Chinese and France populations, respectively (Apoil et al., 1999; Apoil and Blancher, 2000). RHD and RHCE intron 4 of Han Chinese and Tibetan populations are shown in Figure 4.
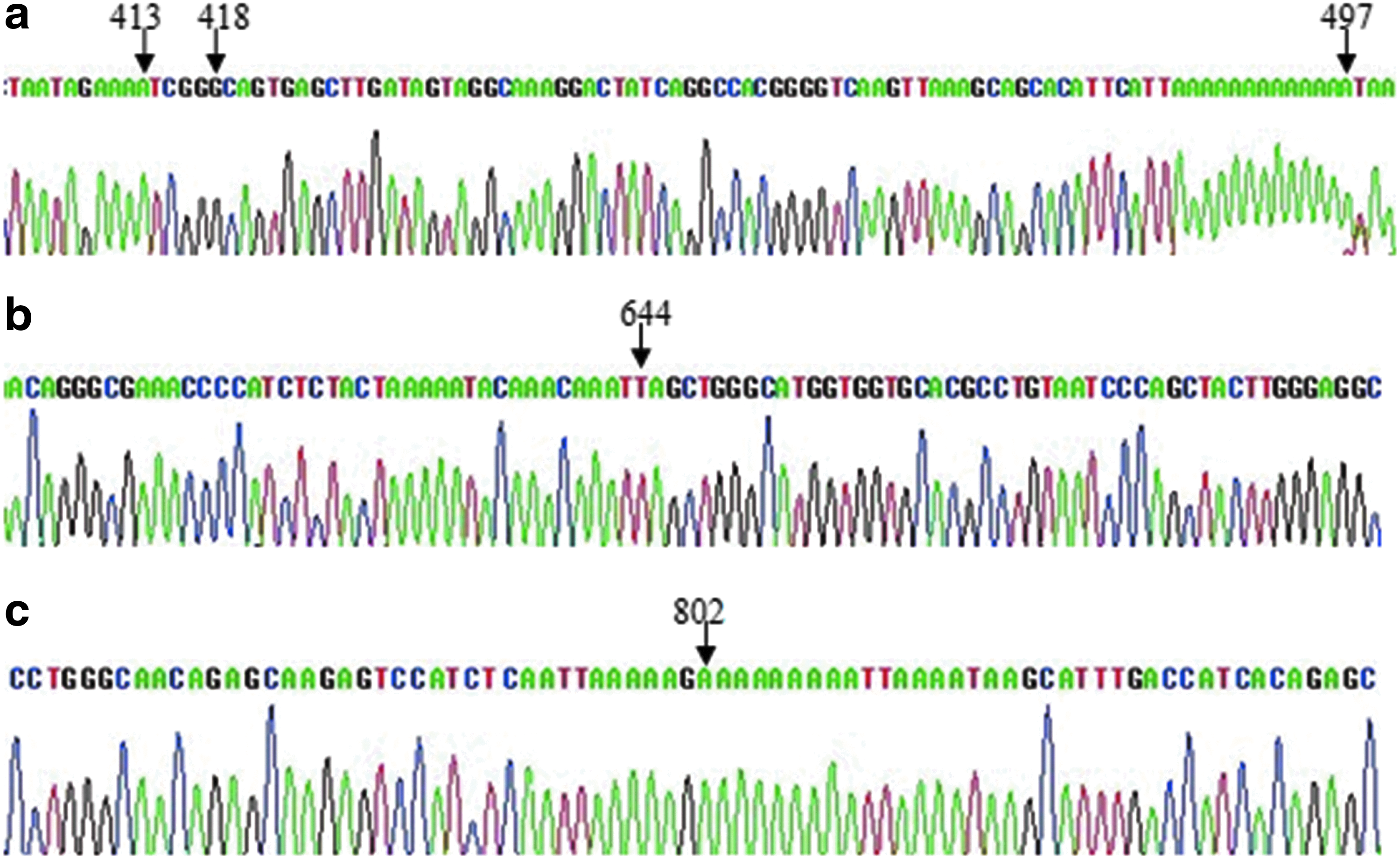
Three insertions (A at nucleotide 413, G at nucleotide 418, and A at nucleotide 497

The intron 4 sizes of the RHCE and RHD genes of a Chinese population were 1078 and 426 bp, respectively. The underlined letters indicate the 652-bp deletion of the RHD gene, in which the double-underlined fragment indicates the Alu sequence. The letters surrounded with an oblong frame demonstrate the three nucleotide insertions in Chinese, whereas the two italicized letters show the nucleotide substitutions between the Chinese and Japanese.
Discussion
Systematic studies and sporadic observations indicated that the RH is one of the most complex blood group systems. Apoil and Blancher (2000) studied RHD and RHCE evolution in primates. Their research group demonstrated that chimpanzees and gorillas possess two types of RH intron 4, one differing from the other by a 654-bp deletion, encompassing an Alu element. One type has a size similar to that of human RHD intron 4, whereas the other displays a 12-bp insertion (Apoil and Blancher, 2000). A number of chromosomal polymorphisms of RHD and RHCE genes in primates suggested that unequal crossing over most probably arose after the original duplication of the RH ancestor gene. The RHCE and RHD genes differ in their coding regions and noncoding sequence, including the length polymorphisms of intron 4 (Arce, 1993). In humans, intron 4 of the RHCE gene differs from intron 4 of the RHD gene in the presence of an approximate 650-bp fragment that encompasses an Alu element (Westhoff and Wylie, 1996; Huang, 1997; Okuda et al., 1997). However, various deletions in the RHD gene among different regions and populations exist. Sequencing analysis of RHD and RHCE intron 4 indicated that Chinese, Caucasian, Japanese, French, and British populations have similarities and differences. The data obtained clearly demonstrated that RHD and RHCE intron 4 were exactly the same among Han Chinese, Tibetan, and Mongol populations. However, despite the geographical proximity of Japan and China, similarities and differences were present. A large sample size should be used in future studies to learn more about the RHD and RHCE among populations of Chinese, Japanese, and other regions for constructing a genetic tree.
Our study observed the Han Chinese, Tibetan, and Mongol populations. Compared with the RHCE gene, the RHD gene intron 4 is missing an entire Alu sequence. The genetics and biological significance of the difference between the two genes remain unclear. On cytogenetic levels, the Alu sequence is often concentrated in the active chromosome segments of gene transcription; it may help in inserting and restructuring mutagenesis in primate genomes, and various potential mechanisms may mediate gene deletions (Deininger and Batzer, 1999). Studies on RHD and RHCE intron 4 of a variety of primates showed that the Alu sequence exists in the RHD and RHCE of the common ancestor of human, apes, Old World monkeys, and New World monkeys (Apoil and Blancher, 2000). Deininger and Batzer (1999) pointed out that the Alu sequence not only contributes to some genetic evolution but also, its range-mediated homologous recombination, may promote human genetic diseases. The Alu sequence, a molecular fossil, can provide DNA recombination and phylogenetic information of primates (Andreassen, 2004). However, the Alu sequence in RHCE gene intron 4 has real functions and its influence remains unclear. We speculated that the Alu sequence in the RHCE genes plays a certain important role in the process of RH genetic evolution (Rudiger et al., 1995), such as mediating about 650-bp deletion of the RHD gene, and it may be associated with alternative splicing of exons.
In conclusion, the numerical genetic polymorphism of RHD and RHCE genes in primates suggested that unequal crossing over arose most probably after the duplication of the ancestor RHD and RHCE genes. Various mechanisms of intergenic exchanges, such as crossing over, gene conversion, or homologous recombination between RHD and RHCE genes, have occurred in humans and other species (i.e., gorillas and chimpanzees). The major difference between the RHD and RHCE genes is intron 4. The polymorphisms of intron 4 and whether it is influenced by natural selection need further evolutionary studies.
Footnotes
Acknowledgment
This work was supported by fund project: National Natural Science Foundation of China (30971580).
Author Disclosure Statement
No competing financial interests exist.