
Abstract
Background: The human chitinase chitotriosidase enzyme, which is encoded by the CHIT1 gene, is produced by macrophages, and may be important in immune responses to chitin-containing organisms, such as fungi. Plasma chitotriosidase activity is used to diagnose and monitor some forms of lysosomal storage disorders, such as Gaucher's and Niemann-Pick disease. However, homozygous duplication of a 24-bp region in exon 10 of the CHIT1 gene eliminates enzyme activity and may complicate disease monitoring. The high prevalence of this mutation highlights the need to determine its frequency in different populations and screen patients for this mutation to verify whether chitotriosidase activity is a reliable marker of lysosomal storage disease. This study investigated the allele frequency of the 24-bp duplication in the general Iranian population. Methods: To identify the 24-bp duplication in exon 10 of the CHIT1 gene (H allele), genotyping of DNA extracted from peripheral blood leukocytes of 577 healthy Iranians was performed using polymerase chain reaction (PCR) amplification and high resolution melting (HRM) PCR techniques. Results: In this study, heterozygous and homozygous duplications were detected in 183 (31.7%) and 35 (6.1%) subjects, respectively. In addition, the allelic frequency was 21.9% (95% confidence interval). Conclusion: Our study indicates that genotype analysis by HRM-PCR is a fast, reliable, and highly accurate screening approach for identifying the 24-bp duplication in CHIT1 exon 10. Due to the wide range of duplication frequencies among different ethnic groups, new biomarkers are necessary for assessing genetic characteristics of lysosomal storage disorders in different populations.
Introduction
H
One of the most common lysosomal storage disorders is Gaucher's disease, which results from autosomal recessive defects in acid β-glucosidase (glucocerebrosidase). Gaucher's disease is also associated with an accumulation of uncleaved membrane glycolipids in macrophages. Since the chitotriosidase activity is significantly increased in the plasma and tissues of Gaucher's disease patients, it is important to determine whether the chitotriosidase activity is a reliable disease marker because it has been shown that the enzyme activity has a direct correlation with the number of Gaucher cells and disease severity (Hollak et al., 1994; Guo et al., 1995; Scriver et al., 2001). Treatment of Gaucher patients with enzyme replacement therapy (ERT) can gradually reduce the level of chitotriosidase (Cabrera-Salazar et al., 2004; Lee et al., 2012), suggesting that this may be a useful marker for disease diagnosis and an indicator of response to therapy. Although other secondary markers are available for Gaucher's disease (Deegan et al., 2005), measuring chitotriosidase activity levels is still an important approach for monitoring patients treated with ERT.
Chitotriosidase is encoded by the CHIT1 gene, which is located on chromosome 1 at 1q31-q32 and contains 12 exons that span ∼20 kb of genomic DNA (Boot et al., 1998; Fusetti et al., 2002). An inherited recessive mutation in the CHIT1 gene, which produces a 24-bp duplication in exon 10, subsequently activates a cryptic 39-bp splice site (Malaguarnera, 2006). The resulting spliced mRNA codes for an enzymatically inactive CHIT1 protein that lacks amino acids 344-372; this mutation is the predominant cause of CHIT1 defects that occur in people of different ethnic races (Boot et al., 1998; Piras et al., 2007). In the subjects having the mutant allele, the activity of chitotriosidase is undetectable in all materials tested, including plasma, urine, leukocytes, and tissues, indicating that this mutation is the predominant cause of chitotriosidase deficiency. Also, cultured macrophages of chitotriosidase-deficient individuals contain very little mRNA and secrete almost no chitotriosidase protein. The level of plasma chitotriosidase in carriers of the 24-bp duplication is approximately half of that observed in individuals carrying the wild-type CHIT1 gene (Schoonhoven et al., 2007). Approximately, 30-40% of the Caucasian population is heterozygous for this mutation, while ∼6% are homozygous with a complete absence of chitotriosidase activity (Rodrigues et al., 2004; Zheng et al., 2005). As such, determining the mutation frequency in different geographical populations and identifying heterozygosity or homozygosity of this mutation is necessary to account for the high frequency of chitotriosidase activity defects, which may compromise the utility of this enzyme as a biomarker.
The purpose of this study was to determine the frequency of the 24-bp duplication in exon 10 of the CHIT1 gene in the general Iranian population by using different mutation analysis methods and identifying which methods provide the simplest, most accurate, and efficient means of detecting mutations in CHIT1.
Materials and Methods
Subjects and CHIT1 assay
The present study was performed on blood samples from 577 Iranian individuals, who were selected based on their medical history and absence of the specific disease. Informed written consent was obtained from the study participants. The chitotriosidase enzyme activity was determined by the fluorometric assay (Hollak et al., 1994), and the activity range was used to detect heterozygosity of the 24-bp CHIT1 duplication.
CHIT1 genotyping analysis by polymerase chain reaction
Genomic DNA was extracted from peripheral blood leukocytes using a standard phenol-chloroform method and a commercial kit from SinaClon Co.
To detect 24-bp CHIT1 duplication mutations, the genomic DNA was first subjected to polymerase chain reaction (PCR) using the primer sequences shown in Table 1. PCR amplification was performed using 500 ng of DNA, 0.5 μL of 10 mM dNTPs, 500 nM of each primer, 0.75 μL 50 mM MgCl2, 2.5 μL 10× PCR Buffer, 0.2 U Hot Start Taq DNA polymerase (SinaClon), and 18 μL sterile H2O in a final volume of 25 μL (Tamanaha et al., 2013). Amplification was carried out with an Eppendorf Gradient Thermocycler using the following protocol: initial denaturation for 5 min at 95°C, 30 cycles of 95°C for 45 s, 60°C for 45 s, and 72°C for 1 min, followed by an extension step at 72°C for 5 min. The resulting 99-bp (mutant allele) and 75-bp (wild-type allele) fragments were separated on a 3% agarose gel and stained with ethidium bromide (Fig. 1).
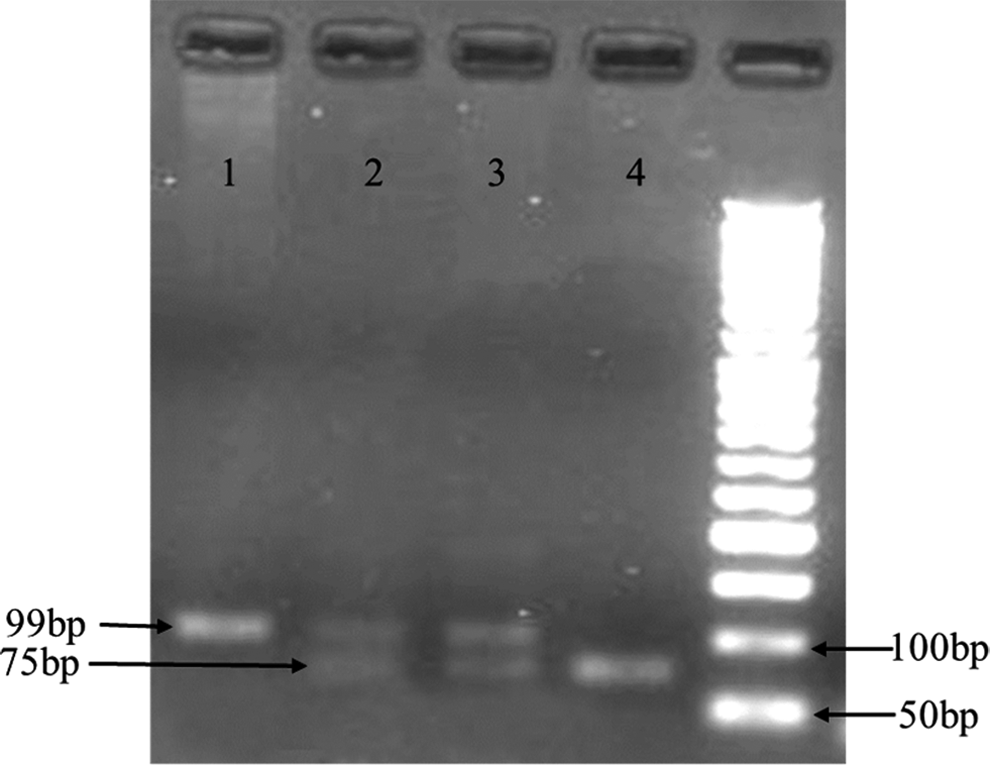
Polymerase chain reaction (PCR) products of CHIT1 polymorphisms on a 3% agarose gel. Lane 1, homozygote mutant (99-bp); lanes 2 and 3, heterozygote wt/mutant (99- and 75-bp); lane 4, homozygote wild type (75-bp).
PCR, polymerase chain reaction.
CHIT1 genotyping method by PCR-HRM
The 24-bp duplication mutation in the CHIT1 gene was also analyzed by PCR-HRM (high resolution melting), which was performed using a Rotor-Gene™ 6000 real-time rotary analyzer (Corbett Life Sciences). The primer sequences used were previously described. The reactions were prepared in 20 μL volumes in 0.2 mL strip tubes of a 36-well rotor. For symmetric PCR-HRM, the amount of DNA in each reaction was adjusted to 50 ng. Each reaction contained 4 μL of 5× HRM PCR Master Mix (Type-it HRM PCR kit; QIAGEN), 2 μL (250 nM) of primer mix, 1.2 μL (50 ng) of DNA, and DNA RNase-free water to a final volume of 20 μL. The PCR cycling profile was as follows: 15 min at 95°C for initial denaturation, then 40 cycles of 95°C for 15 s, 60°C for 60 s, and 72°C for 20 s. After amplification, HRM data were collected from 60°C to 90°C in 0.2°C increments, followed by a waiting time of 10 s.
Melting curves for PCR-HRM were analyzed by a post-PCR procedure using software supplied with the apparatus.
Results
CHIT1 activity
The activity level of plasma CHIT1 was higher in individuals who were homozygous for the wild-type allele compared to those who were heterozygous for the mutant allele (21.7 ± 10.78 vs. 11.33 ± 6.98 nmol/h/mL, respectively, p = 0.0019; Fig. 2). In addition, individuals who were homozygous for the mutant allele showed no plasma CHIT1 activity.

Plasma chitotriosidase activity in wild-type individuals and those carrying the CHIT1 24-bp duplication. As it is often the case between wt homozygous and heterozygous individuals, overlap could be seen between the respective values of enzyme residual activity.
Allele frequency of the 24-bp duplication in exon 10 of the CHIT1 gene
The genotypic and allelic frequency results are shown in Table 2. The study sample population was in Hardy-Weinberg equilibrium (p > 0.05). The frequency of the 24-bp allele duplication was 21.9% and the frequency of the wild-type allele was 78.1% (95% confidence interval). Genotype frequencies for the homozygous H allele as well as the heterozygous and homozygous wild-type alleles were 6.1%, 31.7%, and 62.2%, respectively.
wt, wild type.
Detection of the 24-bp duplication in the CHIT1 gene by electrophoresis is shown in Figure 1. PCR-HRM techniques were performed and the melting curves derived from this technique were plotted using software included with the apparatus and used to evaluate the 24-bp duplication (Fig. 3). A notable feature of the technique is its ability to differentiate the products from the two alleles, such that wild-type, heterozygous, and homozygous individuals are easily recognizable from the different melting temperatures shown in the three PCR-HRM curves (Fig. 3). The heterozygous genotype can be identified clearly by the two peaks in the temperature range of the wild-type and homozygous genotypes. The results from HRM techniques are corroborated with the results of conventional PCR.

Discussion
The 24-bp duplication in exon 10 of the CHIT1 allele causes aberrant mRNA splicing that results in an inactive chitotriosidase enzyme (Piras et al., 2007). In our study, the frequency of this duplication was 21.9%, and the frequencies of heterozygous and homozygous allele carriers were 31.7% and 6.1%, respectively. These results are consistent with previous studies (Boot et al., 1998; Rodrigues et al., 2004; Zheng et al., 2005).
For the first time, the present study determined the allelic frequency of the CHIT1 gene in a healthy Iranian population sample. A wide range of frequencies have been identified in other subject populations throughout the world. In two separate studies of this 24-bp duplication mutation in 40 and 231 healthy Korean individuals, allelic frequencies of 58% and 56.1% were reported, respectively, which supports the existence of a high mutation frequency in healthy populations (Lee et al., 2007; Woo et al., 2014). A high allelic frequency level for the 24-bp duplication in the CHIT1 gene was also reported in some East Asian countries, including Japan, Taiwan, and China (Chien et al., 2005; Lee et al., 2007). However, given the small number of subjects in these studies, larger sample sizes are required for a definitive assessment of allelic frequency in these populations with high accuracy. On the other hand, the allelic frequency for the 24-bp duplication in Caucasian (∼20%), Spanish (8-13%), and African (5-7%) populations is low, indicating that ethnic diversity is a factor in the duplication frequency (Grace et al., 2007; Lee et al., 2007). Therefore, the difference in CHIT1 allele frequency and enzyme activity in the Iranian population was comparable to other populations and might be attributed to ethnic diversity.
Since CHIT1 mediates natural immune responses against chitin-containing pathogens, such as plasmodium, mycobacterium, candida, and other parasites (Boot et al., 2001; Malaguarnera et al., 2003; Lehrnbecher et al., 2005; van Eijk et al., 2005), it was previously thought that the incidence of the 24-bp duplication in the CHIT1 gene may increase the susceptibility to these pathogens. However, subsequent studies refuted the presence of such a relationship (Masoud et al., 2002; Choi et al., 2005). Mutations in CHIT1 are now considered to be an evolutionary advantage in humans (Lee et al., 2007), and thus natural selection may have eliminated the enzyme activity.
In this study, the 24-bp duplication was evaluated by PCR-HRM in addition to the PCR and electrophoresis methods that were used in previous studies. HRM analysis is a relatively new genotyping technique that allows for the visualization of genotypic changes through displacements obtained from HRM curves. Samples from wild type as well as heterozygous and homozygous individuals could be differentiated by the significant differences in HRM curves. Based on accuracy, reliability, cost, and time, the HRM method appears to be the most suitable and powerful tool for genotype screening to date. Unlike other methods that scan for genetic changes, HRM does not require gel electrophoresis. Therefore, the advantage of the HRM method for assessing patient samples is that PCR tests and melting curve analysis processes are carried out in parallel in a closed system, which reduces the risk of product contamination and also saves time (Reed et al., 2007; Erali et al., 2008).
Although the CHIT1 activity is routinely used to monitor patients with Gaucher's disease treated with ERT, despite the presence of other useful secondary markers, questions remain regarding the accuracy of the method. In contrast, a secure genotyping method is valuable for interpreting results in the context of the genotype. It is also noteworthy that although we found that the CHIT1 activity was higher in wild-type homozygotes than heterozygotes, there is still an overlap between these values, which precludes genotype determination by biochemical analysis.
HRM analysis is a fast, reliable, highly accurate, and cost-effective screening method for identifying the 24-bp duplication in exon 10 of the CHIT1 gene. Moreover, due to the wide range in the frequency of this duplication among different ethnic groups, additional studies are needed to identify new biomarkers that take into account the genetic characteristics of different populations.
Footnotes
Acknowledgments
This study was financially supported by a research grant (Grant No. 92031115) from the Iran National Science Foundation. We gratefully acknowledge support from Tarbiat Modares University. We also thank the staff of the Pasteur Institute of Iran and Massoud Laboratory of Tehran, Iran.
Author Disclosure Statement
No competing financial interests exist.