
Abstract
Objective: To assess the relationship between the ataxia telangiectasia mutated (ATM) gene-111 (G>A) polymorphism and cancer risk. Methods: The PubMed, EMBASE, Web of Science, and China National Knowledge Infrastructure databases were searched comprehensively. A total of 16 case-control studies with 12,273 cases and 13,046 controls were included in this meta-analysis; 12 of them were from the Chinese population. Five studies assessed smoking effects, including 3038 smokers and 1003 nonsmokers. Odds ratio (OR) was determined by using a genetic model-free approach. Heterogeneity was quantified by I2 statistics. Publication bias was also evaluated. Results: The recessive model (AA vs. AG + GG) was suggested as the most appropriate genetic model. After elimination of heterogeneity, it was found that the ATM-111 (G>A) AA genotype is significantly associated with increased susceptibility to overall cancer risk in a fixed effects model (OR = 1.09; 95% CI = 1.03-1.15; p < 0.01; I2 < 0.01). In the subgroup analysis, the result of pooled analyses among the Chinese population revealed similar associations (OR = 1.12; 95% CI = 1.04-1.22; p < 0.01; I2 < 0.01). As for specific cancer analysis, an increase was correlated with lung cancer risk (OR = 1.12; 95% CI = 1.01-1.24; p = 0.03) and breast cancer risk (OR = 1.08; 95% CI = 1.00-1.16; p = 0.05). In addition, a stronger association was found among nonsmokers (OR = 1.31; 95% CI = 1.13-1.52; p < 0.01). Conclusion: This meta-analysis suggests that AA genotype of the ATM-111 gene (G>A) may be a risk factor for breast cancer and lung cancer, especially among nonsmokers, within the Chinese population.
Introduction
C
Ataxia telangiectasia mutated (ATM) gene is a DNA repair gene that has an important role in DNA repair, apoptosis, and cell cycle arrest, to maintain genomic integrity (Chaudhary and Al-Baradie, 2014). The protein it encodes is a key mediator of DNA double-strand breaks (DSBs). DSBs are among the most harmful type of DNA damages that could give rise to the development of cancer by impairing genomic stability and inducing deleterious mutations (Bennett et al., 1993; Aparicio et al., 2014). Once DSBs occur, ATM proteins are activated, which then phosphorylate P53, BRCA1, CHK2, and other proteins. It is followed by the activation of cell cycle checkpoints, DNA repair, and apoptosis (Shiloh, 2003; Chaudhary and Al-Baradie, 2014). In fact, ATM is commonly believed to be a tumor-suppressor gene (Rondeau et al., 2015). The downregulation of ATM expression has been correlated with carcinogenesis in a variety of cancers, including prostate cancer (Fan et al., 2006), renal cell carcinoma (Selvarajah et al., 2013), endometrial carcinoma, and more (Shan et al., 2015).
Single-nucleotide polymorphisms (SNPs) of ATM have been associated with cancer susceptibility (Shen et al., 2012). Among them, ATM-111 (G>A), a common functional SNP in the promoter region of the ATM gene, was reported to be a binding site of transcription factors that affect the ATM expression (Chen et al., 2010; Zhang et al., 2010). Early epidemiological studies have identified ATM-111 (G>A) polymorphism as a strong risk factor in lung, breast, and oral cancers (Bau et al., 2010; Lo et al., 2010; Hsia et al., 2013; Zhao et al., 2013; Liu et al., 2014). However, inconsistent results were reported in thyroid cancer, leukemia, and other cancers (Pharoah et al., 2007; Zhao et al., 2013; Damiola et al., 2014; Song et al., 2015). This was largely due to the limitation of ethnicity of the population and small sample size from individual studies. Meta-analysis is known for resolving controversial results by combining the inconsistent results to provide large-scale subjects. Therefore, a comprehensive meta-analysis was performed to clarify the association between ATM-111 (G>A) polymorphism and cancer susceptibility.
Methods
Publication search
The literature from the PubMed, EMBASE, Web of Science, and China National Knowledge Infrastructure (CNKI) was searched to get relevant available articles. The following search terms were used: (“Ataxia telangiectasia mutated” OR ATM OR-111 (G>A) OR “rs189037”) AND (polymorphism OR variation OR mutation) AND (cancer OR tumor OR neoplasm OR carcinoma). The search was restricted to human study only and the language was unlimited. The last search was performed on April 30, 2015. Eligible studies were retrieved, and their references were examined to find other relevant publications. Only published studies with full text accessibility were included.
Inclusion criteria
The inclusion criteria were as follows: (1) a case-control study evaluating the association between ATM-111 (G>A) and the human tumor risk; (2) the control group consistent with Hardy-Weinberg equilibrium (HWE); and (3) sufficient data for calculating the pooled odds ratios (ORs) for all genetic models.
Methodological quality appraisal
To assess the quality of the included case-control studies, we applied the quality appraisal criteria amended from previous meta-analyses (Gao et al., 2011; He et al., 2014) (shown in Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/gtmb). These scores were based on both traditional epidemiological considerations and genetic issues. Methodological quality was evaluated by two independent investigators (J. Liu and B. Lin). Disagreements were resolved by discussion. Scores ranged from 0 to 16. Articles with a score <9 were regarded as “low quality,” whereas those ≥9 were categorized as “high quality.”
Data extraction
Data were carefully extracted independently by two authors (SL.H and T). For conflicting data extraction, a consensus was reached by discussion. The authors of eligible studies were also contacted to get the additional information required for the pooled analysis. The extracted data are presented in Tables 1 and 2.
PB, population-based; HB, hospital-based; RFLP, restriction fragment length polymorphism; TaqMan, real-time TaqMan analysis; HWE, Hardy-Weinberg equilibrium; NPC, Nasopharyngeal carcinoma; TC, thyroid cancer.
HWE, Hardy-Weinberg equilibrium.
Statistical methods
The association between ATM-111 (G>A) polymorphism and cancer risk was assessed by the pooled ORs with 95% CIs. First, the pooled OR was calculated for SNP allele frequency to determine the risk allele (A allele vs. G allele, A was for the minor allele and G was for the major allele). Because of the lack of current knowledge regarding the genetic model of inheritance that might explain the effect of ATM-111 (G>A) on carcinogenesis, the most appropriate genetic model was adopted as described by Thakkinstian et al. (2005), instead of performing pooled ORs for all genetic models that could significantly increase the probability of false-positive report. Briefly, pooled OR1 (AA vs. GG), OR2 (AG vs. GG), and OR3 (AA vs. AG) were calculated in this study. If OR1 = OR2 ≠ 1 and OR3 = 1, a dominant model (AA + AG vs. GG) was suggested. If OR1 = OR3 ≠ 1 and OR2 = 1, a recessive model (AA vs. AG + GG) was suggested. If OR2 = 1/OR3 ≠ 1 and OR1 = 1, a complete overdominant model (AA + GG vs. AG) was suggested. If OR1 > OR2 > 1 and OR1 > OR3 > 1, or OR1 < OR2 < 1 and OR1 < OR3 < 1, a codominant (AA vs. AG, AA vs. GG) model was indicated.
Heterogeneity was checked by the chi-square-based Q test (p > 0.05 indicates a lack of heterogeneity) (Rongling and Li, 1999). A fixed effects model (the Mantel-Haenszel method) was performed when no heterogeneity existed (p < 0.10) (Mantel and Haenszel, 1959). Otherwise, the random effects model (DerSimonian and Laird method) was performed (DerSimonian and Laird, 1986). Heterogeneity was also quantified by I2 statistics (I2 < 25%, no heterogeneity; I2 = 25-50%, medium heterogeneity; I2 = 50-75%, high heterogeneity; I2 > 75%, extreme heterogeneity) (Higgins et al., 2003). The HWE analysis was performed for both case and control groups of each study, using the goodness-of-fit chi-square test. p > 0.05 indicated that the group was consistent with the HWE analysis. Subgroup analyses were performed by the type of cancer, ethnicity, control source, and smoking habits to find the source of heterogeneity. Sensitivity analysis was performed to assess the stability of the results, in which a single study was deleted each time to reflect the influence of individual data set to the pooled ORs. Publication bias was estimated by the funnel plot and Egger's test (p > 0.05 indicated no significant publication bias) (Begg and Mazumdar, 1994; Egger et al., 1997). All statistical analyses were performed with the STATA version 12.0 software.
Results
Extraction process and study characteristic
Based on the search strategy, 4414 articles were found, but only 74 of them were preliminarily identified for further evaluation. Finally, 20 case-control studies were discerned. However, three studies were excluded because of insufficient data (Barroso et al., 2009; Rendleman et al., 2014; Hayashi et al., 2015). Two studies used the same data, so only one of them was included (Pharoah et al., 2007; Yang et al., 2014). The publications of Liu et al. (2014) and Shen et al. (2014) are two separate case-control studies, but the source of nonsmoker female cases might have overlapped. After contacting with the authors, the nonsmoker female cases were removed from the study of Liu et al. Therefore, 16 articles were included in this meta-analysis with 11,556 cases and 12,522 controls (Fig. 1). The characteristics of these studies are summarized in Tables 1 and 2. Of all the 16 studies, 8 kinds of cancers were addressed, including 5 studies on lung cancer, 4 on thyroid cancer, and 2 on breast cancer. In view of ethnicity, 13 out of 16 studies came from the Asian population, including 12 from the Chinese population. There were a total of 11 population-based studies and 5 hospital-based studies. Restriction fragment length polymorphism genotyping methods were used in five studies and TaqMan methods in eight. All studies were consistent with HWE in controls. Only the study of Bau et al. deviated from HWE in cases. There were five studies discussing the interaction of smoking behaviors and ATM-111 (G>A). Of these five studies, three kinds of cancers were addressed, including three studies on lung cancer, one on oral cancer, and one on esophageal squamous cell carcinoma. As for the assessment of study quality, the results are shown in Supplementary Table S2. All 16 studies were categorized as “high quality.” Numbers of AA, AG, and GG carriers of cases and controls in each study were available for further pooled analysis.
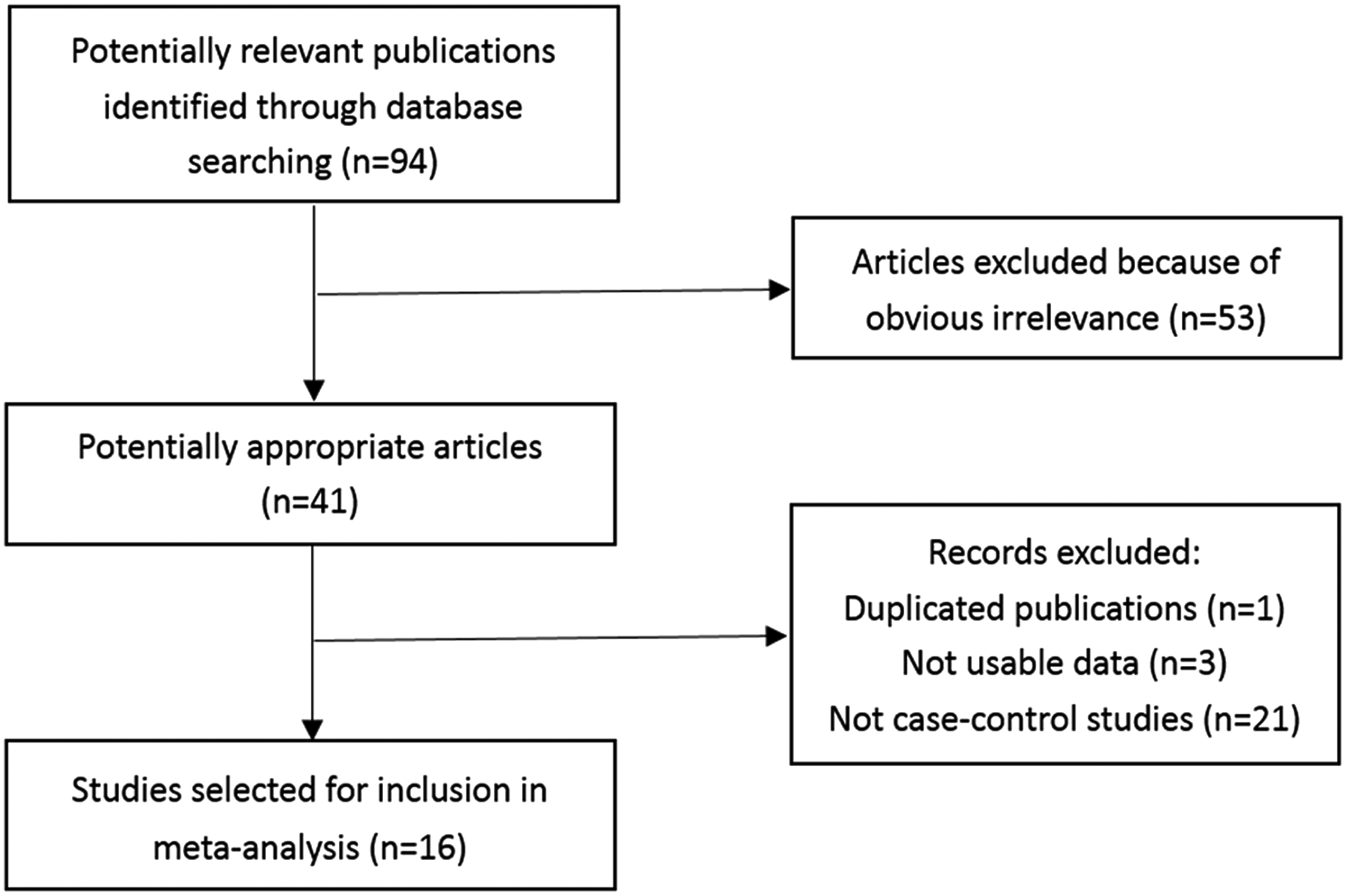
Identification, inclusion, and exclusion of studies in this meta-analysis.
Test of heterogeneity
Medium heterogeneity was detected in the comparison of overall cancer with 16 studies (I2 = 41.1%) and the subgroup analysis of the Chinese population (I2 = 43.0%). When Bau's study on oral cancer was removed, heterogeneity vanished (I2 < 0.01%) in both, the comparison of overall cancer and the subgroup analysis of Chinese studies (Table 3).
LC, lung cancer; BC, breast cancer.
Meta-analysis results
The results of meta-analysis for allele frequency comparison suggested A allele of ATM −111G/A was the risk allele (OR = 1.04; 95% CI = 1.02-1.06; p = 0.004). Further pairwise comparisons showed that OR1, OR2, and OR3 of ATM-111G/A were 1.09 (1.05-1.14), 1.03 (0.97-1.09), and 1.09 (1.04-1.15), respectively. Therefore, the recessive model (AA versus AG + GG) was adopted as the best genetic model for susceptible A allele (OR1 = OR3 ≠ 1 and OR2 = 1).
Sensitivity analysis suggested that Bau's study was the possible cause of heterogeneity. After excluding Bau's study, it was found that ATM-111 (G>A) AA genotype significantly increased the susceptibility to overall cancer in a fixed model with no between-study heterogeneity (OR = 1.09; 95% CI = 1.03-1.15; p < 0.01; I2 < 0.01%; Fig. 2 and Table 3). Subgroup analysis was first performed in view of cancer types. An increase was significantly noted with lung cancer susceptibility (OR = 1.12; 95% CI = 1.01-1.24; p = 0.03) and breast cancer susceptibility (OR = 1.08; 95% CI = 1.00-1.16; p = 0.05), but not with thyroid cancer (OR = 1.09; 95% CI = 0.97-1.22; p = 0.18; Fig. 3 and Table 3). As for ethnicity, only the polymorphism among Chinese population was studied because of the limited number of studies among other ethnicities. The result of pooled analysis among the Chinese population revealed that the relation still existed (OR = 1.12; 95% CI = 1.04-1.22; p < 0.01; Fig. 2 and Table 3). In light of the control source, similar results were obtained in both population-based and hospital-based studies; ORs were 1.18 (1.03-1.35) and 1.12 (1.02-1.24), respectively. In addition, several studies evaluated the interaction of genetic variants of ATM-111 (G>A) and smoking habits. To explore the potential influence of smoking behavior, a pooled analysis was performed in this study in smokers and nonsmokers. A strong association between the ATM-111 (G>A) AA genotype and susceptibility to cancer was found in the nonsmokers (OR = 1.31; 95% CI = 1.13-1.52; p < 0.01; Fig. 4 and Table 3). However, in the smoker population, pooled analysis could not be performed because of the limited number of studies and high heterogeneity (I2 = 71.9%).
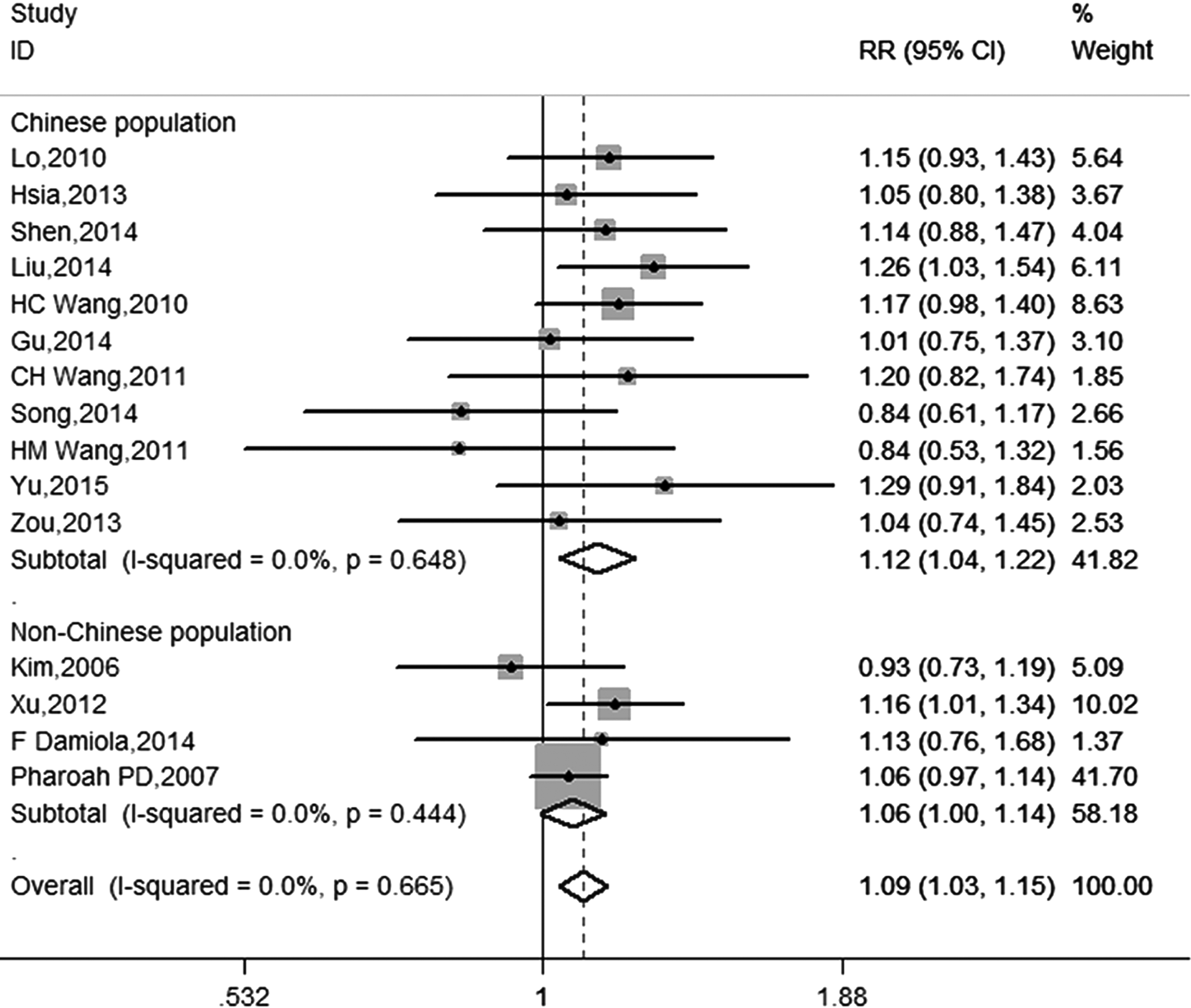
Forest plot of ATM-111 (G>A) polymorphism and cancer risk in the overall population after excluding elimination of heterogeneity (AA vs. GG + GA).
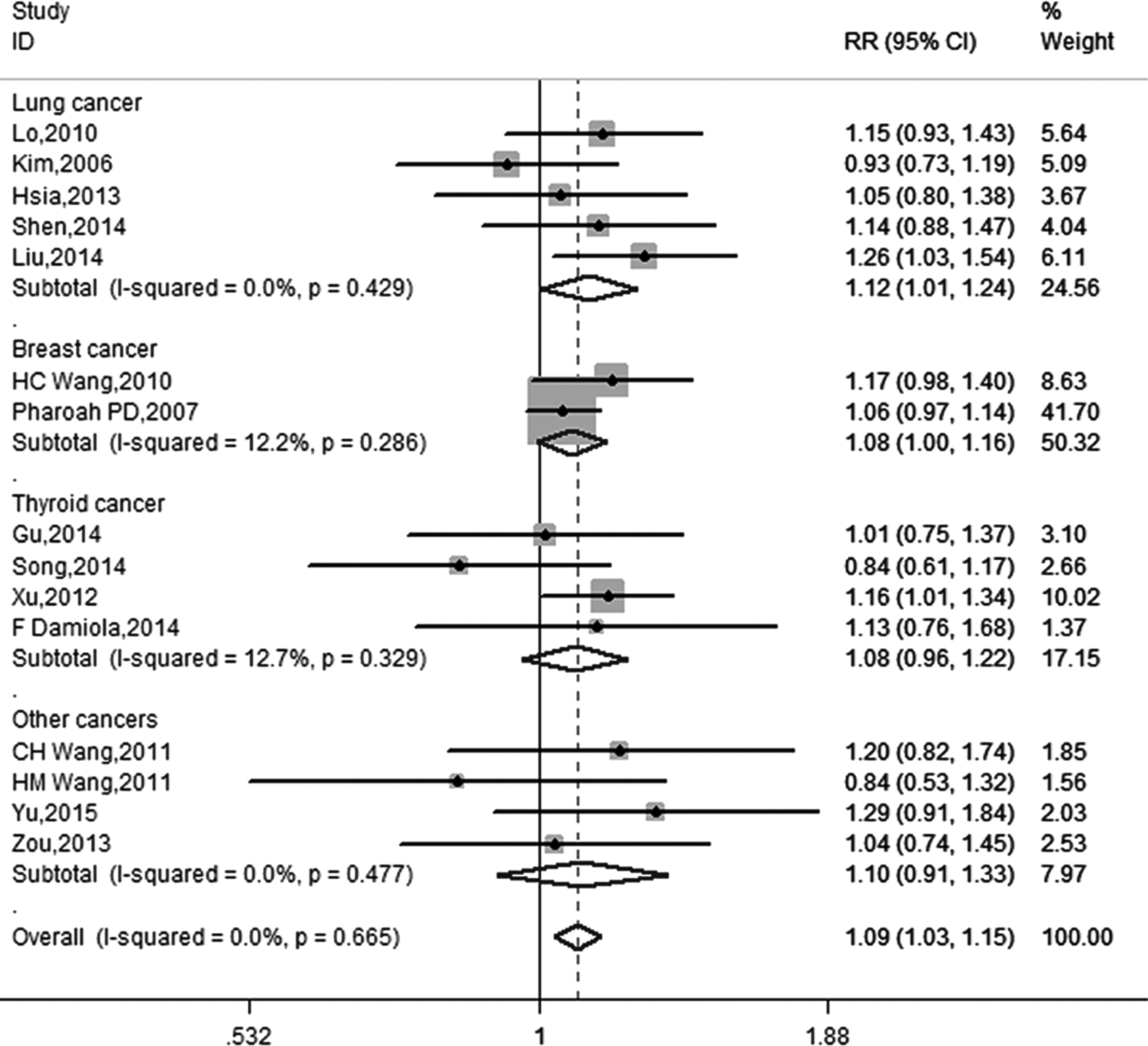
Forest plot of ATM-111 (G>A) polymorphism and cancer risk observed in subgroup analyses by cancer type after excluding elimination of heterogeneity (AA vs. GG + GA).

Forest plot of ATM-111 (G>A) polymorphism and the risk of nonsmoker cancers after excluding elimination of heterogeneity (AA vs. GG + GA).
Sensitivity analysis
Sensitivity analysis was performed by the removal of a single study each time. No individual study had a modifying influence on the pooled ORs, indicating the stability of the present study results (Fig. 5).

Sensitivity analysis of ATM-111 (G>A) polymorphism and cancer risk in overall population after excluding elimination of heterogeneity (AA vs. GG + GA).
Publication bias
Both funnel plot and Egger's test were used to examine the publication bias. After excluding Bau's study, the shape of the funnel plot did not reveal any obvious asymmetry for overall comparison (Fig. 6). Then Egger's test was performed to give the statistical evidence of funnel plot symmetry (p = 0.89). The results suggested that no publication bias existed.

Funnel plot analysis to detect the publication bias between ATM-111 (G>A) polymorphism and cancer risk in the overall population, excluding elimination of heterogeneity (AA vs. GG + GA).
Discussion
This study, perhaps, was the first comprehensive meta-analysis exploring the association between polymorphism of ATM-111 (G>A) and overall cancer risk. The results suggested that carrying at least one A allele, especially ATM-111 (G>A) AA genotype, significantly increased the susceptibility to overall cancer, which is similar to the conclusion of individual case-control studies (Bau et al., 2010; Wang et al., 2011; Liu et al., 2014).
In the following cancer-specific analysis, a significant association was found for lung cancer risk and breast cancer risk. However, there was no evidence of ATM-111 (G>A) polymorphism on susceptibility to thyroid cancer. These features suggested different roles of ATM-111 (G>A) in different cancers. Another possibility of inconsistent results might be that small sample size prevented the studies from detecting small but existing association. In the current study, only four studies with limited sample size were available for thyroid cancer. Therefore, the result might be capricious. Larger, well-designed studies are needed to explore the exact role of ATM-111 (G>A) polymorphism in different types of cancer.
Considering smoking behaviors might affect cancer risk and result in bias, a pooled analysis among nonsmokers was also performed in this study. A remarkable risk was indicated in the nonsmoker subgroup, which is in agreement with major previous studies (Bau et al., 2010; Wang et al., 2011; Liu et al., 2014; Shen et al., 2014). Shen's study also reported that increased risk was associated with ATM-111 (G>A) AA genotype and lung adenocarcinoma in nonsmokers without exposure to cooking oil fumes. In light of the lack of obvious exposure to known carcinogens, the genetic effect may play a more dominant role in never smokers with lung cancer. However, high heterogeneity was shown in the smoker subgroup. So the pooled analysis in smokers was not performed in this study. High heterogeneity may come from the interaction of ATM-111 (G>A) polymorphism and differing intensity of environmental factors. The study among atomic bomb survivors described that survivors with ATM-111 (G>A) AA genotype who were exposed to >530-mGy dose level radiation revealed the highest breast cancer risk (relative risk, RR = 4.76). However, for <530-mGy radiation dose levels, survivors with GG genotype showed a higher breast cancer risk (RR = 1.85) (Hayashi et al., 2015). In this meta-analysis, only four studies were available for smokers and none of them tried to control the smoking dosage, which could be a possible source of heterogeneity. Further researches are required to clarify the interaction by controlling the smoking dosage.
Heterogeneity among studies is an important index for determining the quality of meta-analysis. Unfortunately, medium heterogeneity was found in the comparison of overall cancer and in the subgroup analysis of the Asian population-based studies. Bau's study on oral cancer played a significantly modifying role on heterogeneity by sensitivity analysis. In addition, this study was the only “off-group point” in the funnel plot (data not shown). Interestingly, this study is the only study where obvious deviation from HWE was detected in cases. Departure from HWE in cases often indicates an additional support for genetic association. Other possibilities include chance, genotyping error, sampling bias, and population stratification (Zintzaras and Lau, 2008). Considering its OR value was the highest among available studies (OR = 1.93), whether ATM-111 (G>A) polymorphism was more dominant in oral cancer requires larger, well-designed studies.
Some limitations of this meta-analysis should be mentioned. First, the number of included studies was still not sufficient enough, particularly for ethnicity subgroup analysis and the single type of cancer. Second, no adequate data were offered so the potential interaction was not further estimated such as gene-gene interaction and gene-environment interaction. Third, only published studies were retrieved so inevitable publication bias may have existed, although Begger's funnel plot and Egger's test indicated no publication bias.
In conclusion, this meta-analysis suggested that ATM-111 (G>A) polymorphism is significantly associated with susceptibility to cancer, especially for Asians and nonsmokers. Large sample and well-designed studies are needed to confirm the findings and determine the role of gene-environment interaction.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of PR China (no. 81201831), Guangdong Planning Project of Science and Technology (no. 2014A020212190), Social Science and Technology Development Program of Dongguan (no. 2013108101061), and Dongguan Science and Technology Planning Project of Medical Treatment and Public Health (no. 20131051010005) to TZ. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
The authors declare no competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.