
Abstract
Aims:
The aim of this study was to evaluate the expression of OLR1 and its alternative splicing isoform Loxin in unexplained recurrent miscarriage (uRM).
Methods:
Sixty-three women of reproductive age were recruited and were divided into four groups: 18 pregnant and 23 non-pregnant women with uRM, and 12 pregnant and 10 non-pregnant women with physiological pregnancies. Complementary DNA derived from peripheral blood mononuclear cells (PBMCs) was analyzed by quantitative real-time PCR to evaluate the expression of OLR1 and Loxin. Oxidized low-density lipoproteins (ox-LDLs) were assayed from serum by a commercially available kit.
Results:
Pregnant uRM women presented with a higher, though not significant, OLR1/Loxin ratio and a higher ox-LDLs serum level (p ≤ 0.05) compared with pregnant control women. OLR1 and Loxin levels were significantly decreased in non-pregnant uRM women compared with the control (OLR1: 0.00018 vs. 0.00043, p ≤ 0.005; Loxin: 0.00018 vs. 0.00060, p ≤ 0.005, respectively). Loxin expression decreased by about two-thirds (p ≤ 0.005) in pregnant women compared with non-pregnant control women. A higher expression of OLR1 in pregnant women compared with non-pregnant women with uRM (p ≤ 0.05) was observed, but no variation in Loxin expression was observed.
Conclusions:
The results of this study show an association of peripheral OLR1 and Loxin expression levels in uRM women, and they suggest the possible existence of an uncontrolled oxidative stress in these women in the first trimester of pregnancy.
Introduction
M
Although some of the causes of RM are well known, about 50-60% of these events are still unexplained RM (uRM), and in these cases it has been suggested that an aberrant immune response or an enhanced oxidative stress (OS) could be involved (Christiansen et al., 2008).
OS at the placental interface has been involved in the pathogenesis of pregnancy complications such as abortion, pre-term labor and birth, fetal growth restriction, and preeclampsia (Hubel, 1999; Vural, 2000; Burton and Jauniaux, 2004; Duhig et al., 2016; Jauniaux and Burton, 2016). Recent data on animal models clearly demonstrate that OS leads to spontaneous abortion and RM; in fact, OS causes placental inflammation and embryonic developmental arrest, resulting in fetal loss under hypoxic conditions (Ishii et al., 2014).
Literature data indicate that systemic and placental OS may have a potential key role in the etiology of RM (Vural, 2000; Gupta et al., 2007; Ghneim and Alshebly, 2016). Indeed, an increased production of active oxygen species in the granulocytes of patients with a history of RM compared with women with normal reproductive function was found (Safronova et al., 2003). Moreover, a recent work reported a significant increase of total oxidant level and OS index together with a significant decrease of total antioxidant capacity in pregnant women with a history of RM (Yiyenoğlu et al., 2014).
Oxidized low-density lipoproteins (ox-LDLs) constitute one of the principal studied OS biomarkers. Elevated levels of ox-LDLs and lower antioxidant defenses in women affected by preeclampsia or RM suggest that an uncontrolled lipid oxidation or a reduction in ox-LDLs turnover can lead to endothelial dysfunctions and to placental injuries (Branch et al., 1994; Tulppala et al., 1995; Zhang et al., 2015). Interestingly, women with RM have been found to have higher rates of endothelial dysfunction compared with women who had physiological pregnancies (Germain et al., 2007; Banerjee et al., 2013; Pasquier et al., 2013). Moreover, ox-LDLs can inhibit human extravillous trophoblast invasion in a dose-dependent way (Pavan et al., 2004).
Accordingly, a controlled uptake and degradation of modified LDLs could prevent their toxic effect on placental cellular function as well as on the growth and development of the fetus (Bonet et al., 1995).
The most important endothelial receptor for the ox-LDLs is the lectin-like ox-LDL receptor-1 (LOX-1), which can bind, internalize, and degrade ox-LDLs in response to different stimuli, such as angiotensin II, tumor necrosis factor alpha, advanced glycation end products, and ox-LDLs themselves (Moriwaki et al., 1998; Aoyama et al., 1999; Li et al., 2000; Chen et al., 2001; Yoshimoto et al., 2011; Zeya et al., 2016). LOX-1 is highly expressed in vascularized tissues such as placenta, lungs, bone marrow, and spinal cord (Yamanaka et al., 1998).
The most elevated LOX-1 messenger RNA (mRNA) expression is in the placenta, and this prompted its potential crucial role in placental function, especially during the earliest stages of pregnancy (Moriwaki et al., 1998). In fact, at the end of the first trimester, placental OS increases quickly and placental vascular development occurs to facilitate the embryo implantation through an immunological modulated response (Jauniaux et al., 2006). LOX-1 is also involved in trophoblast invasion during the early stages of pregnancy (Lee et al., 2005; Fournier et al., 2007; Zuniga et al., 2014), in the trophoblast accelerated apoptosis process (Satoh et al., 2008; Zuniga et al., 2014), in endothelial dysfunction (Zuniga et al., 2014), and in regulating trophoblast angiogenic pathways in preeclampsia (Zhang et al., 2015).
LOX-1 is also expressed on the surface of immune cells, such as dendritic cells and macrophages. This wide pattern of LOX-1 expression suggested that it might have additional immune functions (Joo et al., 2014).
Moreover, LOX-1 upregulation induces the activation of proinflammatory process, through nuclear factor-κB (NF-kB) signaling (Robbesyn et al., 2004), that leads to the expression of different cytokines, such as interleukin-6 (IL-6) and necrosis factor alpha (TNF-α) (Hashizume and Mihara, 2012), the activation of mitogen-activated protein kinase (MAPK) proinflammatory cascade, and the decrease of the anti-inflammatory IL-10 (Mehta et al., 2007).
Recently, LOX-1 was found to trigger NLRP3 inflammasome, a “molecular platform for immune defense,” which implies the production of the proinflammatory IL-1 family of cytokines, such as IL-1b and IL-18 (Ding et al., 2014; Yang et al., 2016).
Interestingly, RM is characterized by an abnormal immune response, which includes high TNF-α, interferon alpha (INF-α), and IL-6 levels, an increase in the proinflammatory response of IL17+ T cells, a decrease of Foxp3+ T regulatory cells, an increase of TNF-a/IL10-producing T cell ratio (Christiansen, 2013), and an increased endometrial expression of NALP-3 (D'Ippolito et al., 2016).
Furthermore, it has pointed out the hypothesis that an increased cellular immunity and perhaps a compensative OS could be considered an etiological factor for RM, showing a link between lipid peroxidation and activated T cell function (Daglar et al., 2016). Even more, the immunoregulatory role of OS in the early first-trimester hypoxic condition on uterine natural killer (uNK) cell development and maturity, T cell cytotoxicity suppression, and enhancement of Treg function were guessed (Wu et al., 2016).
All these data make OLR1 a candidate gene with a key role in the pro-inflammatory processes that are characteristic of the RM pathogenesis.
The LOX-1 receptor is encoded by the OLR1 gene (NM_002543), which is located on human chromosome 12 (Sawamura et al., 1997). OLR1 gene is subjected to alternative splicing that generates two mRNA isoforms called, respectively, Loxin (NM_001172633) and OLR1D4 (NM_001172632). Loxin, the best-known OLR1 isoform, lacks exon 5 and this leads to the synthesis of a non-functional receptor (LOXIN). LOXIN is unable to bind and internalize ox-LDLs due to the missing of two-thirds of the extracellular lectin-like binding domain (Mango et al., 2005). Studies on murine embryos show the existence of functional murine OLR1 splicing isoforms that are specifically expressed during embryogenesis, suggesting a putative different role of these isoforms during pregnancy (Vecchione et al., 2012).
No data are available on OLR1 and its splice isoform expression in women affected by RM and, in particular, by unexplained RM.
This pilot study was designed with the aim of characterizing expression patterns of OLR1 and Loxin in uRM with a diagnostic and prognostic value.
Materials and Methods
Subjects
This study involved 63 women of reproductive age who were divided into four groups: (1) Eighteen pregnant women with uRM (p-uRM), (2) twenty-three non-pregnant women with uRM (np-uRM), (3) twelve pregnant women with physiological pregnancies (pregnant control group, p-CTR), and (4) ten non-pregnant women with physiological pregnancies (non-pregnant control group, np-CTR).
All the women included in this study attended as outpatients the Complex Operative Unit of Gynecology and Obstetrics at the University Hospital Policlinico Tor Vergata of Rome. Women with RM were followed at the RM Outpatient Clinic, whereas controls were followed at the General Gynecology and Physiological Pregnancy Outpatient Clinics.
Women with RM underwent a standardized diagnostic workup (Masini et al., 2009; Ticconi et al., 2011, 2016; Pietropolli et al., 2015), which included the following: (a) collection of familial and personal medical, gynecological, and obstetrical history with specific application to the previous miscarriages; (b) gynecological examination; (c) transvaginal ultrasound; (d) hysteroscopy and endometrial biopsy; (e) endocrine evaluation panel: assay of luteinizing hormone, follicle-stimulating hormone, prolactin, progesterone in the midluteal phase, thyroid-stimulating hormone, free triiodothyronine, free thyroxine, pituitary and ovarian androgens, and insulin and glucose curve; (f) karyotype of both partners; (g) immunity panel: anti-phospholipid antibodies, lupus anticoagulant, anti-β2GPI and anti-annexin V antibodies, anti-thyroid antibodies, anti-nuclear antibodies, extractable nuclear antigen, anti-double stranded DNA, anti-smooth muscle, and anti-mitochondrial antibodies; (h) thrombophilia screening: protein C, protein S, AT III, activated protein C resistance (with factor V-depleted plasma [APCR]), and homocysteine; and (i) determination of the following mutations: factor V [G1691A Leiden], factor II prothrombin [G20210 A], plasminogen activator inhibitor [PAI-1 4G/5G], and methenyl tetrahydrofolate reductase [MTHFR C677T and A1298C].
This workup was aimed to identify proven, probable, and doubtful causes of RM (Christiansen et al., 2008). When all the known causes for pregnancy loss mentioned earlier had been discarded, women were diagnosed with uRM and were included in this study.
All pregnant women affected by RM were treated with low-molecular-weight heparin (LMWH) (Clexane® 4000) from the first positive serum β-human chorionic gonadotropin (β-hCG) assay.
Control groups included non-pregnant (np-CTR) and pregnant (p-CTR) women with a history of least two normal pregnancies at term, without any miscarriage. These women were followed according to standard protocols for normal gynecologic surveillance or for physiological pregnancy.
Pregnant women with uRM and control pregnant women were enrolled within 7w + 0d/13w + 6d of gestational age. Blood samples (10 mL) were obtained only after written informed consent had been provided.
This study was approved by the Bioethical Committee of Policlinico Tor Vergata (139/12).
PBMC and plasma isolation
Peripheral blood mononuclear cells (PBMCs) and plasma have been isolated by whole blood (10 mL) by using Ficoll® Paque Plus (GE Healthcare, Little Chalfont, UK) according to the manufacturer's instructions. Isolated PBMC, resuspended in 1 mL of Trizol® (Ambion, Waltham, MA), and plasma samples were stored at −80°C until further analysis.
RNA extraction and retrotranscription
RNA extraction was performed from isolated PBMCs according to Trizol protocol. RNA concentration was evaluated by using a NanoDrop ND-1000 Spectrophotometer (EuroClone), whereas RNA quality was checked on agarose gel 1%. Next, 1.5 μg of total RNA was retrotranscribed by using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA).
Quantitative real-time PCR
A quantitative real-time PCR (SYBR Green Assay; Applied Biosystems) assay was performed by using an ABI 7500 Fast Real-Time PCR System (Applied Biosystems) to verify the expression level of each OLR1 isoform (OLR1 NM_002543; Loxin NM_001172633) and by using specific primer pairs designed with the software Primer Blast. RPL37A (NM_000998.4) and β-ACTIN (NM_001101.3) genes were used for data normalization (Table 1). Each analysis was performed in triplicate at least twice (n = 6). To evaluate the expression level of each OLR1 isoform, the threshold cycle (Ct) values of each sample were extracted from the Real-Time PCR 7500 Software (Applied Biosystems).
mRNA, messenger RNA.
The Ct values express a relative measure of the concentration of a target in the PCR reaction. The normalization of the Ct values with the selected housekeeping genes (RPL37A and β-ACTIN) allows one to calculate the 2−ΔCt value for every sample. The 2−ΔCt value represents the absolute expression level of the target gene in the sample, and it is shown in the graphs.
Biochemical analysis
Routine biochemical parameters such as apolipoprotein A1 (ApoA1) and apolipoprotein B100 (ApoB100) were measured on the Siemens Dimension Vista® 1500 System (Siemens, Inc., South San Francisco, CA). Quantitative measurement of ox-LDLs in human serum was performed by using an immunoassay kit (Mercodia AB, Uppsala, Sweden).
The assay characteristics (intra-assay precision, inter-assay precision, and detection limit) are provided by the kit manufacturer.
Statistical analysis
Kolmogorov-Smirnov test was used to analyze the distribution of the data. Data are expressed as mean ± standard deviation or as median and range, as appropriate. Women's clinical characteristics were analyzed by the Student's t-test. Statistical analysis for normally distributed data was carried out by using one-way analysis of variance followed by Tukey's test as post hoc multiple comparisons among groups. For data not normally distributed, Kruskal-Wallis followed by Dunn's test for multiple comparisons were used; for comparisons between two groups, Mann-Whitney test was used. Significance was set at p ≤ 0.05. Statistical analyses were performed by using GraphPad Prism version 6.0 (GraphPad Software, San Diego, CA).
Results
Clinical characteristics of studied women
The clinical characteristics of studied women are reported in Table 2. No significant differences were detected, except for age between p-CTR and p-uRM women (Student's t-test, p = 0.02). All unexplained RM patients who were enrolled in this study were negative to the standardized RM diagnostic workup, which is previously described.
Student's t-test, p < 0.05.
BMI, body mass index; np-CTR, non-pregnant control; np-uRM, non-pregnant unexplained recurrent miscarriage; n.s., not significant; p-CTR, pregnant-CTR; p-uRM, pregnant-uRM.
All uRM and control pregnant women successfully carried the pregnancy to term.
Expression level of OLR1 and Loxin isoforms in uRM
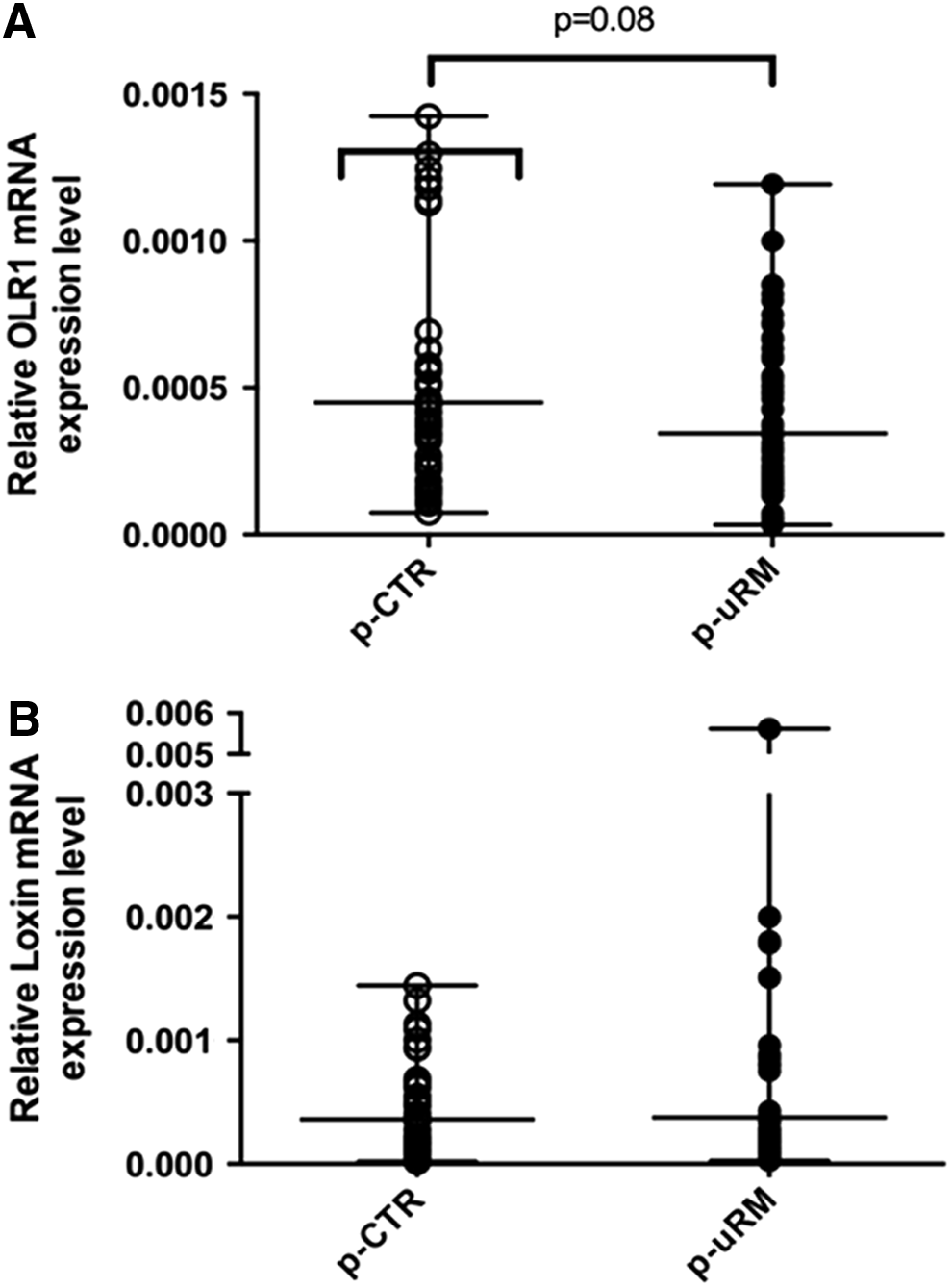
In a preconceptional period, uRM women showed a lower expression level of both OLR1 and Loxin isoforms (Fig. 1A, B). In particular, OLR1 expression is 50% lower in the uRM versus the CTR group (p ≤ 0.005) (Fig. 1A), whereas Loxin expression is decreased by 66% (p ≤ 0.005) (Fig. 1B). In the first trimester of pregnancy, however, the expression of OLR1 and Loxin did not change between women with physiological pregnancies and women with uRM (Fig. 2A, B), even though a not statistically significant (p = 0.08) lower OLR1 expression has been observed in uRM (Fig. 2A). In term of OLR1/Loxin ratio, both in a preconceptional period and during the first trimester of pregnancy, uRM women showed a higher OLR1/Loxin ratio compared with CTR women, even though it was not statistically significant (Fig. 3).
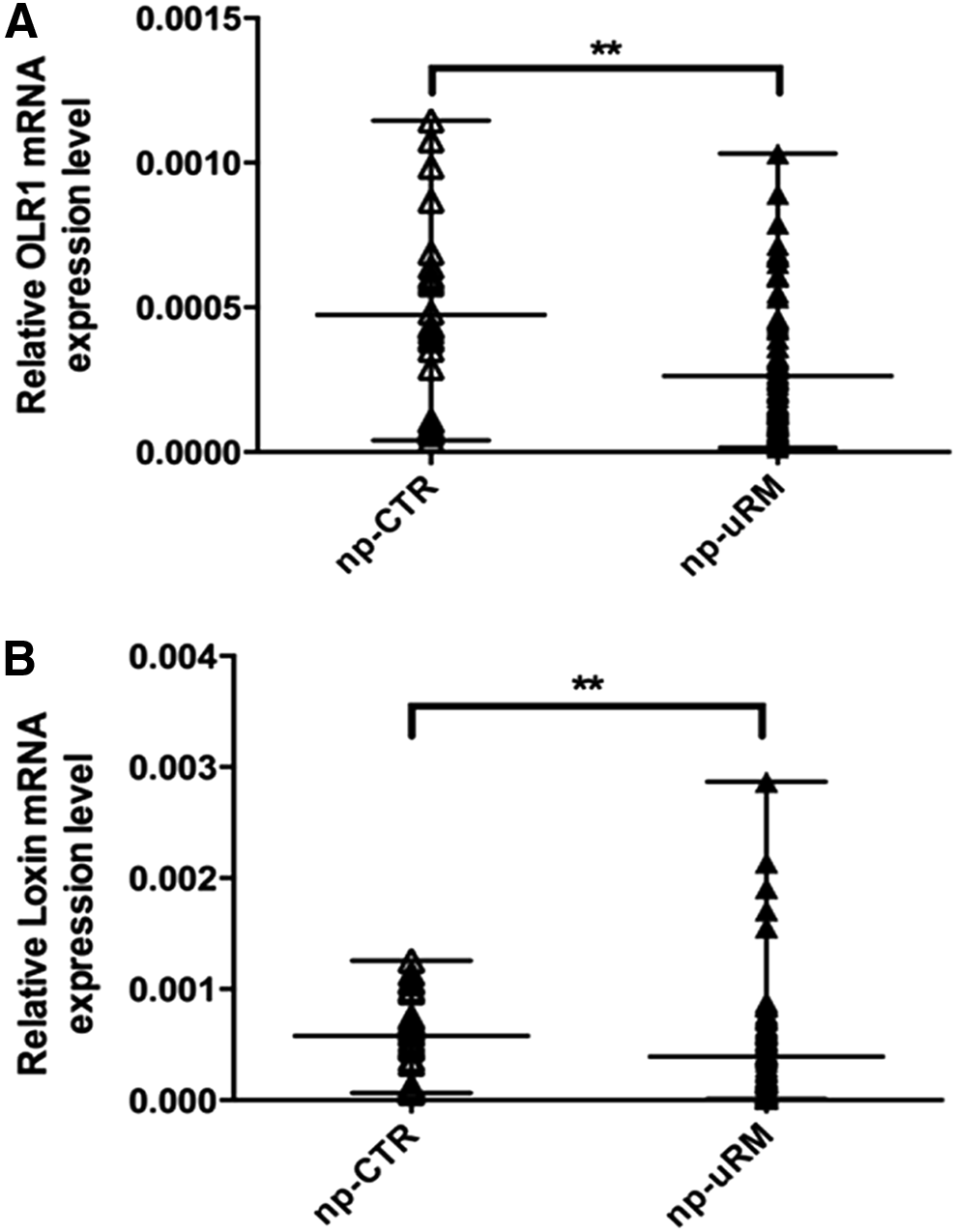
OLR1
OLR1

OLR1/Loxin messenger RNA ratio in np-CTR, np-uRM, p-CTR, and p-uRM groups. Kruskal-Wallis statistic 4.143, ns. Data are expressed as median and range.
Interestingly, if we compare the peripheral expression level of OLR1 isoforms during physiological pregnancies, we observed that OLR1 expression did not vary (Fig. 4A) whereas Loxin expression decreased by about two-thirds (p ≤ 0.005) (Fig. 4C). Conversely, during uRM pregnancies, we observed a higher expression of OLR1 (p ≤ 0.05) (Fig. 4B), but no variation in Loxin expression (Fig. 4D).
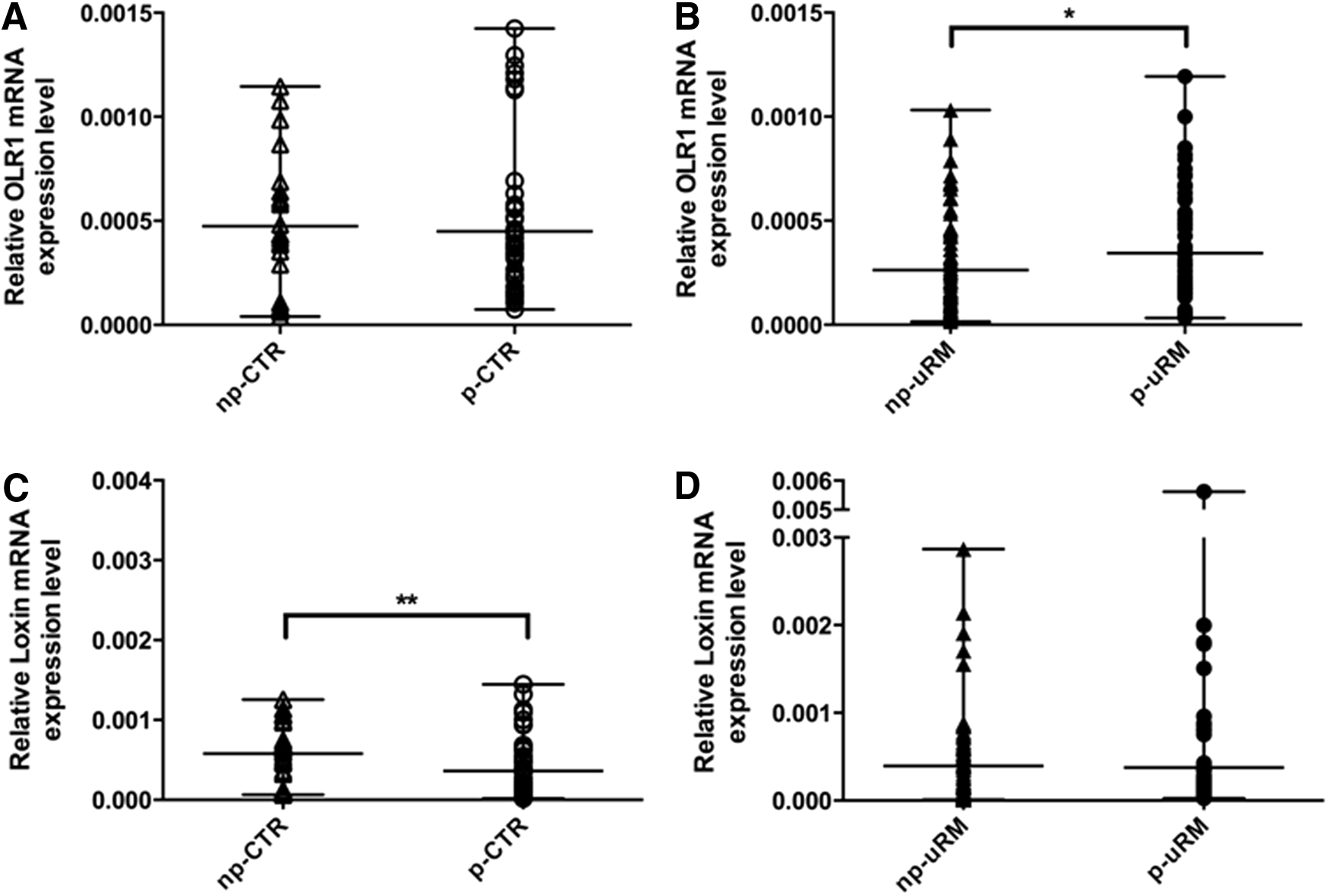
OLR1
Biochemical serum analysis
Since ox-LDLs are the principal ligands of the LOX-1 receptor, the ox-LDLs concentration in the sera of control and uRM women was measured.
A higher level of circulating ox-LDLs (p ≤ 0.05) in the sera of pregnant women with uRM (p-uRM) was found, compared with the sera of women with physiological pregnancies (p-CTR) (Fig. 5).
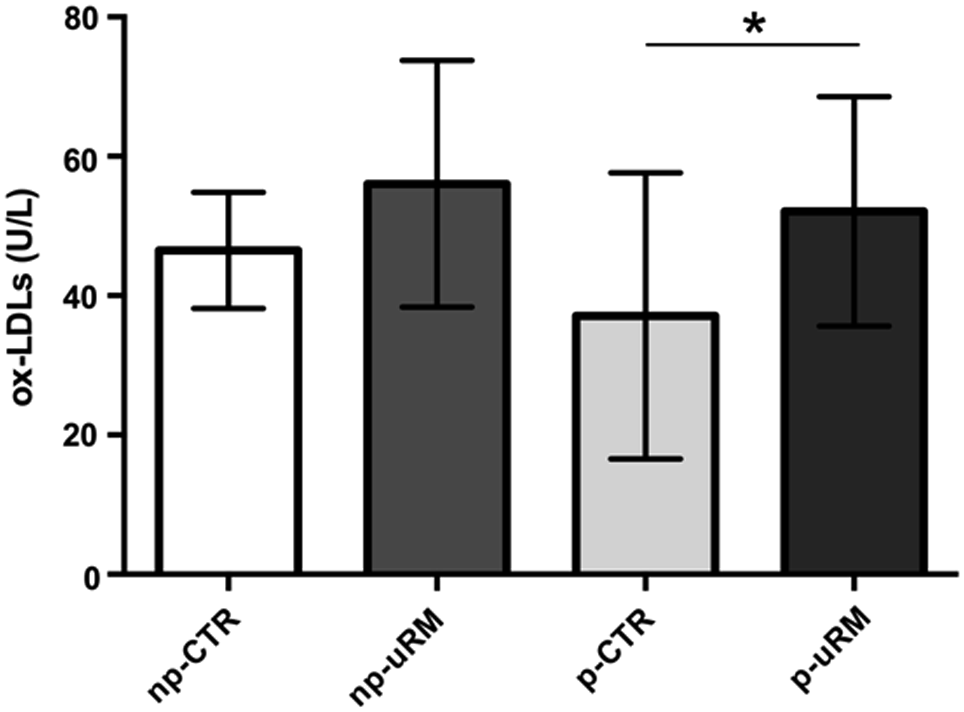
Serum concentration of ox-LDLs in np-CTR, np-uRM, p-CTR, and p-uRM groups. Data are expressed as mean ± standard deviation. One-way analysis of variance, F: 3.576, *Tukey's test p ≤ 0.05. ox-LDLs, oxidized low-density lipoproteins.
The levels of ApoA1 and ApoB100 were investigated. However, no differences in these levels among the groups were observed (data not shown).
Discussion
There is evidence that in pregnancy, especially during the first trimester, a controlled enhancement of the OS process occurs to facilitate the embryo implantation, through an immunological modulated response. In this context, it is likely that the expression of LOX-1 is higher during this gestational period (Moriwaki et al., 1998; Jauniaux et al., 2000). Indeed, it is during this gestation period that the higher incidence of miscarriages can be found in women suffering from RM (Ticconi et al., 2015). Accordingly, in this pilot study, pregnant women with uRM and control pregnant women within 7w + 0d-13w + 6d of gestational age were enrolled.
Our results show that during the preconceptional period, uRM women express significantly lower peripheral levels of both OLR1 and Loxin isoforms compared with control women (Fig. 1). In the first trimester of uRM pregnancy, however, the peripheral OLR1 expression significantly increases (Fig. 4B). Interestingly, during a physiological pregnancy, the peripheral expression level of OLR1 does not change, but the expression of the less active isoform, Loxin, decreases (Fig. 4C). In fact, LOXIN protein, encoded by Loxin isoform, forms inactive dimers with LOX-1 that are not able to bind and internalize ox-LDLs (Biocca et al., 2008). Therefore, a reduction of peripheral Loxin mRNA expression, in the first trimester of a physiological pregnancy, may mirror a mechanism that allows a higher presence of the active LOX-1 receptor on the cellular surface to regulate the OS, making it potentially useful for a normal placentation and prosecution of the pregnancy. On the contrary, an uncontrolled over-expression of OLR1 (encoding for the functional receptor LOX-1), as observed in uRM pregnant women (Fig. 4B), could lead to endothelial dysfunction (such as in atherosclerosis) and placental injury that, in turn, may be responsible for miscarriage. In addition, uRM women show an increase of OLR1/Loxin ratio (although not significant) both during the first trimester of pregnancy and in a preconceptional period when compared with the controls (Fig. 3).
A specific and balanced expression of the two isoforms of the OLR1 gene, OLR1 and Loxin, may have an important physiological role, that is, to guarantee a balanced OS, in physiological pregnancies and this may be missing in RM pregnancies. Little is known on the regulation of OLR1 and Loxin splicing. A very recent article demonstrated that a single-nucleotide polymorphism (SNP) in intron 4 (rs3736234) of OLR1 gene as the main contributors to Loxin expression, influencing the differential recognition of regulatory motifs by SR protein family members (SRSF1, SRSF2) and HMGA1 splicing factors (Tejedor et al., 2015). We have analyzed the distribution of this polymorphism in our population, but due to the low number of women (63 in total) participating to this study, we did not find any significant correlation between SNP rs3736234 and uRM (data not shown).
Interestingly, a recent study on Turkish women affected by gestational diabetes mellitus associated two known OLR1 SNPs (3′-UTR188C/T and K167N) to a higher OS and to a major presence of diabetic and cardiovascular risk factors. This evidence supports the hypothesis of an involvement of the OLR1 gene in obstetrical pathologies that are characterized by metabolic disorders, and it also suggests an involvement of the allelic configuration of OLR1 polymorphisms that deserves further investigation (Aydemir et al., 2015).
To the best of our knowledge, this is the first study suggesting that a different regulation of mRNA isoform expression, probably driven by an enhanced OS, seems to be played out at the beginning of physiological and RM pregnancies.
In this scenario, RM women who present a lower concentration of OLR1 and Loxin in a preconceptional period may be predisposed to an inappropriate and uncontrolled response to the physiological increase of OS during the first trimester; this, in turn, could have a preconceptional negative role in RM pregnancy, such as a not proper endometrial or immunological preparation to the pregnancy or an adverse environment for the continuation of the pregnancy (Fig. 6). Conversely, in women with physiological pregnancies, the balanced expression of OLR1 and Loxin can lead to a normal regulation of ox-LDLs concentration and, consequently, to a normal placentation during the first trimester of pregnancy (Fig. 6).
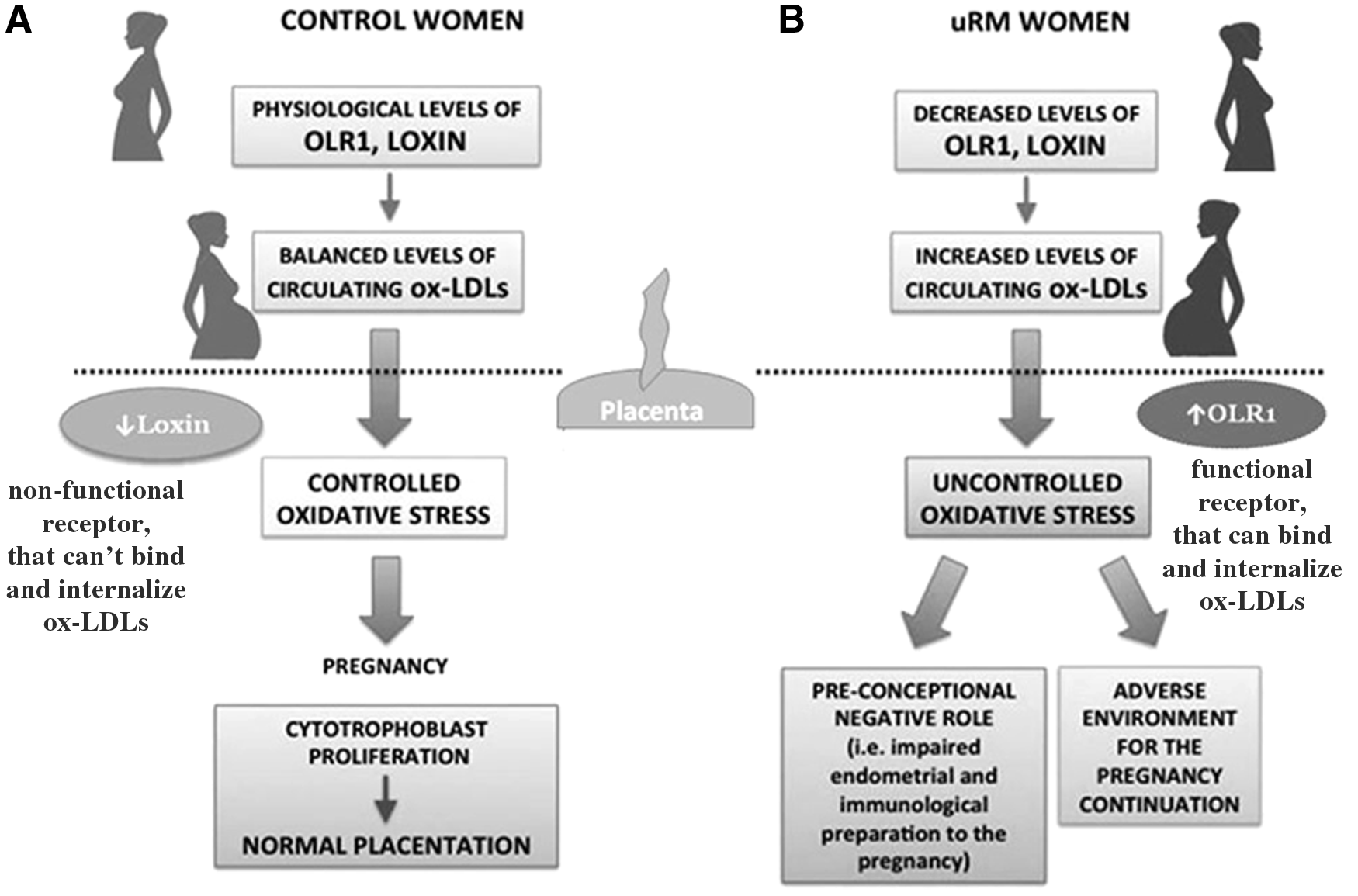
Potential LOX-1 pathophysiological mechanism of action in physiological and recurrent miscarriage pregnancies.
The adverse preconceptional role linked to decreased OLR1 and Loxin expression levels could be also correlated to the potential role of LOX-1 in proinflammatory process underlining the RM pathogenesis via MAPK and NF-kB activation or the NLP3 inflammasome (Joo et al., 2014; Ding et al., 2014), as well as through its interaction of different cytokines, such as IL-6 and TNF-α (Robbesyn et al., 2004). However, further investigation is needed to confirm one or the other hypothesis.
All pregnant uRM women participating to this study received an anticoagulant therapy (LMWH) from the first positive serum beta-HCG assay, and all of them had an at-term delivery. The use of LMWH in RM is based on the assumption that, in addition to its anticoagulant effect, LMWH has immunomodulatory and anti-inflammatory effects, which could antagonize the proinflammatory cytokine response involved in RM development (Matsumori et al., 1998; Clark et al., 1999; Arck et al., 2001; Wang et al., 2002; Erlebacher et al., 2004; Okada et al., 2004; Monien et al., 2009). Immunological effects of LMWH have been mainly studied in the context of innate immunity, showing, for example, effects on cellular adhesion and transmigration through the endothelium (Wan et al., 2001).
The LMWH also has an inhibitory action on the complement system and a protective mechanism against cell apoptosis (Downing et al., 1998; Girardi, 2005; Hills et al., 2006). A role of LMWH in the trophoblastic invasion and development and in the embryo implantation mechanism has also been proposed (Girardi, 2005; Di Simone et al., 2007; Fluhr et al., 2011; Tersigni et al., 2012). Even though there are no randomized double-blind trials that show the real efficacy of LMWH in RM, the scientific evidence previously described leads us to administer LMWH, since it has no important side effects. To the best of our knowledge, there are no studies on the expression of LMWH and LOX-1. Therefore, we may not rule out a role of the anticoagulant therapy (LMWH) in modulating the expression of OLR1 and Loxin, with a consequent effect in reducing the number of miscarriages.
In the past years, studies on animal models have reported the beneficial effects of statins, (inhibitors of the 3-hydroxy-3-methylglutaryl coenzyme A reductase which is the major rate-limiting enzyme in cholesterol biosynthesis) which can be useful in order to prevent pregnancy complications such as intrauterine growth restriction, RMs and preeclampsia (Redecha et al., 2008, 2009; Ahmed et al., 2010; Costantine et al., 2010). Treatments with pravastatin reduced OS and bad pregnancy outcomes in preeclamptic CBA/J × DBA/2 mice (Redecha et al., 2009). Moreover, it has been shown that pravastatin restored VEGF levels and placental perfusion and development in the CBA/J × DBA/2 model through two different mechanisms: inhibition of the release of the anti-angiogenic molecule sFlt-1 (soluble receptor for vascular endothelial growth factor) from macrophages (Redecha et al., 2009) and stimulation of vascular endothelial growth factor (VEGF) release from trophoblast (Ahmed et al., 2010). The release of sFlt-1 from macrophages causes defective placental development, intrauterine growth restriction and fetal death. Thus, the decrease of sFlt-1 and the increase of VEGF levels, induced by pravastatin, are able to restore the angiogenic balance in preeclamptic mice and rescue the pregnancies, also suggesting the promising therapeutic role of pravastatin for women with preeclampsia. Similarly, other studies also demonstrated that statins inhibit the release of sFlt-1 highlighting the potential of these molecules to prevent preeclampsia (Cudmore et al., 2007). Interestingly, exposure to statins disrupts LOX-1 receptor cluster distribution in plasma membranes and blocks LOX-1-mediated ox-LDL binding and internalization, suggesting that statins may protect vascular endothelium against the adverse effects of ox-LDLs by disruption of LOX-1 receptor function (Matarazzo et al., 2012; Biocca et al., 2015).
These data further support the key role of LOX-1 in pregnancy and in pregnancy complications, reinforcing the necessity of its deeper investigation.
Conclusions
The results of this pilot study suggest that an altered peripheral OLR1 and Loxin expression pattern could reflect a functional role of these splice isoforms in the dysregulation of OS controlling pathways working during pregnancy.
Furthermore, the reduction of OLR1 and Loxin expression in uRM women could be used in clinical practice for the detection of a selected population of RM patients showing no other risk factors that could explain the repeated occurrence of miscarriages.
Strengths and Limitations
The present study is a pilot study that for the first time highlights the role of OLR1 splice isoforms in the pathology of unexplained recurrent pregnancy loss. Nevertheless, it has some limitations. This study was a relatively small cohort and was conducted in a single center. Further detailed investigations from similar populations from other clinical centers are necessary to overcome these limitations. Even so, our results can be considered promising and worthy of further analysis.
In fact, there are evidence suggesting that women with a history of miscarriages or RMs are at a higher risk of coronary heart disease (CHD) and cardiovascular diseases (Oliver-Williams et al., 2013; Kessous et al., 2014; Heida et al., 2015), thereby indicating the possibility of a common mediator of these diseases shared among uRM women. Since LOX-1 is highly correlated with atherosclerotic and cardiovascular risk (Sawamura et al., 1997; Robbesyn et al., 2004), it could be of interest to perform a retrospective study in uRM women to evaluate the correlation between OLR1 and Loxin expression and to assess the women's risk of developing CHD.
Moreover, it is well known that seven different SNPs, located within intron 4 (rs3736232 G>C; rs3736234 C>T; rs3736235 A>G), intron 5 (rs17174597 A>G; rs13306593 G>T), and 3′-UTR (rs1050283 C>T) of the OLR1 gene and comprising a linkage disequilibrium block, are responsible for regulating the expression level of OLR1 and Loxin transcripts by modulating the retention/skipping of exon 5 (Mango et al., 2005). The results of this pilot study raise the possibility of exploring the association of these functional polymorphisms with uRM susceptibility, and also of investigating the effects of the SNPs mentioned earlier on LOX-1 expression. However, the small size of our samples group did not allow us to explore the influence of SNPs on OLR1 gene expression, remarking the necessity of an expansion of the case study.
Footnotes
Acknowledgment
The authors are grateful to the Paola Borgiani for kindly supporting them in conducting statistical analysis.
Author Disclosure Statement
The authors declare that no competing financial interests or any other conflicts of interest exist.