
Abstract
Aim:
The present study was designed to examine the relationship between gene polymorphisms of C1q, tumor necrosis factor-related apoptosis-inducing ligand (TRAIL), T cell immunoglobulin mucin (Tim-1), and systemic lupus erythematosus (SLE).
Materials and
Results:
As for the C1q gene, the frequency of the T allele at the rs631090 locus in the study group was lower than that in the controls, and the frequency of the C allele was higher in the study group than in the healthy donors. The frequency of the G allele at the 1525A/G locus of TRAIL gene in the study group was significantly higher than those in the control group. The frequency of the G allele at −1454G/A of Tim-1 was dramatically higher in the study group than in the control group. Anti-C1q antibody concentrations of subjects carrying CC and CT genotype at the rs631090 locus were statistically higher than TT genotype carriers. The sTRAIL protein level of the TRAIL 1525A/G GG genotype carriers was significantly higher than that of GA and AA genotype carriers, as well as CC genotype carriers at 1595T/C site compared with CT/TT genotype carriers. GG genotype carriers at −1454G/A had higher Tim-1 expression levels than GA/AA genotype carriers.
Conclusion:
The C allele at the rs631090 locus of C1q, the G allele at 1525A/G site of TRAIL, and the G allele of Tim-1 at −1454G/A site are susceptibility variants associated with SLE.
Introduction
S
Previous studies showed that the polymorphisms of rs292001 loci, rs631090 loci, and rs294223 loci of C1q gene were associated with the susceptibility to various diseases (Rafiq et al., 2010; Zakharyan et al., 2011; Mosaad et al., 2015b). However, the relationship between the abovementioned SNPs of C1q gene and their susceptibility to SLE in the Chinese population is still not clear. Luo et al. (2015) showed that the TRAIL gene polymorphisms at 1525A/G, 1588A/G, and 1595T/C sites were associated with the susceptibility to non-small-cell lung cancer (NSCLC) in China, but susceptibilities of TRAIL gene polymorphisms to SLE have been almost not reported. Similarly, another study from Mosaad et al. (2015a) showed that the TIM-1 rs41297579 G > A (−1454) polymorphism was associated with susceptibility to rheumatoid arthritis (RA), but whether the rs41297579 G > A (−1454) polymorphism is also related to the occurrence of SLE has not been studied yet. In this study, we evaluated the association of C1q, TRAIL, and Tim-1 gene polymorphism with the risk of SLE.
Materials and Methods
Subjects
A total of 245 patients with SLE in the General Hospital of Ningxia Medical University from February 2012 to August 2016 were enrolled in the study, including 20 males and 225 females. The study was approved by the Ethics Committee of the General Hospital of Ningxia Medical University, and written informed consent was obtained from each patient.
All SLE patients were diagnosed in line with the American College of Rheumatology Response Criteria for SLE Clinical Trials (2014) (American College of Rheumatology Ad Hoc Committee on Systemic Lupus Erythematosus Response, 2004). Patients were from 18 to 78 years of age, mean age 42.15 ± 8.15 years, course of disease from 7 to 59 months, and the average duration of disease was 35.10 ± 14.25 months. Patients taking a large dose of corticosteroids or anticytotoxic drugs within 1 month were excluded. Another 245 cases were recruited, including 21 males and 224 females from 18 to 75 years of age, with mean age 42.15 ± 8.15 years as healthy control. Patients with rheumatism history and family history of rheumatism were eliminated. There was no statistically significant difference in age, gender between study group, and normal control (p > 0.05).
Genomic DNA isolation and polymerase chain reaction
Genomic DNA was isolated from the peripheral blood of subjects using the QIAmp DNA Blood Mini Blood Kit (Qiagen, Germany). Polymerase chain reaction (PCR) primers for C1q, TRAIL, and Tim-1 amplification were concluded in Table 1. PCR was carried out in a total volume of 20 μL containing 10 ng of genomic DNA, 2 μL 10 × Buffer, 200 nM dNTPs (Fermentas, MA), 0.8 μL MgCl2 (2.5 mM), 6 pmol of each primer, and 0.4 U of KOD-Plus polymerase (Toyobo, Japan). The amplification was conducted as follows: 1 cycle at 94°C for 2 min, followed by 35 cycles of 94°C for 15 s, 56°C (C1q gene) for 30 s, 68°C for 40 s, and 68°C for 10 min. As for TRAIL and Tim-1 amplification, the annealing temperature was 55°C and 60°C, respectively. Purified PCR products were sequenced by ABI automatic DNA sequencing. The genotypes were determined according to the sequencing results.
PCR, polymerase chain reaction; TIM-1, T cell immunoglobulin mucin; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand.
Quantitative determination of anti-C1q, sTRAIL, and Tim-1 in serum
The levels of anti-C1q, sTRAIL, and Tim-1 in serum of both SLE patients and controls were quantified by enzyme-linked immunosorbent assay (ELISA). Serum samples that were collected were diluted in 1/100 PBS buffer and were placed in wells coated with complement C1q antigen, antibody against sTRAIL, or Tim-1, respectively. Samples were treated following common protocols. The amount of converted substrate was photometrically determined at 450 or 620 nm. The amount over 10 U/mL was defined as anti-C1q antibody positive. The ELISA Kit for anti-C1q (Cat No. xy-A747Hu) and sTRAIL (Cat No. xy-E10129) detection were from BioLegend (CA), the ELISA Kit for Tim-1 (Cat No. EHHAVCR1) was brought from Thermo Fisher Scientific (MA).
Statistical analyses
SPSS 20.0 software (IBM, NY) was used for the statistical analysis. The measurement data were expressed as (
Results
Demographic information
A total of 245 cases of SLE patients 18 to 78 years of age with a duration of 7 to 59 months were included in this research. To exclude the influence of different races, all SLE patients were from the Chinese Han population. Demographic data of SLE patients are shown in Table 2.
Genotype and allele distribution of polymorphism in C1q gene
The genotype and allele distribution of C1q polymorphism in the study group and the control group were in Hardy-Weinberg equilibrium (p > 0.05). There were significant differences of multiple genotype frequencies at C1q rs631090 locus in both the study group and the control group (p < 0.05). Specifically, the study group had lower frequency of the T allele, but higher frequency of the C allele of C1q gene at rs631090 (χ2 = 15.551, p < 0.001). C1q rs292001 and rs294223 sites had no difference of genotypic and allelic frequencies between the study and control groups (p > 0.05) (Table 3, Fig. 1A-C).
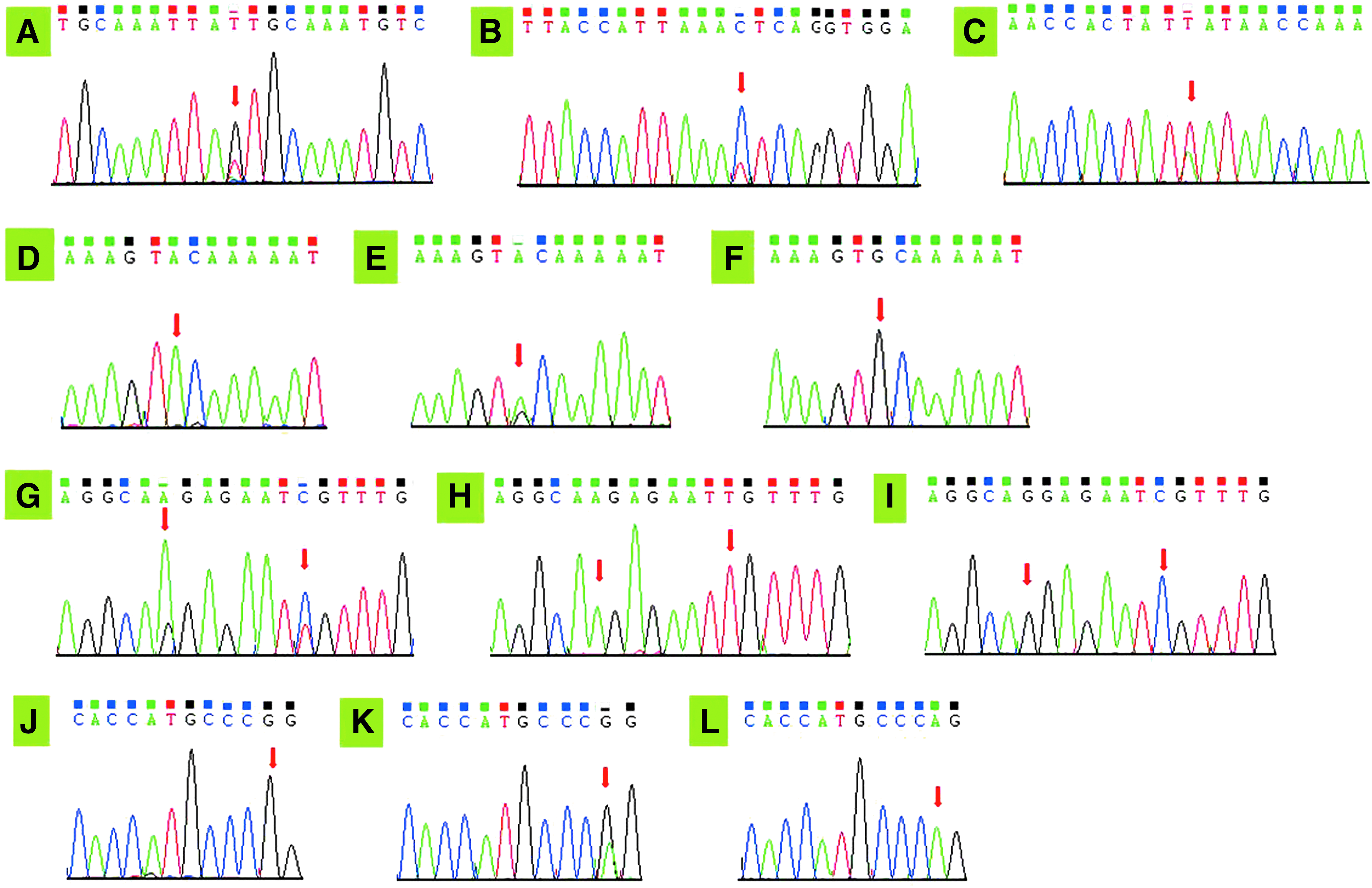
Sequencing chromatogram of gene polymorphism in C1q, TRAIL, and Tim-1. The red arrows indicate the location of the mutation.
Genotype and allele distribution of polymorphism in TRAIL
All the genotype and allele distribution of TRAIL polymorphism in both of the study and control group were in accordance with Hardy-Weinberg equilibrium (p > 0.05). The frequency of A alleles at TRAIL 1525A/G locus of SLE patients was significantly lower than that of the control group (p < 0.001), whereas the allele frequencies of the 1588A/G locus and 1595T/C locus displayed no significant difference between the study group and the control group (p > 0.05) (Table 3, Fig. 1D-I).
Genotype and allele distribution of polymorphism in Tim-1
Departure from Hardy-Weinberg equilibrium was observed neither in the study group nor in the control (p > 0.05). Significant difference in frequencies among different genotypes and alleles of Tim-1 −1454G/A locus occurred in both the study and control groups (p < 0.05). Moreover, SLE patients had higher G allele frequency at the −1454G/A site of Tim-1 gene than healthy donors (χ2 = 19.067, p < 0.001) (Table 3, Fig. 1J-L).
The protein levels of C1q, TRAIL, and Tim-1
ELISA test revealed that in the SLE patient group, anti-C1q antibody levels were significantly higher (p < 0.001) (Fig. 2A) and the levels of sTRAIL and Tim-1 were statistically lower than in the control group (p < 0.001) (Fig. 2B, C). Based on ELISA data, individuals carrying AG, AA, or GG genotype at the C1q rs294223 site had comparable protein levels of anti-C1q antibody (p > 0.05). The same result was observed among individuals with the rs292001 AG, AA, or GG genotype (p > 0.05). As shown in Figure 3A-C, the C1q antibody levels of subjects with CC and CT genotype at C1q rs631090 locus were significantly higher than TT genotype carriers (p < 0.05). As for TRAIL, sTRAIL levels of individuals carrying GG at both 1525A/G and 1588A/G sites or CC at 1595T/C were significantly higher than GA/AA genotype or CT/TT carriers, separately (p < 0.05) (Fig. 3D-F). In addition, Tim-1 protein levels of individuals carrying GG at the −1454G/A site were also observed to be much higher than GA/AA genotype carriers (Fig. 3G). All the mentioned differences were statistically significant (p < 0.05).
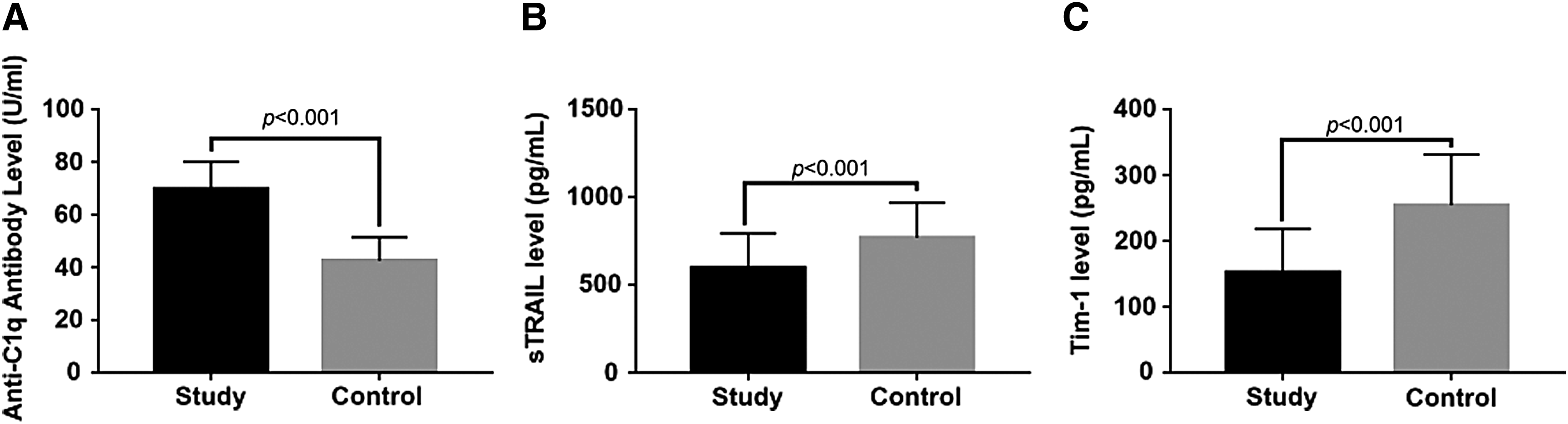
The protein levels of C1q, TRAIL, and Tim-1 between SLE patients and health control.

The protein levels of C1q, TRAIL, and Tim-1 corresponding gene locus.
Discussion
As a recognition molecule, C1q is an important component of complement system, composed of 18 peptide chains known as 6A, 6B, and 6C. C1q initiates the classical complement pathway activation through binding to immunocomplex, C-reactive protein, and the ligands on the surface of apoptotic cells (Kishore and Reid, 2000; Agrawal et al., 2001). In the active phase of SLE, complement system, especially C3 will be deposited in the body site with inflammatory reaction. Dysfunction of complement system may induce the occurrence and development of SLE. Several studies found that SLE patients with inherited complement deficiency appeared to have higher morbidity than the normal population (Cook and Botto, 2006; Barilla-Labarca et al., 2013; Leffler et al., 2014). The C1q gene cluster comprises three genes, C1qA, C1qB, and C1qC. Only one SNP site on each gene was selected for this study, including rs292001 (C1qA), rs631090 (C1qB), and rs294223 (C1qC), respectively. Until now a number of complement system genes related to SLE susceptibility have been identified, including C1q, C2, C4, etc.
This study analyzed the association between the susceptibility to SLE and gene polymorphism of the C1qB rs631090 locus. Statistical analyses revealed that the frequency of the T allele in the study group was lower than that of normal controls, whereas the frequency of C allele was higher (p < 0.001). The serum quantitative determination showed that anti-C1q antibody levels of individuals with CC and CT genotype at C1q gene rs631090 locus was significantly higher than that of TT genotype carriers (p < 0.05). These results demonstrated that the C1q gene polymorphism, especially the C allele of the C1q rs631090 locus, was associated with SLE susceptibility, which was consistent with the previous investigations (Martens et al., 2009).
The rs292001 and rs294223 loci of the C1q gene are located in the of C1q noncoding region, so that they will not affect the expression of C1q protein. The rs631090 locus is located in the expression regulatory region of the C1q, where the C1q protein will be abnormally high after T > C mutation. Mosaad et al. (2015b) found that in adolescent SLE (jSLE) patients in Egypt, the A allele of C1q rs292001, especially the AA genotype was associated with jSLE. However, no association between polymorphism of rs292001 and SLE was found in this study, indicating that the rs292001 locus polymorphism may be related to race. Martens et al. (2009) showed that the SNP at the C1q rs631090 locus was significantly associated with SLE and the level of serum C1q, which was consistent with results of this study. Another study in Europeans also proved that the polymorphism of rs294223 had no correlation with SLE (Rafiq et al., 2010).
TRAIL, one member of TNF family, plays an important role in tumor development, tumor metastasis, and autoimmune diseases (Allen et al., 2015; Lim et al., 2015). Through activating exogenous apoptotic pathways, TRAIL participates in a series of immune responses, including immune regulation, surveillance, and immune homeostasis. The TRAIL gene is located on chromosome 3q26, which contains five polymorphic sites. The 1525A/G site, the 1588A/G site, and the 1595/C locus analyzed in this study are located in the 3′ untranslated region (3′UTR), whereas the other two polymorphic sites are located on exon 1 and exon 5.
This study showed that the A allele at −1525 A/G locus was a protective factor for SLE. A previous study found that Chinese populations carrying the A allele at the 1525 A/G locus of TRAIL had lower risk of fatty liver disease (Yan et al., 2009). Besides, Chinese nonalcoholic fatty liver disease patients carrying the A allele at the 1525 A/G locus of TRAIL also displayed lower degree of steatosis. No association between polymorphisms and susceptibility to SLE was found in other sites within TRAIL gene (p > 0.05). ELISA data revealed that subjects with GG genotype at 1525A/G and 1588A/G sites had higher serum sTRAIL protein level compared with GA or AA genotype carriers (p < 0.05). In addition, individuals with CC genotype at 1595T/C loci had more serum sTRAIL protein than CT or TT genotype carriers (p < 0.05).
This result may be caused by the reason that the 1525A/G, 1588A/G, and 1595T/C sites are located in the TRAIL expression regulatory region, where gene polymorphism will lead to the decrease of sTRAIL protein expression. However, only the polymorphism of 1525A/G locus correlates with the occurrence of SLE, indicating that the mentioned association cannot directly affect sTRAIL expression. There may be other mechanisms involved in the regulation, which remains to be further studied.
Tim-1, also known as HAVCR1, is encoded by HAVCR1 gene. The encoding genes of the Tim family are located on chromosome 5q33.2. Tim-1 is highly expressed in Th2 cells, which regulates Th1/Th2 responses by modulating Th2 cytokine expression (Santiago et al., 2007; Kanzaki et al., 2012). Tim-1 not only binds phosphatidylserine to affect apoptotic cell clearance, but also regulates the immune function of B cells (Uchida et al., 2010). It has been reported that the localization of Tim-1 on chromosome is overlapped with the SLE-associated region (García-Lozano et al., 2010). Moreover, the correlation between Tim-1 polymorphism and autoimmune disease has been confirmed by researchers (Mazrouei et al., 2016; Yuan et al., 2016).
Our results revealed that the frequency of the G allele at Tim-1 −1454G/A locus in the study group was significantly higher than that in normal controls (p < 0.001). Besides, Tim-1 protein level of the carriers with GG genotype at −1454G/A site was significantly higher than that in a population with GA or AA genotype (p < 0.05). All the experimental data showed that the gene polymorphism of Tim-1 −1454G/A site was associated with the susceptibility to SLE. Moreover, carriers with the G allele at −1454G/A had a higher level of Tim-1 in serum, indicating that such individuals had a higher risk of SLE. The Tim-1 protein level in serum of G allele carriers was positively correlated with the susceptibility to SLE, which was consistent with these relevant reports (Xiao et al., 2012). The analysis suggested that the −1454G/A mutation might be located in the Tim-1 gene expression regulatory region, so that mutant individuals had a lower Tim-1 protein level. One study in Southeast Asians presented a hypothesis that the polymorphism at the −1454G/A locus of Tim-1 was associated with the severity of malaria because A allele at −1454G/A locus could promote the expression of Tim-1 protein to protect against cerebral malaria (Nuchnoi et al., 2008), which was consistent with our study. The C allele of C1q at rs631090 locus, the G allele of TRAIL at 1525A/G sites, and the G allele of Tim-1 at −1454G/A loci are susceptible genes of SLE.
Acknowledgments
This study was supported by the Scientific and Technological Project of the Ningxia Hui Autonomous Region (Grant No.: 2011056).
Footnotes
Author Disclosure Statement
No competing financial interests exist.