
Abstract
Objectives:
MicroRNA-155 (miR-155) is an important regulator of immune responses in humans. However, its role in T-cell activation in hepatitis B virus (HBV) infection remains unclear.
Materials and Methods:
Eighty-one patients with chronic hepatitis B (CHB), 77 HBV carriers, and 51 healthy controls were recruited. HBV DNA and serologic tests were carried out for each subject. Levels of miR-155 in peripheral blood were detected by quantitative reverse transcription/polymerase chain reaction. Immune activation of T-cells was determined by detection of surface molecules CD38 and HLA-DR using flow cytometry.
Results:
We found higher miR-155 levels in CD4+ and CD8+ T-cells of CHB patients than HBV carriers or healthy controls (p < 0.01), moreover, miR-155 levels in the CD8+ T-cells of HBV carriers were higher than in healthy controls (p < 0.01). Furthermore, immune activation of CD4+ and CD8+ T-cells in CHB patients was much higher than in healthy controls (p < 0.01).
Conclusion:
Our findings suggest that miR-155 expression positively correlates with T-cell activation, especially in CHB patients, and is a potential biomarker for immune activation and disease progression in HBV infection.
Introduction
Despite the availability of effective vaccines, hepatitis B virus (HBV) infection remains a global public health concern. Eighty-two percent of hepatocellular carcinoma is related to viral hepatitis and 55% to HBV (Michielsen and Ho, 2011). The clinical outcome of HBV infection depends on the host immune response. T-cell immune responses are very important in the pathogenesis and control of HBV infection, as the CD8+ T-cell response plays a major role in controlling virus replication (Bertoletti and Gehring, 2006; Baumert et al., 2007). Compared with acute self-limiting infections, lack of an intensive and specific T-cell immune response to HBV is the major cause of failure of viral clearance and disease progression in chronic hepatitis B (CHB) patients (Bertoni et al., 1997; Zhang et al., 2012).
Immune exhaustion and dysfunction of T-cells are thought to be the main reason of persistent viral replication (Barathan et al., 2015, 2016; Gogoi et al., 2015; Saeidi et al., 2015; Ellegård et al., 2018; Yong et al., 2018). A major factor contributing to immune exhaustion is persistent, chronic nonspecific immune activation of T-cells. CD38 and HLA-DR expression has been established as a chronic activation marker of T-cells (Rosso et al., 2010; Jin et al., 2012, 2017a; Greenough et al., 2015). Higher CD38 and HLA-DR expression levels on T-cells were observed in many acute and chronic infections, including human immunodeficiency virus (HIV) among others (Benito et al., 1997; Rosso et al., 2010; Jin et al., 2012, 2017; Greenough et al., 2015). Shankar's group has investigated immune activation and exhaustion of T-cells in chronic HBV and hepatitis C virus infection. They found that increased activation (CD38 and HLA-DR expression) is related to T-cell immune dysfunction, such as increased PD-1, TIM-3, and CTLA-4 and decreased CD127 and costimulatory markers (Barathan et al., 2015, 2016; Saeidi et al., 2015; Ellegård et al., 2018; Yong et al., 2018). Higher proportions of CD8+CD38+ T-cells in CHB patients have been reported compared with HBV carriers and healthy controls, indicating that immune activation associated with HBV may be a promising target for treatment (Cao et al., 2011).
MicroRNAs are small noncoding RNAs that function in RNA silencing and posttranscriptional regulation of gene expression (Jin et al., 2016). Many microRNAs can be used as disease biomarkers (Jin et al., 2014). Compared with other laboratory tests, such as flow cytometry, detection of microRNAs is much easier and cheaper, and samples can be stored at −80°C for long time. So it is very necessary to find microRNAs that can be used as markers of T-cell immune activation in virus infection, including HBV infection. MicroRNA-155 (miR-155) is a newly discovered miR-155 that regulates the immune response at multiple levels (Seddiki et al., 2014). miR-155 is an important regulator of B cell differentiation and antibody production (Landgraf et al., 2007). It is found to be greatly increased in activated CD8+ T-cells and may regulate CD8+ T-cell responses (Martin et al., 2006; Salaun et al., 2011; Jin et al., 2017b). miR-155 also plays a vital role in T-cell responses against infections (Lind et al., 2013). Jin et al. (2017a) reported that peripheral miR-155 levels in HIV-1-infected patients were increased and associated with T-cell activation. However, it is still unclear whether miR-155 is related to T-cell activation in HBV infection.
In this study, we investigated levels of miR-155 in CD4+ and CD8+ T-cells from CHB patients, HBV carriers, and healthy controls, and analyzed the correlation between miR-155 levels and immune activation of T-cells. Our results showed that the expression levels of miR-155 in the peripheral T-cells of HBV-infected patients (comprising both CHB patients and HBV carriers) were increased, and this was also positively correlated with enhanced immune activation of T-cells. Therefore, our findings indicate that miR-155 expression is correlated with T-cell activation and may be a potential biomarker of immune activation in HBV infection.
Materials and Methods
Patients
A total of 158 patients positive for hepatitis B surface antigen (HBsAg) were enrolled from January 1 to May 31, 2017, at the outpatient clinic of the First Affiliated Hospital of Zhejiang University, the Second Affiliated Hospital, and Yuying Children's Hospital of Wenzhou Medical University. Among them, 81 patients were CHB patients with the following clinical, biochemical, and virological evidence: HBsAg (+), hepatitis B e antigen (HBeAg) (+), anti-hepatitis B e antibody (HBeAb) (−), anti-hepatitis B core antibody (HBcAb) (+), increased alanine aminotransferase (ALT) levels (range: 74-315 U/L), and higher HBV DNA levels (range: 7.3 × 104-2.6 × 108 copies/mL). Seventy-seven patients were HBV carriers with HBsAg (+), fluctuating HBVDNA levels, and normal ALT. The diagnosis and inclusion criteria were based on the American Association for the Study of Liver Diseases Guidelines 2015 on CHB (Terrault et al., 2016). Our study also included 51 healthy subjects as controls. All participants in the three groups were negative for other viral infections and immune system diseases. This study was approved by the Ethics Review Boards of all three recruiting hospitals. All subjects were volunteers and provided written informed consent to participate in the study.
Laboratory tests
HBV antigens and antibodies (HBsAg, HBsAb, HBeAg, HBeAb, and HBcAb) were measured by enzyme-linked immunosorbent assay (ELISA) (Abbott, Inc., Santa Clara, CA). An automated system, the COBAS AmpliPrep/COBAS TaqMan 48 analyzer (CAP/CTM) (Roche, Branchburg, NJ), was used for HBV DNA extraction and quantitative real-time polymerase chain reaction (qRT-PCR) determination of HBV DNA load in serum, with a lowest limit of detection of ∼103 viral copies/mL. All experimental procedures were performed according to the manufacturer's instructions.
Quantitative real-time polymerase chain reaction
About 2 mL of whole blood was collected in a tube containing ethylenediaminetetraacetic acid from every participant. Peripheral blood mononuclear cells (PBMCs) were isolated using Ficoll-Paque PLUS (GE Healthcare Life Science, Marlborough, MA). CD4+ and CD8+ T-cells were purified from PBMCs using magnetic-activated cell sorting human CD4 and CD8 microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany), with a purity of >90%. Total RNA was extracted from CD4+ to CD8+ T-cells using TRIzol (Life Technologies, Carlsbad, CA). Levels of miR-155 were detected by qRT-PCR using a commercial Bulge-Loop™ miRNA qRT-PCR Starter Kit (RiboBio, Guangzhou, China) and an hsa-miR-155 qRT-PCR primer set (RiboBio). U6 small nuclear RNA was used as the internal control. The experiments were performed according to the manufacturer's instructions. Fold change in miR-155 expression relative to control was determined by the standard 2−ΔΔCt method.
Flow cytometry
Absolute cell counts and subsets of peripheral T-cells were analyzed via four-color flow cytometry using commercial flow cytometry antibodies (Becton Dickinson, Franklin Lakes, NJ) on an FACSCanto II (Becton Dickinson). Cell staining was performed according to the protocol described by the manufacturer. The absolute cell counts for CD3+CD4+ and CD3+CD8+ T-cells were determined using anti-CD3−APC, anti-CD4−FITC, and anti-CD8−PE-Cy7 (Becton Dickinson). Markers of immune activation (CD38 and HLA-DR) on the surface of CD4+ and CD8+ T-cells were analyzed using anti-CD38-PE and anti-HLA-DR-PE-Cy5.5 (Becton Dickinson). The percentage of CD3+CD4+CD38+, CD3+CD4+HLA-DR+, CD3+CD8+CD38+, and CD3+CD8+HLA-DR+ T-cells was assessed.
Statistical analyses
Statistical analyses were performed using SPSS for Windows version 20.0 (SPSS, Chicago, IL). Student's t-test was used to compare between two groups, and a one-way analysis of variance was used when comparing more than three groups. The correlation was tested using Spearman's correlation test. All tests were two tailed, and p values <0.05 were considered statistically significant.
Results
The clinical information of subjects
The three groups of participants were all appropriately age and gender matched. The mean serum ALT and total bilirubin levels in CHB patients were much higher than in HBV carriers and healthy controls (p < 0.01). No significantly different HBV DNA loads were found between CHB patients and HBV carriers. We found lower CD3+CD4+ T-cell counts in CHB patients (p < 0.05), but no difference in the percentage of CD3+CD4+ T-cells between HBV carriers and healthy controls. The percentages of CD3+CD8+ T-cells in both CHB patients and HBV carriers were higher than in healthy controls (p < 0.05). Detailed clinical data are presented in Table 1.
Clinical Data for the Study Participants
p < 0.05, **p < 0.01, compared with healthy controls; #p < 0.05, ##p < 0.01, compared with HBV carriers.
ALT, alanine aminotransferase; CHB, chronic hepatitis B; HBV, hepatitis B virus; NA, not applicable; TBIL, total bilirubin.
Increased miR-155 levels in peripheral blood CD4+ and CD8+ T-cells of CHB patients
We first isolated CD4+ and CD8+ T-cells from PBMCs and detected miR-155 expression levels in CD4+ and CD8+ T-cells. We found significantly higher miR-155 levels in CD4+ and CD8+ T-cells of CHB patients compared with HBV carriers and healthy controls (p < 0.05 formiR-155 levels in CD4+ between CHB patients and HBV carriers; p < 0.01 for comparisons between other groups) (Fig. 1a, b). Furthermore, the miR-155 levels in CD8+ T-cells of HBV carriers were higher than those of healthy controls (p < 0.01) (Fig. 1b). These results indicate that miR-155 levels are increased in CD4+ and CD8+ T-cells of CHB patients. The detailed data were shown in Supplementary Data.
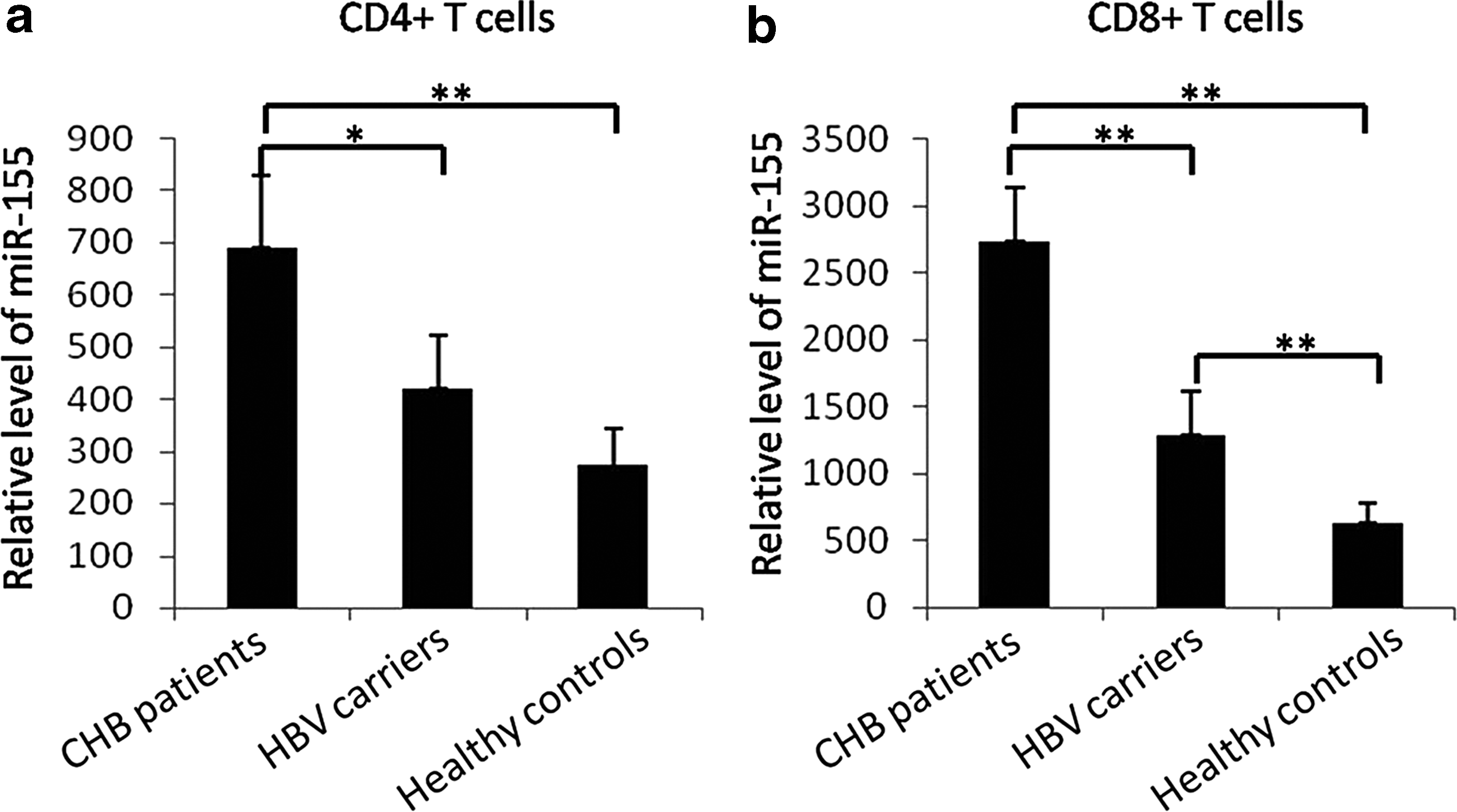
miR-155 levels in peripheral T-cells of HBV patients and healthy controls. miR-155 levels in
Increased CD38+ and HLA-DR+ T-cell subsets were found in hepatitis B patients
We then detected immune activation of CD4+ and CD8+ T-cells in HBV patients and healthy controls by testing the percentage of T-cells expressing CD38 and HLA-DR. The results showed that immune activation (both CD38+ and HLA-DR+) of CD4+ and CD8+ T-cells in CHB patients was much higher than that in healthy controls (p < 0.01) (Fig. 2a-d). The percentage of CD8+CD38+ T-cells in CHB patients was also higher than that in HBV carriers (p < 0.05) (Fig. 2b). Immune activation of CD4+ T-cells in HBV carriers was higher than that in healthy controls (p < 0.05) (Fig. 2a, c), but no difference in the percentage of CD8+CD38+ and CD8+ HLA-DR+ T-cells was found between HBV carriers and healthy controls. The detailed data were shown in Supplementary Data.

T-cell immune activation of HBV patients and healthy controls. Immune activation levels of CD4+
miR-155 expression positively correlates with immune activation in T-cells of hepatitis B patients
Finally, we analyzed the correlation between immune activation and miR-155 expression in CD4+ and CD8+ T-cells. In CHB patients, we found that expression of activation markers in both CD4+ and CD8+ T-cell subsets positively correlated with miR-155 expression (Table 2). Immune activation of T-cells in HBV carriers also showed a positive relationship with miR-155 expression, except for CD4+HLA-DR+ T-cells. In healthy controls, a correlation was found between HLA-DR+CD4+/CD8+ T-cells and miR-155 expression (Table 2). Our results indicate that miR-155 may be an important regulator of immune activation in HBV infection.
Correlation Between microRNA-155 Levels and Immune Activation
All correlations are reported as Spearman r and p values (two tailed). Significant values are shown in bold.
miR-155, microRNA-155.
Discussion
In addition to HBV DNA levels and liver function, the activation of T-cells is vital for evaluation of disease progression during HBV infection (Bertoletti and Gehring, 2006). The activation of T-cells, especially CD8+ T-cells, plays an important role in virus clearance (Peng et al., 2015; Ye et al., 2015), yet excessive activation of CD8+ T-cells can also lead to immune-mediated damage and aggravation of disease progression. Cao et al. (2011) have reported persistent chronic immune activation in patients with CHB. Our study found that HBV infection could cause significantly increased T-cell activation, which was higher in CHB patients than in HBV carriers.
miR-155 is critical for T-cell fitness and effector function, and selective deletion of miR-155 leads to severe defects in immune responses (Seddiki et al., 2014). miR-155 has been reported to impact disease progression in HIV-1 infection by regulating the transformation of naive regulatory and CD4+ T-cell subsets into activated subsets (Bignami et al., 2012; Seddiki et al., 2012). Jin et al. (2017a) found that miR-155 levels in the peripheral blood of HIV-1 patients were increased and associated with T-cell activation, indicating that miR-155 is a potential biomarker of the immune response following HIV-1 infection. In HBV infection, overexpression of miR-155 can inhibit HBx expression and help to reduce the hepatitis B viral load (Su et al., 2011). However, an in vivo study showed that miR-155 expression level in PBMCs is downregulated in HBV CHB patients (Yu et al., 2016). To date, no studies have described miR-155 expression levels and the relationship with immune activation in HBV infection. Therefore, we conducted a cross-sectional analysis of miR-155 expression in HBV-1-infected individuals (comprising both CHB patients and HBV carriers) to understand the effect of miR-155 on HBV infection and immune activation in this disease.
We found that there were significantly increased levels of miR-155 in CD4+ and CD8+ T-cells from HBV-infected patients compared with healthy controls, and that this increase was greatest in CHB patients. The higher miR-155 levels in CD4+ and CD8+ T-cells of CHB patients were consistent with their increased T-cell activation, which was confirmed by the positive correlation between miR-155 expression and CD4+ and CD8+ T-cell activation levels. Notably, the positive correlation between miR-155 and CD8+ T-cell activation was stronger than that between miR-155 and CD4+ T-cell activation. Indeed, miR-155 can affect multiple targets and genes associated with type I IFN signaling and response, and is also important for CD8+ T-cell proliferation (Gracias et al., 2013; Cheng et al., 2015). In herpes virus infection, miR-155 was reported to be an important regulator in the generation and maintenance of both effector and effector memory CD8+ T-cells (Tsai et al., 2013). Our results indicate that miR-155 is an important regulator of T-cell immune activation, especially for CD8+ T-cells. Our results are inconsistent with two recent studies that indicated lower miR-155 levels in PBMCs and natural killer cells of CHB patients (Yu et al., 2016; Ge et al., 2017). However, PBMCs are a mixture of immune cells and the functional aspects are uncertain. In fact, these studies showed that CHB patients with elevated ALT had higher levels of miR-155 (Yu et al., 2016), and lower expression of miR-155 contributes to dysfunction of natural killer cells (Ge et al., 2017), indicating the important role of miR-155 in immune cell activation. Studies revealed that the function and quality of immune cells are important for immune activation and response (Barathan et al., 2015, 2016; Gogoi et al., 2015; Saeidi et al., 2015; Ellegård et al., 2018; Yong et al., 2018). The different miR-155 expression levels in different immune cells or even in different patients may be related to the functional aspects of the cells.
Conclusion
In summary, our study found increased expression levels of miR-155 in peripheral CD4+ and CD8+T-cells of HBV-infected patients, especially in CHB patients. This increased expression also positively correlated with enhanced immune activation of T-cells. Our findings indicate that miR-155 expression is correlated with T-cell activation and may be a potential biomarker of immune activation and disease progression in HBV infection. However, our study is cross sectional and small in scale, and thus, it is difficult to comment on causality. A deeper understanding of miR-155 in HBV infection, especially the relationship between functional aspects of T-cells and miR-155 expression, should be pursued in future prospective studies.
Footnotes
Acknowledgment
We thank all the participants in this study.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.