
Abstract
Background:
Gastric cancer (GC) is one of the most prevalent malignant tumors and has a very high incidence of mortality. Recently, long noncoding RNAs (lncRNAs) have been demonstrated to play an important role in the development and progression of GC.
Materials and Methods:
In the present study, we investigated the biological function and molecular mechanisms of lncRNA MRPL39 in GC.
Results:
We found that MRPL39 was significantly downregulated in GC tissues and cell lines and that its expression level was negatively associated with carcinoma size, tumor, lymph node, metastasis (TNM) stage, and lymphatic metastasis. Patients with low MRPL39 expression levels revealed a short overall and disease-free survival period. Over-expression of MRPL39 in the GC cell lines BGC823 and SGC-7901 inhibited cell growth, proliferation, migration, and invasion. MiR-130, a putative target gene of MRPL39, displayed an inverse association with the expression of MRPL39 in GC tissues and cell lines. Moreover, a luciferase assay demonstrated a direct binding between the miR-130 and MRPL39, and the reintroduction of miR-130 abrogated the anti-tumor effect of MRPL39 on GC cells.
Conclusion:
Taken together, these findings indicate that MRPL39 serves as a tumor suppressor by directly targeting miR-130 in GC, which suggests that it might be a novel biomarker in the diagnosis and prognosis of GC.
Introduction
G
As the development of the human genome project, a large quantity of mammalian genomes are identified and found to be transcribed to noncoding RNAs (ncRNAs) (Esteller, 2011). LncRNAs, belonging to a group of ncRNAs, are generally identified as 200 nt-100 kb long transcripts without the open-reading frame (Troy and Sharpless, 2012; Bolha et al., 2017). Mountain evidences have demonstrated that lncRNAs are involved in a variety of biological functions, including chromosome remodeling, reprogramming stem cell pluripotency, and immunoregulation (Batista and Chang, 2013; Rogoyski et al., 2017). In addition, lncRNA expression was reported to be dysregulated in a number of cancers, such as hepatocellular carcinoma, colorectal cancer, and cervical squamous cancer (Ning et al., 2016; Chang et al., 2017; Lan et al., 2017; Qian et al., 2017; Wang et al., 2017b). Increasing studies have indicated that dysregulation of lncRNAs plays important roles in GC, and growing lncRNAs are identified as oncogenes or antioncogenes in GC, but the underlying molecular mechanism is still under investigation (Fu et al., 2016; Huang et al., 2016; Shao et al., 2016). A recent study found that MRPL39 was obviously lower in advanced GC tumor tissues than adjacent nontumor tissues, which may act as a possible regulator in GC development (Li et al., 2016).
In this study, we aimed to investigate the biological function and associated molecular mechanisms of MRPL39 in GC. The MRPL39 expression and clinicopathological characteristics were detected in GC patients. The function of MRPL39 on cell proliferation and migratory and invasive capabilities was examined in GC cells. In addition, we assessed the correlation between the MRPL39 and miR-130 in GC cells. According to bioinformatic analysis, we found that MRPL39 acts as a molecular sponge for miR-130. For the first time, we present evidence of molecular mechanisms of MRPL39 in carcinogenesis and metastasis of GC.
Materials and Methods
Patients and tissue samples
Sixty frozen GC specimens and paired adjacent normal gastric tissues were from the Department of Surgery, Hangzhou Third Hospital. All specimens were immediately divided into two parts: one part was snap-frozen in liquid nitrogen and stored at −80°C for RNA extraction, and the remaining part was fixed in 4% paraformaldehyde for immunohistochemical analysis. All samples were rediagnosed and identified by two pathologists in our hospital, respectively. Then, we followed up all the GC patients every 5 months after surgery for 5 years. The clinical characteristics were recorded, including gender, age, tumor size, tumor, lymph node, metastasis (TNM) stage, and lymphatic metastasis. Overall and disease-free survival rate was defined as the time period from surgery date to the death of the patients or last contact. Informed consents were obtained from the GC patients and this research was approved by the Ethics Committee of Hangzhou Third Hospital.
Cell culture
The human GC cell lines (BGC823, AGS, and SGC-7901) and normal gastric cell line (GES-1) were purchased from Academy of Sciences of China (Shanghai, China). The cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Gaithersburg, MD) containing 10% fetal bovine serum (Gibco) and were incubated in a humidified atmosphere of 37°C and 5% CO2.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from clinical samples or GC cell lines with TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. All the extracted RNA was reverse-transcribed under standard conditions utilizing the Reverse Transcription Kit (Applied Biosystems, Foster City, CA). Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using SYBR Premix Ex Taq (Takara, Dalian, China) on an Applied Biosystems 7500 Real-Time PCR System (Applied Biosystems). Relative gene expression was examined with a comparative cycle threshold method. The expression of lncRNA, mRNA, and microRNA (miRNA) was normalized to U6 small nuclear RNA. Relative quantification was calculated using the 2−ΔΔCt method.
Cell transfection
The full length sequence of MRPL39 was PCR amplified by PrimeSTAR HS DNA Polymerase (TaKaRa, Shanghai, China) from GES-1 cells and cloned into the PcDNA3.1 vector (Invitrogen). Lipofectamine 2000 reagent (Invitrogen) was applied to transfect the cells when their confluency reached 70% according to the manufacturer's instructions. At 48 h post-transfection, the cells were collected for further analysis.
Cell proliferation
Cell proliferation was assessed using a Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto, Japan). A total of 3 × 103 GC cells were seeded in 96-well plates with three repeat wells in each group. After incubation of 1, 2, 3, and 4 days, a 10 μL CCK-8 was added to each well and cultured at 37°C for another 2 h. The absorbance of each well at 450 nm for each group was detected using a Benchmark Microplate Reader (Bio-Rad Laboratories, Inc., Hercules, CA).
Immunohistochemical staining assay
The paraffin-embedded tissue blocks were cut into 4 μm sections and immunohistochemistry (IHC) staining was performed following the previous methods (Mao et al., 2017). Briefly, the slices were incubated with the primary antibodies (mouse anti-human Ki67, dilution 1:500; Abcam, Cambridge, United Kingdom) overnight at 4°C followed by incubating with a secondary antibody labeled by horseradish peroxidase (mouse anti-HRP, dilution 1:500; Abcam) for 30 min at room temperature. After diaminobenzidine working solution and counterstained with hematoxylin (HercepTest; Dako, Glostrup, Denmark), the Ki67 expression in GC tissue sections was analyzed using light microscope.
Transwell invasion assay and wound healing assay
Invasion assay was conducted using the 24-well transwell chamber (Corning, Inc., Corning, NY). Briefly, 1 × 105 pretransfected cells were seeded into the upper chamber containing serum-free DMEM. To the lower chamber was added 0.7 mL DMEM with 10% fetal bovine serum. The cells were stained with 1% crystal violet dye for 10 min. The invasion ability was determined by the ratio between invaded cells and the total cells on both sides of transwell membrane. To detect cell migration, wound healing assay was performed. A sterile 200 μL pipette tip was used to make a uniform scratch at the center of the well when the cell reached full confluence. Cells were washed with phosphate-buffered saline to remove cell debris and then incubated in serum-free DMEM for 24 h. The horizontal distance between the sides of the wound was measured.
Vector construction and luciferase reporter assay
The bioinformatic databases (LNCipedia.org, https://lncipedia.org/db/search) were applied to explore the putative miRNAs that bind to MRPL39. The putative wild-type and mutant sequence of MRPL39 was cloned into a pGL3 luciferase vector (Promega Corporation, Madison, WI) to form the MRPL39-mutant-type (Mut MRPL39) and MRPL39-wild-type (WT MRPL39) reporter vector. Afterward, the BGC-823 and SGC-7901 cells were cotransfected with the reporter vectors and miR-130 or control mimics (GenePharma, Shanghai, China) using Lipofectamine 2000 Reagent (Invitrogen; Thermo Fisher Scientific, Inc.). Luciferase activity was evaluated 48 h after cotransfection using a dual-luciferase reporter assay system (Promega).
Statistical analysis
Data displayed as mean ± standard deviation and SPSS version 16.0 software (SPSS, Inc., Chicago, IL) was used to analyze all the data. Student's t-test and one-way analysis of variance were applied to detect the difference among groups. Survival curves were calculated using the Kaplan-Meier method and analyzed using the log-rank test. Pearson correlation coefficients were examined to assess the significant relationship between MRPL39 and miR-130 expression. p < 0.05 was identified to be statistically significant.
Results
MRPL39 expression is downregulated in GC tissues and cell lines
To investigate whether MRPL39 is associated with gastric carcinogenesis, we measured the expression of MRPL39 in GC tissues and cell lines. qRT-PCR analysis showed that the expression of MRPL39 decreased in GC tissues compared with normal gastric tissues (Fig. 1A). In addition, MRPL39 expression was also downregulated in GC cell lines (BGC823, AGS, SGC-7901) compared with the normal gastric cell line (GES-1) (Fig. 1B).
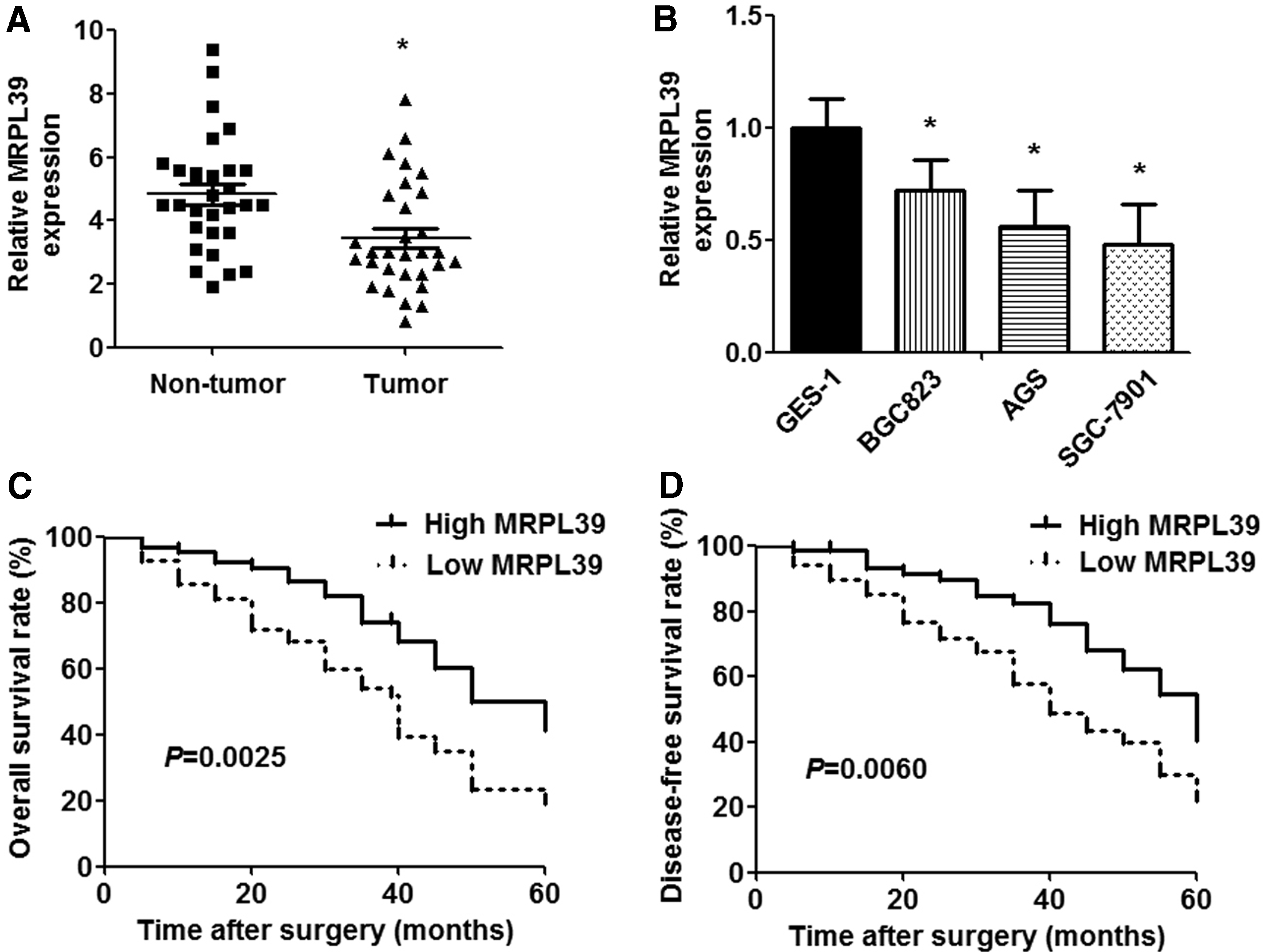
The expression and prognostic value of MRPL39 in GC specimens and cell lines.
To elucidate the clinicopathological role of MRPL39 in GC, 60 patients were divided into the MRPL39 high-expression group (n = 30) and MRPL39 low-expression group (n = 30) according to the cutoff value, which was defined as the median of the cohort. As shown in Table 1, patients with lower MRPL39 expression were significantly associated with larger carcinoma size (p = 0.0029), advanced TNM stage (p = 0.0449), and lymphatic metastasis (p = 0.0084). Furthermore, Kaplan-Meier analysis demonstrated that the patients with low MRPL39 expression revealed poor overall and disease-free survival period than those with low MRPL39 expression (p = 0.0025 and p = 0.0060, respectively; Fig. 1C, D).
Correlations Between MRPL39 Expression in Gastric Cancer and Clinical Characteristics
TNM, tumor, lymph node, metastasis.
MRPL39 inhibits cell proliferation, migration, and invasion
To clarify the function of MRPL39 in the development of GC, the MRPL39 expression plasmid was transfected in GC cells. The overexpression of MRPL39 was confirmed by qRT-PCR in transfected cells (Fig. 2A). CCK-8 assay showed that the cell growth was suppressed significantly with overexpression of MRPL39 compared with the control group (Fig. 2B). Moreover, IHC assay determined that the percentage of Ki67-labeled nuclei was lower in the MRPL39 overexpression group than that of corresponding control groups in both cell lines (Fig. 2C). In addition, cell migratory and invasive capacity also displayed an obvious downregulation in MRPL39 overexpressed cells compared with the control groups (Fig. 2D, E).

The effects of MRPL39 on GC cell proliferation, migration, and invasion. The BGC823 and SGC-7901 cells were transfected with MRPL39 expression vector (PcDNA3.1-MRPL39) and control vector (PcDNA3.1-ctrl) for 48 h.
MRPL39 acts as a molecular sponge of miR-130
To investigate the underlying molecular mechanism of MRPL39, LNCipedia.org database (https://lncipedia.org/db/search) was used to identify the potential target miRNA of MRPL39. Then, we analyzed the potential binding site by Lasergene 6.0 software (DNASTAR, Inc., Madison, WI), referred by relevant literature on lncRNA studies, establishing a lot of luciferase reporter recombinant plasmids containing the full-length MRPL39 sequence. Luciferase report assays were performed to determine the target gene. MiR-130 was found to be the potential target miRNA of MRPL39. Consequently, an inverse correlation was observed between MRPL39 level and miR-130 expression in GC tissues (Fig. 3A). The expression of miR-130 also decreased after transfection with MRPL39 plasmid in GC cell lines (Fig. 3B). Moreover, qRT-PCR assay displayed that transforming growth factor beta receptor 2 (TGFβR2) expression (the target gene of miR-130) increased significantly with the presence of MRPL39 (Fig. 3C).
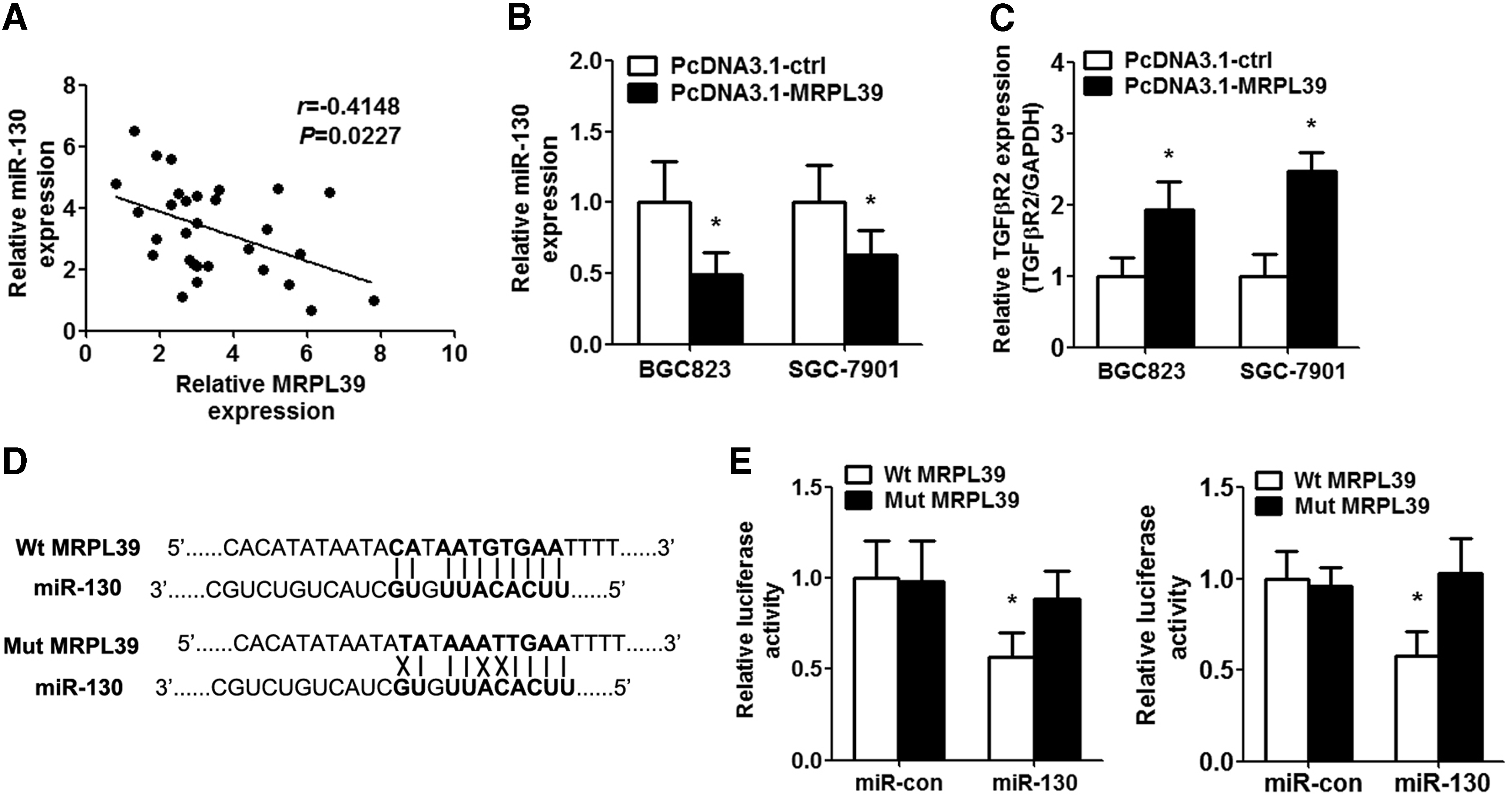
MRPL39 targeted miR-130 directly.
To clarify the direct relationship between MRPL39 and miR-130 in GC cells, we cloned the wild-type MRPL39 sequence with the potential binding site of miR-130 and the mutant MRPL39 sequence into a luciferase reporter gene system (Fig. 3D). The result demonstrated that miR-130 suppressed the luciferase activity of wild-type MRPL39 but not the luciferase activity of mutant MRPL39 in GC cell lines (Fig. 3E).
Reintroduction of miR-130 reverses MRPL39-induced effects on GC cells
To elucidate whether MRPL39 serves its role by sponging miR-130, we transfected the MRPL39 overexpressed GC cells with miR-130/miR-con mimics (Fig. 4A). According to the CCK-8 assay, cell growth was significantly promoted with the restoration of miR-130 in the BGC823 and SGC-7901 cells (Fig. 4B). Moreover, IHC assay revealed that the overexpression of miR-130 promoted Ki67-labeled nuclei in the MRPL39 overexpressed GC cells (Fig. 4C). Similarly, the enhanced migratory distance and invasive cells were also presented in the miR-130 overexpressing cells (Fig. 4D, E).
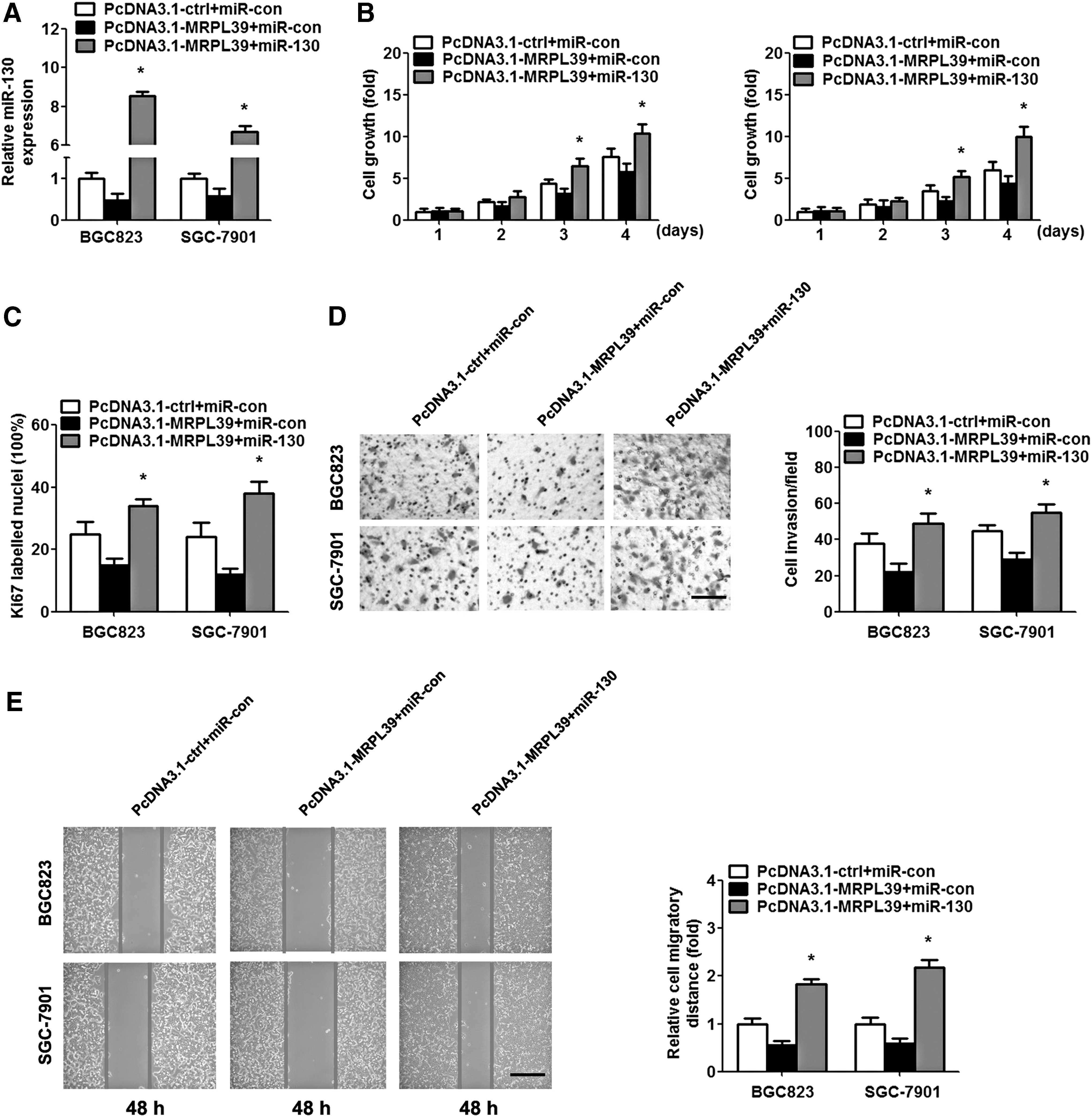
Overexpression of miR-130 reverses the MRPL39 effect on GC. BGC823 and SGC-7901 cells were cotransfected with the PcDNA3.1-MRPL39/PcDNA3.1-ctrl and miR-130/miR-con mimics for 48 h.
Discussion
In recent years, lncRNAs, the novel molecular biomarkers, have shown potential roles as biomarkers in cancers (Zhuang et al., 2014). In GC, the profile of lncRNA expression was also abnormal, for example, lncRNA XIST was reported to modulate GC progression by acting as a molecular sponge of miR-101, and lncRNA MALAT1 was revealed to promote GC development by targeting miR-1297 (Chen et al., 2016; Li et al., 2017a). Even if the extensive lncRNAs are investigated, the unsatisfactory treatments drive countless researchers to devote their attention to better the situation. In this study, we aimed to clarify the expression and molecular mechanisms of lncRNA underlying GC carcinogenesis.
LncRNAs are involved in multiple biological processes, including cell growth, cell differentiation, cell invasion, and cell apoptosis (Guttman et al., 2011; Khaitan et al., 2011). MRPL39, a 1725 bp in size, is expressed from chromosome 21. However, its biological effect has not yet been fully known, and whether MRPL39 is involved in the biological process remains to be elucidated. In our study, we observed that the MRPL39 expression level was significantly downregulated in GC tissues and BGC823 and SGC-7901 cell lines. These findings suggested that MRPL39 may serve a critical role in gastric tumorigenesis. Moreover, MRPL39 expression was significantly associated with carcinoma size, TNM stage, and lymphatic metastasis, indicating that MRPL39 may be implicated in GC development and metastasis. Besides, the patients with high expression of MRPL39 displayed longer overall and disease-free survival time, which revealed that MRPL39 might be a possible biomarker in predicting overall and disease-free survival outcomes of GC patients. These results demonstrated that MRPL39 is critical for evaluating the prognosis in GC patients. In addition, the reintroduction of MRPL39 in GC cells suppressed cell proliferation, migration, and invasion, suggesting that MRPL39 may be associated with the advanced development of GC and acts as an important biomarker in the prediction of GC progression.
Growing evidences demonstrated that lncRNAs acted as competitive endogenous RNAs (ceRNAs) for miRNAs, which function as molecular sponges to suppress miRNAs in the progression of gene expression and signaling pathways (Zhang et al., 2013; Shi et al., 2014). To clarify whether lncRNA MRPL39 is a ceRNA in GC and the underlying molecular mechanism of MRPL39 in GC, we identified miR-130 to be a direct-target gene of MRPL39 using bioinformatic databases, and we found a negative correlation between the expression of MRPL39 and miR-130 in GC tissues and GC cells. MiR-130 has been validated in modulating tumorigenesis of human nonsmall-cell lung cancer (Ye et al., 2017) and promoting cell migration and invasion in bladder cancer (Egawa et al., 2016). A previous study showed that miR-130 played an unfavorable role in proliferation and migration by targeting TGFβR2 in GC (Duan et al., 2016). Consistently, our study confirmed that miR-130 promotes GC cell proliferation and invasion (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/gtmb). Also, miR-130 was downregulated with the re-expression of MRPL39, and TGFβR2 expression was enhanced compared with the control group, suggesting that MRPL39 might inhibit cell proliferation by suppressing miR-130 and further regulating TGFβR2 expression. In addition, we found that the restoration of miR-130 abrogated the antitumor effects of MRPL39 on cell proliferation, migration, and invasion in GC cells, which indicated that MRPL39 exerted its biological effect by sponging miR-130 in GC.
Taken together, our study first demonstrated that the downregulation of MRPL39 might be a valuable biomarker for GC progression, and miR-130/TGFβR2 axis might be implicated in the MRPL39 dysregulation in GC (Fig. 5). These findings provided a novel insight in the diagnosis and prognosis of GC.

A schematic diagram illustrating the involvement of MRPL39/miR-130/TGFβR2 in GC.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.