
Abstract
Background:
Uterine leiomyoma (UL) is the most common benign smooth muscle tumor of the uterus in reproductive women. Prior studies indicated that methyl-CpG-binding domain proteins (MBDs) may be involved in the pathogenesis of UL.
Materials and Methods:
In this study, UL tissues and paired adjacent myometrium were collected from a total of 51 patients. The expression of MBD mRNAs and their cognate proteins were analyzed via quantitative polymerase chain reaction assays and western blotting, respectively. The relationships between the MBD expression levels and the patients' clinicopathologic variables were assessed using Student's t test, nonparametric tests, or Pearson χ2 methods.
Results:
Our results show that both the mRNA and protein levels of MBD2 were significantly decreased in ULs compared to the adjacent myometrium. In addition, MBD6 protein expression was also decreased significantly in UL samples when compared to the adjacent myometrium. There was, however, no significant difference on the mRNA expression of MBD6 between these two groups. Neither the mRNA nor the protein levels of the other MBD members (MBD1, MBD3, MBD4, MBD5, and MeCP2) showed any significant differences between ULs and the adjacent myometria. The decreased expression of the MBD6 protein was correlated with the tumor size of ULs.
Conclusions:
These results suggest that the dysregulated expression of MBD2 and MBD6 in ULs may play a role in their development; however, a larger sample size together with cellular functional assays should be carried out to further elucidate the precise role of MBD6 in ULs.
Introduction
Uterine leiomyomas (UL
Despite the high percentage of women affected by UL, their molecular pathogenesis remains unclear and their biologic features are poorly understood. There is accumulating evidence which shows that three main epigenetic alterations play a crucial role in modulating the gene expression in fibroid formation: DNA methylation, histone modification, and microRNAs (miRNAs) (Wei et al., 2011; Greathouse et al., 2012; Islam et al., 2013; Maekawa et al., 2013; Navarro, et al., 2012; Styer and Rueda, 2016; and Yang, et al., 2016). Two recent global DNA methylation analyses revealed that there were differential DNA methylation levels in a large number of genes between leiomyomas and their adjacent myometrium (Navarro et al., 2012; Maekawa et al., 2013). Recent studies have demonstrated that estrogen receptor alpha (ERα) expression is regulated by DNA methylation in UL formation (Asada et al., 2008; Maekawa et al., 2013). In addition, a recent study also showed that ERα expression is regulated by DNA methylation in ULs, and the expression of histone deacetylase 6 (HDAC6), a key HDAC, is associated with regulation of the expression of ERα in ULs, implicating a potential pathogenicity of increased HDAC6 expression in ULs (Wei et al., 2011).
Methyl-CpG-binding domain proteins (MBDs) function as “readers” of the epigenome via recruitment of chromatin remodeling proteins, which adjust the gene expression (Du et al., 2015; Gigek et al., 2016). Abnormal expression of MBDs has been observed in a plethora of tumor types, including cervical, breast, pancreatic, hepatocellular, and gastric carcinomas (Cui et al., 2015; Kar et al., 2014; Pontes et al., 2014; Wu et al., 2015; Xu et al., 2013). Considering the potential involvement of epigenetic aberrations in ULs (Navarro et al., 2012; Maekawa et al., 2013; Yang et al., 2016) and the presence of dysregulated expression of MBD proteins in human tumors prompted us to test whether MBDs play a role in the pathogenesis of ULs.
In this study, we evaluated the expression of MBDs (MBD1, MBD2, MBD3, MBD4, MBD5, MBD6, and methyl CpG 2 [MeCP2]) in ULs and their paired adjacent myometrium as well as a further analysis of the relationship between MBD2 and MBD6 expression and clinical pathological parameters in 51 patients with ULs.
Materials and Methods
Patients and clinical samples
Leiomyoma tissues and their adjacent myometrium (about 1-2 cm from the tumor border) were collected from 51 patients with UL, who underwent a surgical excision of the UL at the Department of Gynecology of Jiangxi Maternal and Child Health Hospital in Nanchang, China between 2014 and 2016. None of the patients had taken any hormonal medication for at least 3 months before surgery nor had history of malignancy. Serial paraffin sections were stained with hematoxylin and eosin and used for immunohistochemistry for pathologic diagnoses performed by an experienced pathologist from the hospital (Fig. 1). The available clinical information was collected. This study was approved by the Ethics Committee of Jiangxi Maternal and Child Health Hospital and an informed consent was obtained from all the participating women.
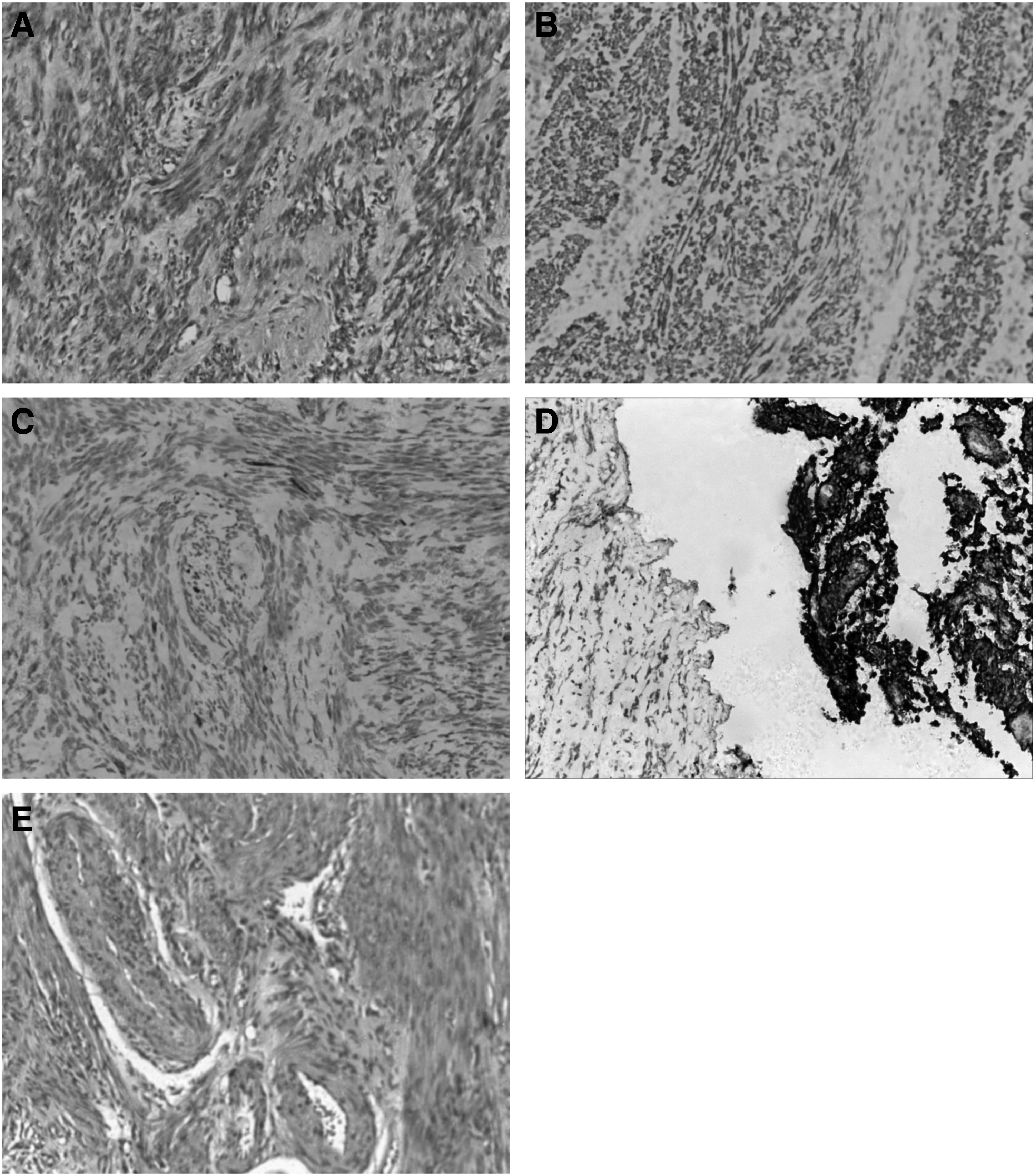
H&E staining and immunohistochemical results of ULs and individual-matched control myometrium. From
Real-time quantitative polymerase chain reaction assay
Total RNA was extracted from the tissues using the TRIzol reagent (Takara, Japan) according to the manufacturer's instructions. Before cDNA synthesis, the total RNA concentration was quantified spectrophotometrically (SmartSpec Plus Spectrophotometer; Bio-Rad Laboratory), and its integrity was evaluated by agarose gel electrophoresis. The first strand of cDNA was synthesized from 1 μg of total RNA using the PrimeScript™ RT Reagent Kit (Takara, Japan) according to the manufacturer's instructions. Each quantitative polymerase chain reaction (qPCR) was performed in a final volume of 20 μL, containing 5 μL cDNA and 15 μL of PCR master mix solution (Takara, Japan). The qPCR was performed with an initial 10 min incubation at 95°C, followed by 40 cycles of 15 s incubations at 95°C and 60 s amplification at 60°C in the ABI 7500 LightCycler (Applied Biosystems). An analysis of the data was performed using the ΔΔCt method (Livak and Schmittgen, 2001), which was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA levels. The definition of high and low mRNA expression of MBDs was dependent on the value of gene expression in the control group. Expression levels that were higher in ULs than in the paired adjacent myometrium were defined as high. Expression levels that were lower in ULs than in paired adjacent myometrium were defined as low.
Western blotting
Total protein was extracted from ULs and adjacent myometrial tissues with RIPA buffer (Applygen Technologies, Inc., Beijing, China) containing 1% (v/v) of protease inhibitor cocktail (Boster Biological Technology, Ltd., Wuhan, China). The isolated protein (30 μg of total protein) was resolved by using sodium dodcyl sulfate polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes (Solarbio Science & Technology Co., Ltd., Beijing, China), followed by blocking with 5% nonfat dry milk and incubating overnight with the primary anti-MBDs antibody at 4°C. The next day, the membranes were washed three times in Tris-buffered saline with tween and incubated with a secondary antibody conjugated with horseradish peroxidase secondary antibody (1:5000) for 1 h at room temperature. The protein bands were visualized by using the ECL Western Blotting Substrate (Thermo Fisher Scientific, IL), and the relative MBDs protein expression was normalized to GAPDH. Image J program was used to quantify the Western blotting (WB) results. The definition of high and low protein expression of MBDs was dependent on the value of the protein expression in the control group. Expression levels that were higher in ULs than in the paired adjacent myometrium were defined as high. Expression levels that were lower in ULs than in the paired adjacent myometrium were defined as low.
Statistical analyses
All the data are expressed as the mean ± standard deviation to check the data of all groups with a homogeneity test for variance and test of normality. The p-value was calculated using Student's t test or nonparametric test or Pearson χ2. The correlation between two variables was evaluated by Spearman correlation. A p-value of <0.05 was considered statistically significant. All statistical analyses were performed with the SPSS statistical package (version 19.0) or GraphPad Prism 5.0 software (GraphPad Software, Inc., San Diego, CA).
Results
Patient information
The clinicopathologic features of the 51 cases in this study are summarized in Table 1. The patients' ages ranged from 30 to 64 years (median: 47.0; mean: 47.6). Presenting symptoms included abnormal uterine bleeding (irregular vaginal bleeding, menostaxis, and menorrhagia), pelvic pain, pelvic pressure, and so on. Gross tumor measurements were available in all cases, with maximum tumor dimension ranging from 2.1 to 16.2 cm (median: 6.8; mean: 7.1).
The Clinicopathologic Features of the 51 Cases in this Study
SD, standard deviation.
The mRNA expression of MBDs in ULs
The primer sequences for the qPCR assays are listed in Table 2. The mRNA expression levels of MBD1, MBD2, MBD3, MBD4, MBD5, MBD6, and MeCP2 were examined in 51 patients who were diagnosed with ULs and their matched adjacent myometriums (Fig. 2). Our results indicate that the mRNA expression of MBD2 decreases significantly in ULs compared to their adjacent myometrium (95% confidence interval [CI]: −3.529 to −0.3940; p = 0.015); while mRNA levels of all other MBD members (MBD1, MBD3, MBD4, MBD5, MBD6, and MeCP2) showed no significant differences between ULs and their adjacent myometriums (p > 0.05).

The expression of MBDs members mRNA in ULs and individual-matched control myometrium. From
Forward and Reverse Primers Used for Quantitative Polymerase Chain Reaction
The protein expression of MBDs in ULs
We applied a WB assay to detect the protein expression of MBDs between ULs and their adjacent myometrium with antibodies that specifically and individually recognize MBD1, MBD2, MBD3, MBD4, MBD5, MBD6, and MeCP2 protein expressions (Fig. 3). All the antibodies that were used in this study are listed in Table 3. Our results show that the protein expression of MBD2 decreases in ULs when compared to their adjacent myometriums (95% CI: −0.9928 to −0.07873; p = 0.022). In addition, we also found that the expression of MBD6 was significantly reduced in ULs when compared to normal controls (95% CI: −0.3562 to −0.03765; p = 0.018). Furthermore, the protein levels in all the other MBD members (MBD1, MBD3, MBD4, MBD5, and MeCP2) showed no significant difference between ULs and their adjacent control myometriums (p > 0.05).
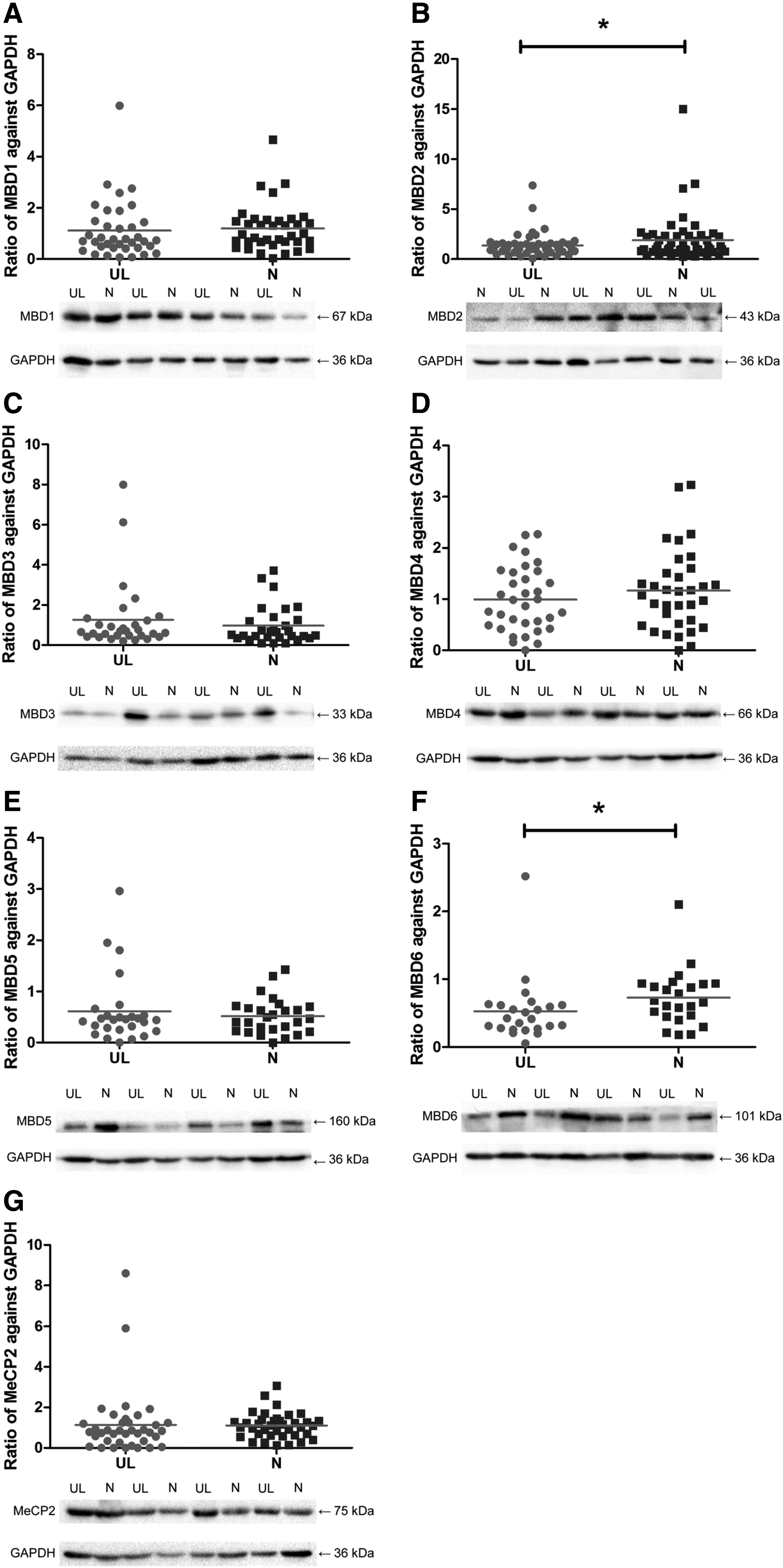
Representative picture of WB results of MBDs expression in ULs and individual-matched control myometrium. From
All the Antibodies that Were Used in this Study
The relationship between MBD2 and MBD6 expression and clinical pathological parameters
Table 4 displays the relationship between MBD2 and MBD6 expression and their clinical pathological parameters for 51 patients with ULs. These results show that decreased expression of the MBD6 protein is correlated with the tumor size of ULs (χ2 = 5.445, p = 0.038; odds ratio: 2.5, 95% CI: 0.791-7.898; r = −0.476, p = 0.019). There was no positive correlation between the expression of the MBD2 protein and the clinicopathologic data (p > 0.05).
The Relationship Between MBD2 and MBD6 Expression and Clinical Pathological Parameters in 51 Patients with ULs
p < 0.05.
UL, uterine leiomyoma.
Discussion
The MBDs are a class of proteins that specifically combine with the MBD domain, which are primary candidates for the readout of DNA methylation as they recruit chromatin remodelers, HDACs, and methylases to methylated DNA associated with gene repression (Du et al., 2015). Currently, seven MBDs have been identified, including MeCP2, MBD1, MBD2, MBD3, MBD4, MBD5, and MBD6 (Li et al., 2015). Dysregulated expression of MBD members have been widely observed in multiple human cancers (Cui et al., 2015; Kar et al., 2014; Pontes et al., 2014; Wu et al., 2015; Xu et al., 2013). Recent studies have confirmed that aberrant DNA methylation is implicated in the pathogenesis of ULs (Jankowska et al., 2015; Yang et al., 2016). Given that MBDs are primary candidates for the readout of DNA methylation, in this study, we investigated their mRNA and protein expression levels in ULs compared with adjacent normal myometrial tissues in 51 patients with ULs.
Our results show that both the mRNA and protein expression of MBD2 and the protein expression of MBD6 decreased in ULs when compared to adjacent myometrium. The mRNA and protein levels of the other MBDs (MBD1, MBD3, MBD4, MBD5, and MeCP2) showed no significant difference between these two groups. Our results indicate that the decreased expression of the MBD6 protein is correlated with tumor size of ULs. These results suggest that MBD2 and MBD6 might be involved in the pathogenesis of ULs.
The MBD2 protein is dispensable for animal viability, but it is implicated in the genesis of many tumors, including lung and colon cancers (Campbell et al., 2004), breast cancer (Alvarado et al., 2013), glioblastomas (Zhu et al., 2011), hilar cholangiocarcinomas (Chen et al., 2011), and gastrointestinal stromal tumors (He et al., 2013). This study is the first report to analyze the MBD2 expression in ULs. Although our results showed the mRNA and protein expression of MBD2 decreased in ULs, the underlying molecular mechanisms remain unknown. Prior studies have reported reduced MBD2 levels in neoplastic samples when compared with adjacent nonneoplastic samples, moreover, the neoplastic tissues exhibited higher MBD2 promoter methylation than in normal tissues, Moreover, a weak correlation between gene expression and methylation was observed (Pontes et al., 2014).
Another study showed that arginine methylation of MBD2 repressed the expression of the MBD2 protein, and then reduced the MBD2-methyl-DNA complex formation and impaired the transcriptional repression function of MBD2 (Tan and Nakielny, 2006). These findings further support the hypothesis that the promoter methylation of MBD2 might play an important role in regulating MBD2 protein expression, and that MBD2 dysregulation might lead to expression changes of downstream genes. Therefore, further studies exploring the promoter methylation status of the MBD2 gene between ULs and control myometrium are needed to elucidate whether the promoter methylation of MBD2 is responsible for the decreased mRNA and protein expression of MBD2 in ULs.
MBD6 is also a member of the MBD family of proteins, but is poorly characterized. It has no other annotated domains in addition to the MBD domain and it localizes to heterochromatin foci in mouse nuclei in a DNA methylation independent manner (Laget et al., 2010). In this study, MBD6 mRNA levels were shown to be similar in both normal myometrium and leiomyomas, but protein levels were decreased in UL tissue. We speculate that the differential expression of the MBD6 protein between the leiomyomas and their normal myometrium might be attributed to differential posttranscriptional or posttranslation modification. It has been reported that mutational and expressional alterations of MBD6 with respect to the microsatellite instability status in Gastric and Colorectal Cancers (Choi et al., 2015). MBD6 has been found to interact directly with the human polycomb deubiquitinase complex, which catalyzes deubiquitination of H2AK119 (Baymaz et al., 2014). Importantly, a recent study showed MBD6 to be an octamer-binding transcription factor 4 (Oct4) target gene in adipose tissue stem cells and was regulated by Ago2 via an interaction with Oct4, which altered self-renewal and gene expression in hATSCs (Jung et al., 2013). Therefore, we hypothesize that MBD6 as an Oct4 target gene may play a crucial role in the pathogenesis of ULs through the Ago2-Oct4 pathway by promoting cell proliferation. Next, we will explore the relationship between MBD6 and Oct4 in ULs and further mechanistic studies will be needed to validate this speculation.
In conclusion, our findings of the dysregulated expression of MBD2 and MBD6 in ULs suggest that these genes might be associated with the development of UL. Specifically, our study shows for the first time that the decreased expression of the MBD6 protein is correlated with tumor size of ULs and will likely provide new opportunities to develop an efficient therapeutic approach. This study is, however, only a primary exploration. We have not investigated the underlying mechanisms of the decreased expression of MBD2 and MBD6 in ULs. Accumulating evidence demonstrates that some intrinsic abnormalities of the myometrium, abnormal myometrial receptors for estrogen, and hormonal changes or altered responses to ischemic damage during the menstrual period may be responsible for the initiation of (epi)genetic changes found in uterine myomas (Yang et al., 2016; Laganà et al., 2017). We plan to further elucidate the relative underlying mechanisms based on a larger sample size of ULs and cellular functional assays.
Footnotes
Acknowledgments
We thank all participants involved in this study, and we extremely acknowledge Dr. Chunxiao Zhou for his recommendations in revising this article. This study was supported by grants from Jiangxi Provincial Department of Science and Technology (No. 20181BBG78016) and Health and Family Planning Commission of Jiangxi Province (No.20185391).
Authors' Contributions
Conceived and designed the experiments: O.-P.H., Y.Z.
Performed the experiments: M.-R.L., Y.Z., J.-W.Z.
Analyzed the data: M.-R.L., Y.L.
Contributed reagents/materials/analysis tools: B.-C.Y., Z.-Y.Z., F.-Y.L.
Wrote the article: M.-R.L.
Gathered specimens for experiment: S.-Y.Z.
Edited the article: M.-R.L.
Author Disclosure Statement
No competing financial interests exist.