
Abstract
Background:
The gene programmed cell death 5 (PDCD5) has recently been characterized as a tumor suppressor gene and is believed to be an important prognostic cancer marker; it is frequently involved in neoplastic transformation and apoptosis of tumor cells. Several studies have demonstrated a decrease or loss of expression of PDCD5 in certain tumors. However, the relevance of PDCD5 expression in human osteoclastoma and its clinicopathological significance have not been extensively studied.
Methods:
The aim of this study was to explore the relative transcriptional and translational expression levels of PDCD5 in 79 osteoclastoma samples using multi-modal methods of analysis.
Results:
Our findings showed that 52% (15/29) of osteoclastoma cases exhibited reduced PDCD5 expression at the transcriptional level, and 56% (44/79) exhibited lower PDCD5 expression at the protein level, when compared with nontumor tissue. In addition, the statistical significance of the altered PDCD5 protein expression was examined using the Campanacci grading system for osteoclastoma. More importantly, the decreased expression at the translational level was observed to have a negative association with the Ki-67 staining index.
Conclusion:
Based on these findings, abnormal PDCD5 expression might be an important biomarker in human osteoclastoma and may contribute to tumor progression and malignant cell proliferation.
Introduction
Programmed cell death 5 (PDCD5), first discovered in 1999, has been reported in several organisms ranging from yeast to mice; PDCD5 is known to induce or strengthen the process of cell apoptosis caused by serum or growth factor deprivation (Liu et al., 1999). The PDCD5 protein contains a core structural domain and two separated terminal regions and is extensively expressed in different types of cells and tissues (Liu et al., 2004; Wang et al., 2016). This suggests that it may have an important effect on pathophysiological processes and disease progression (Li et al., 2016). PDCD5 may inhibit the growth of tumor cells by regulating proliferation, producing cycle arrest, and inducing apoptosis (Li et al., 2018).
Multiple studies recently reported downregulation of PDCD5 expression in a variety of tumor types, including lung, breast, colorectal, prostate and gastric carcinoma, high-grade astrocytic glioma, acute or chronic myelogenous leukemia, and hepatocellular, renal clear cell, ovarian, and bladder carcinoma (Hedenfalk et al., 2001; Ruan et al., 2006, 2007; Spinola et al., 2006; Tan et al., 2006; Xiong et al., 2006; Yang et al., 2006; Li et al., 2008; Du et al., 2009; Yin et al., 2010; Zhang et al., 2011; Fu et al., 2013). It has been demonstrated that PDCD5 fragments may suppress tumorigenicity by affecting the MEK/ERK molecular signaling pathway in malignant bone tumors (Han et al., 2012). In addition, decreased levels of PDCD5 transcript and protein were detected in chondrosarcoma tissues, and it was found to be a potential target gene for chondrosarcoma treatment (Chen et al., 2010). The results described earlier suggest that PDCD5 may play a crucial role in the malignant progression of bone tumors, making it a potential therapeutic target for such tumors.
Osteoclastoma is a benign and typically local aggressive tumor with unpredictable biological behavior (Campanacci et al., 1987). It makes up 5% of most of the idiopathic bone tumors. In the East and in Southeast Asia osteoclastoma patients are predominantly male, whereas in Western countries they are more often female. Although rarely lethal, 8-62% of osteoclastoma cases result in local recurrence of the tumor after surgical treatment accompanied by radiotherapy, whereas ∼5% of cases produce pulmonary metastases and 2% result in the development of spontaneous malignant transformations.
In the past few decades, several researchers have concentrated on different aspects of the molecular and cellular regulation of osteoclastoma to decrease recurrence rates and improve clinical outcomes. However, biological markers that can predict the degree of recurrence or malignant transformation have yet to be identified. By comparison, it has been demonstrated that gene expression profiling is the most valuable in identifying critical molecules that influence tumor malignancy. Therefore, the purpose of this study was to find new biological molecules involved in the malignant progression of osteoclastoma, which could improve patients' prognosis and help predict the degree of malignancy in these tumors. Although several studies have reported that PDCD5 has an inhibitory effect on the progression of osteocarcinoma, the expression of PDCD5 and its clinicopathological significance in human osteoclastoma remain largely unknown.
In this study, we examined PDCD5 expression at the transcriptional and protein levels in osteoclastoma tumors using real-time reverse transcription-polymerase chain reaction (RT-PCR) analysis, Western blotting, and immunohistochemistry (IHC). We compared our results with similar analyses conducted in adjacent nontumor tissue, in an attempt to determine whether there is a correlation between abnormal PDCD5 expression, malignant biological behavior, and its corresponding clinicopathological characteristics.
Methods
Patient selection and tissue collection
Tumor specimens, including 29 fresh surgical excision specimens and 50 paraffin samples, were collected from 79 patients with osteoclastoma, who were undergoing operation treatment at the department of bone surgery, Shandong Provincial Hospital, Shandong University (Jinan, China). Specimens retrieved fresh after surgical operation were immediately frozen in liquid nitrogen and stored at −80°C in an ultracold storage freezer, therefore were not paraffin embedded. All of the nontumor tissues were obtained from areas adjacent to the tumors. All the osteoclastoma patients received extended curettage, without adjuvant radiotherapy or chemotherapy. The clinical parameters assessed in the patients and adjacent control samples included gender, age, tumor size, tumor location, Campanacci grade, and recurrence (Table 1). The biological behavior and malignant progression of osteoclastoma were assessed using the Campanacci method (Campanacci et al., 1987) and the Enneking staging system (Enneking et al., 2003). Informed consent was obtained from all subjects involved in our studies, and their tissue samples were used only for research purposes. The current research was approved by Shandong Provincial Hospital Institutional Review Board.
The Demographical and Clinical Characteristics in Patients with Primary Osteoclastoma and Adjacent Nontumor Tissues
SD, standard deviation.
RNA extraction, cDNA synthesis, and real-time RT-PCR
RNA of frozen tissue samples from the fresh tumor and adjacent nontumor region was isolated through a modified extraction method using TRIzol reagent (TIANGEN, Beijing, China) as described in previous studies (Gao et al., 2012, 2018). The FastQuant RT Kit was used to synthesize cDNA on the basis of the manufacturer's instructions (TIANGEN). PCR was carried out in a 20 μL system consisting of 1 μL cDNA, 2 μL primers (PDCD5 and β-actin), 10 μL SYBR® PrimeScript Master Mix (CWBIO, Beijing, China), and 7 μL ddH2O. The sequences of the PDCD5 primers used are forward primer sequence 5′-TCG TCA TCT TCA TCA GAG TCC A-3′ and reverse primer sequence 5′-GTT CTG GAT CAG TCG GCC C-3′. Real-time RT-PCR conditions were as follows: hold denaturation at 95°C for 10 min, PCR Stage (40 cycles) 95°C (15 s), annealing at 60°C (1 min), and melt curve stage 95°C (15 s), 60°C (1 min), 95°C (15 s). Each experiment was repeated in triplicate and analyzed using the 2−ΔΔCq method. All samples were normalized to β-actin.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis and Western blot analysis
Tissues from the fresh tumor and adjacent nontumor region were promptly frozen in liquid nitrogen. Total protein was resuspended using RIPA buffer (Haoran Biological Technology Co., Ltd., Shanghai, China). Subsequently, protein concentrations were determined using a Bradford method kit (Thermo Fisher Scientific, Rockford, IL). Forty micrograms per lane of each protein sample was loaded and separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then shifted onto polyvinylidene fluoride membranes (Haoran Biological Technology Co., Ltd.). The membranes were then blocked in 5% skimmed milk in TBS-T containing 50 mM Tris, 150 mM NaCl, and 0.1% Tween 20, for 1 h at room temperature. This was followed by incubation with specific primary antibodies, including rabbit polyclonal against PDCD5 (dilution 1:500, no. ab83958; Abcam, Inc., Cambridge, MA), in TBS-T containing 5% milk, overnight at 4°C. The membranes were then incubated in horseradish peroxidase-conjugated secondary antibodies (dilution 1:2,000, no. 7074; Cell Signaling Technology, Danvers, MA) for 1 h at 37°C. After extensive washing with PBS-T, the proteins were visualized and imaged using enhanced chemiluminescence detection.
Immunohistochemistry
Protein expression was further evaluated and analyzed by a standard IHC staining method for the 29 frozen and 50 paraffin-embedded tissue samples, as well as the 12 control adjacent nontumor samples. Four to 6 μm paraffin-embedded tissue sections underwent IHC staining followed by microwave-based antigen retrieval. All of the slides were incubated with anti-PDCD5 antibody (dilution 1:1,000, No. 75430; Abcam) and anti-Ki-67 antibody (dilution 1:400; Cell Signaling Technology, Danvers, MA) for 1 h in a wet chamber at 37°C. The sections were then probed using an HRP-streptavidin DAB kit (GeneTec, GK600505, Shanghai, China). The results of the IHC-staining were independently assessed and analyzed by experienced pathologists blinded to the origin of the tumor tissues. The percentage area and intensity of PDCD5 staining were scored from grades 0 to 5 as follows: 0 (−), 1 (+), 2 (++), 3 (+++), 4 (++++) and 5 (+++++). The cutoff norms for the sum of the scores were described as follows: 0 for no expression, score 1-3 for weak low expression, and score 4-5 for moderate to strong positivity (high protein expression). The labeling indexes of Ki-67 were measured using a semiquantitative counting standard and defined as the ratio of immunoreactive positive tumor cells to the total number of counted cells.
Data analysis
The data were compiled on Excel 2007 and analyzed using the SPSS v22.0 software (SPSS, Inc., Chicago, IL). The measurements were expressed as mean ± standard error. All results were analyzed using one-way ANOVA, two-tailed Student's t-test, and Pearson's coefficient tests. Spearman's statistics and multivariate regression analyses were employed to evaluate the associative relationships, which were considered to be statistically significant at p < 0.05.
Results
Identification of diminished PDCD5 expression at the transcriptional level in primary osteoclastoma tissues by RT-qPCR
To confirm whether there were abnormalities in PDCD5 expression at different levels, total RNA from each fresh frozen sample was isolated to measure relative gene transcript levels. Twelve specimens of adjacent nontumor tissues were found to have high or moderate levels of PDCD5 mRNA expression, whereas 52% (15/29) of the frozen osteoclastoma samples exhibited decreased PDCD5 expression at the transcriptional level (Fig. 1A). These data indicated that there was a significant difference between PDCD5 mRNA expression in osteoclastoma group relative to the control nontumor group tissues (p < 0.01; Fig. 1B).
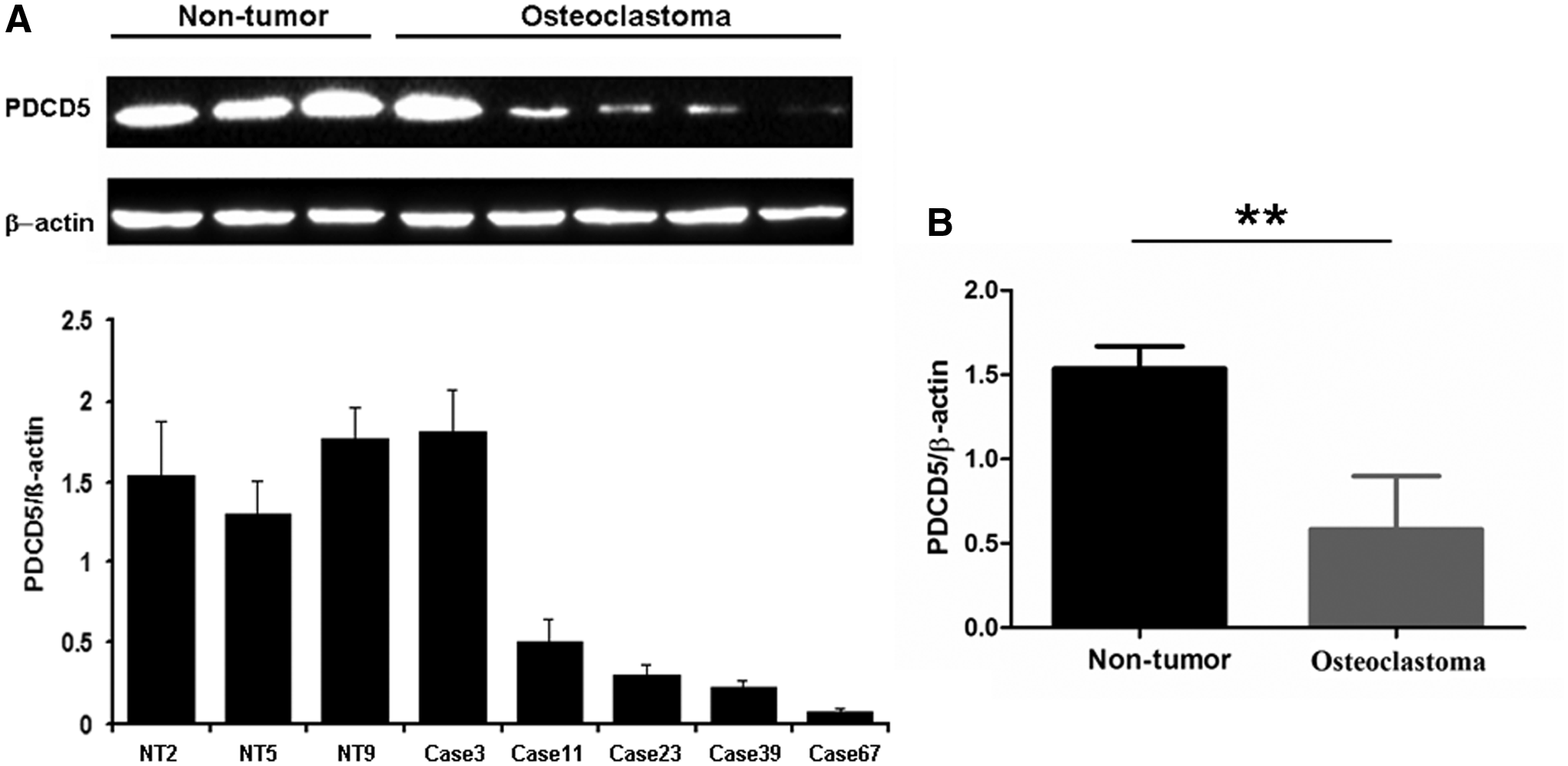
Verification of decreased PDCD5 expression at the transcriptional level in primary osteoclastoma.
Establishment of downregulated PDCD5 protein expression in primary osteoclastoma tissues by Western blotting and IHC
Western blotting and IHC, using specific antibodies against the PDCD5 protein, were performed to further confirm the RT-qPCR results. For the frozen tumor samples, Western blot analysis demonstrated that the expression of PDCD5 was significantly reduced at the protein level in tumor samples when compared with the control adjacent nontumor group (p < 0.01; Fig. 2). In addition, IHC detection further confirmed that PDCD5 expression at the translational level was lower in 56% (44/79) of primary osteoclastoma specimens, including both the fresh surgical excision specimens and paraffin-embedded tissues. As shown in Figure 3A, a positive staining of PDCD5 was observed in the slices of nontumor tissues adjacent to the tumors. However, there was weak staining in primary osteoclastoma tissues (Fig. 3B-E). In brief, a comparison of PDCD5 expression at the protein level revealed a significant difference between nontumor and tumor tissues (p < 0.01; Fig. 4).
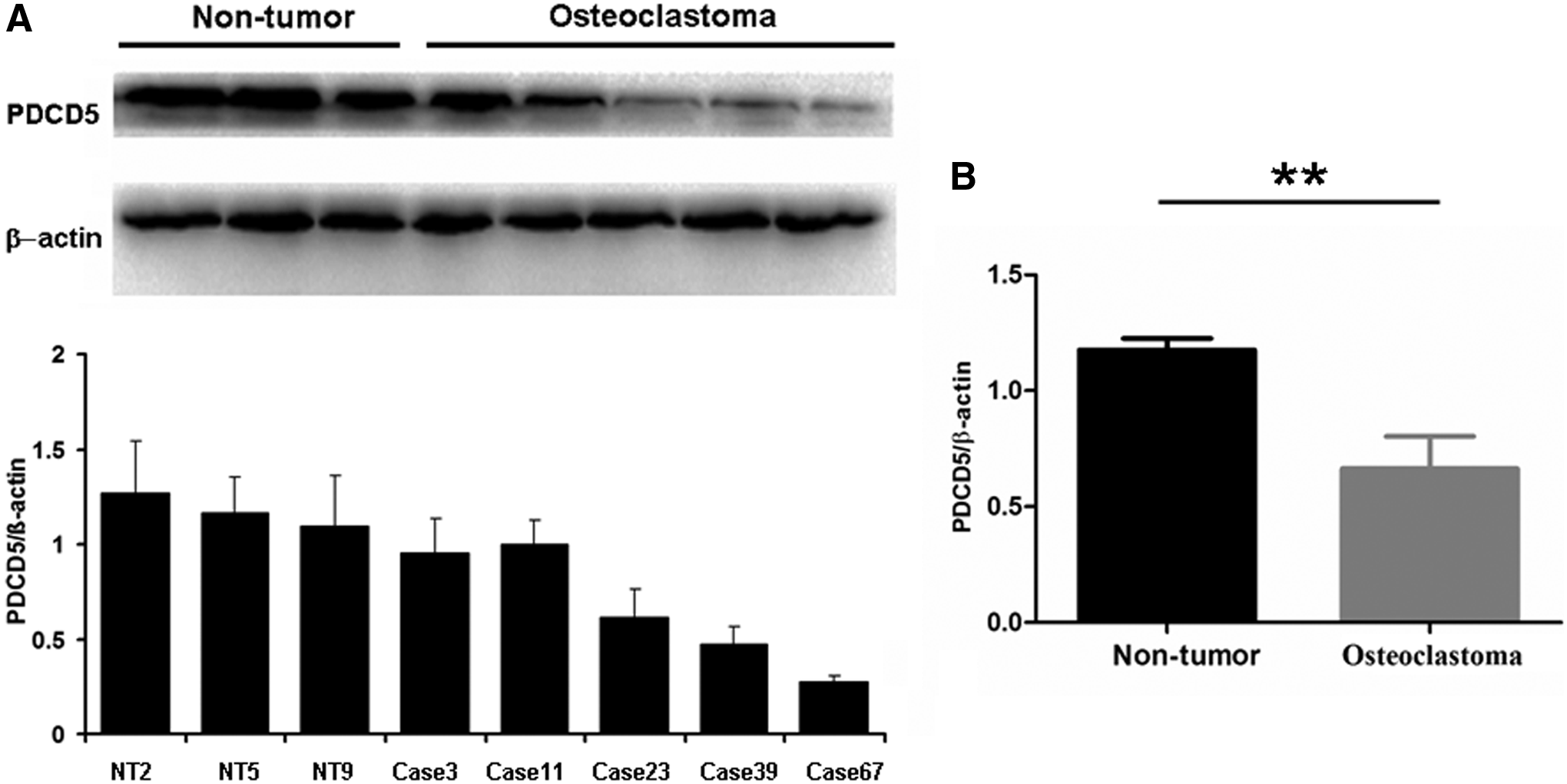
Validation of loading controls for PDCD5 protein through Western blot in primary osteoclastoma.
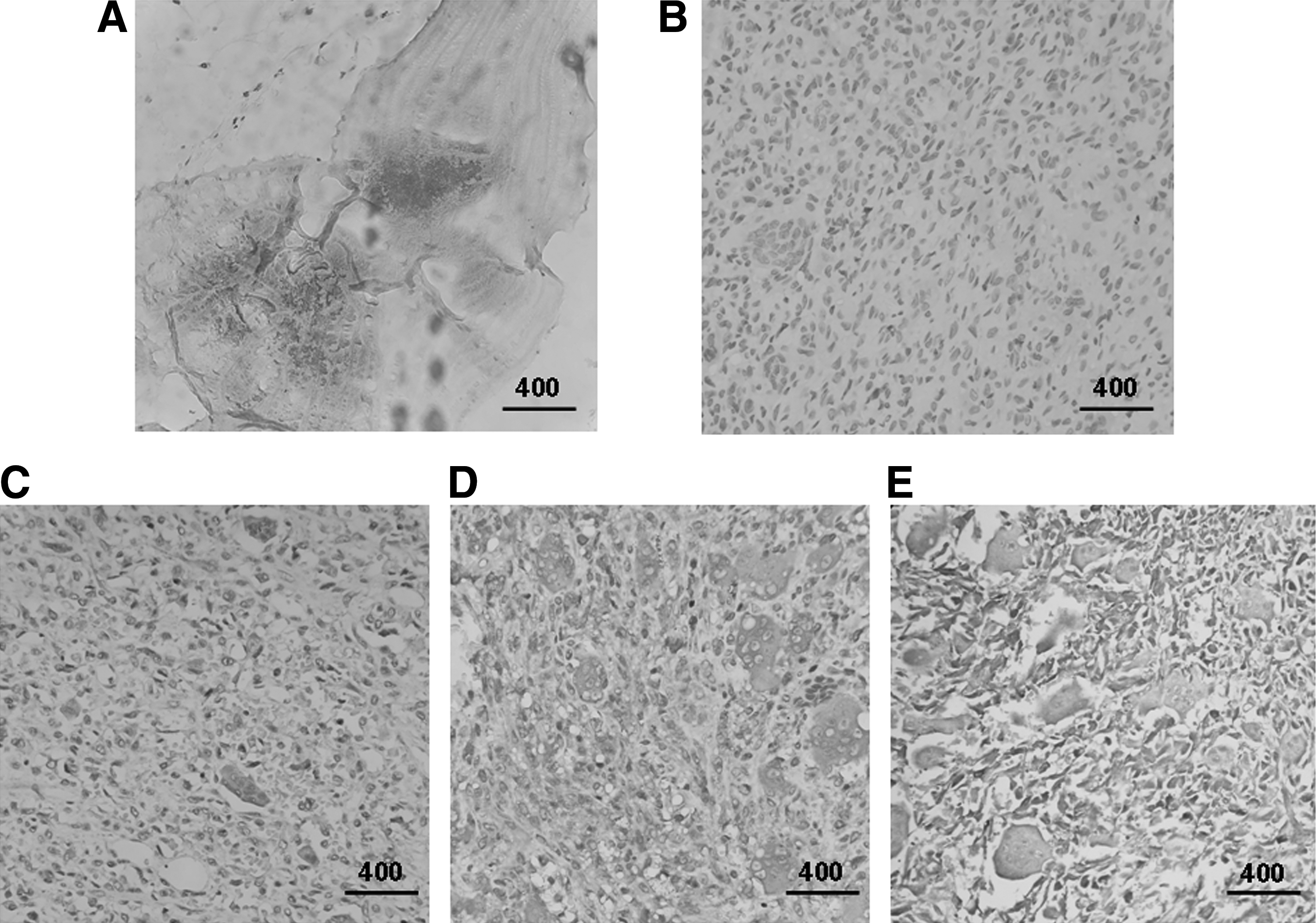
Detection of PDCD5 protein expression by IHC analysis in all of the primary osteoclastoma and control nontumor tissues (scale bar denotes 400 μm).

Statistical analysis of PDCD5 protein expression in 79 tumor cases and 12 adjacent nontumor tissues. IHC index of PDCD5 expression in osteoclastoma specimens was significantly lower than that in control nontumor tissues. **p < 0.01.
Association of clinical parameters with reduced PDCD5 protein expression in primary osteoclastoma tissues
To evaluate the clinical relevance of PDCD5 in osteoclastoma, we analyzed the relationship of PDCD5 expression at the translational level, as determined by IHC, with the clinicopathological characteristics of the patients. No significant associations were observed between the level of PDCD5 protein expression and gender, age, tumor size, location, or recurrence (p > 0.05). Interestingly, there was a statistical association between PDCD5 protein expression and the Campanacci grade (p = 0.0455), indicating that osteoclastoma might inhibit PDCD5 expression to accelerate malignant transformation.
Inhibitory effect of PDCD5 expression on malignant proliferation of primary osteoclastoma
To investigate whether PDCD5 expression affects the malignant proliferation of osteoclastoma, Ki-67 detection through IHC analysis was utilized to study the inhibitory impact of PDCD5 at the protein level (Fig. 5). Ki-67 plays a key role in the malignant transformation of osteoclastoma (Antal et al., 1999). In our study, Ki-67 expression was demonstrated to be lower in the group with high PDCD5 expression in the osteoclastoma samples relative to the samples with low PDCD5 expression (p < 0.001; Fig. 6A). As shown in Figure 6B, the Ki-67 labeling index was used to identify a negative correlation between PDCD5 protein expression and malignant proliferation of primary osteoclastoma (r = −0.5011, p < 0.001), suggesting that PDCD5 might have an inhibitory effect.

Different levels of the Ki-67 protein in primary osteoclastoma analyzed by IHC (scale bar: 400 μm).
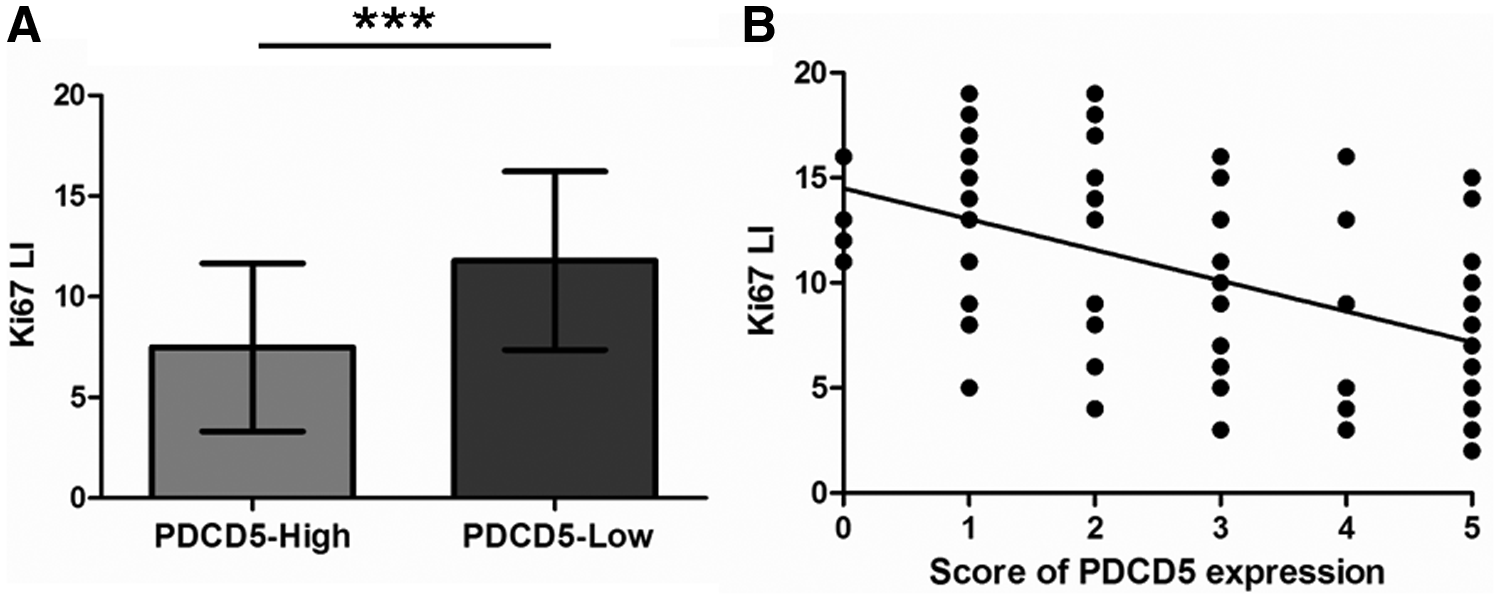
Inhibitory effect of PDCD5 protein expression on malignant proliferation of primary osteoclastoma.
Discussion
In this study, we found that PDCD5 expression was markedly downregulated in osteoclastoma tissues at the transcriptional and translational level when compared with adjacent nontumor tissues (Figs. 1-3). Furthermore, decreased protein expression was closely associated with the Campanacci grade of osteoclastoma (Table 2). In particular, the Ki-67 labeling index identified a negative correlation between PDCD5 protein expression in primary osteoclastoma and tumor malignancy. These results suggest that PDCD5 may be a key molecular marker for tumor progression.
The Association Between PDCD5 Expression at Translational Level and Clinicopathological Characteristics in Patients with Osteoclastoma
p < 0.05; ***p < 0.001.
PDCD5 expression in human osteoclastoma.
PDCD5, programmed cell death 5.
To identify possible abnormalities in PDCD5 expression in primary osteoclastoma, we first investigated PDCD5 expression at the transcriptional level by collecting fresh-frozen tissue samples. We observed that 52% of these tissue samples exhibited lower expression levels relative to the adjacent nontumor tissues. However, the precise regulatory mechanisms underlying the abnormal PDCD5 mRNA expression pattern observed in tumors have yet to be fully established. In recent years, multiple epigenetic studies demonstrated that DNA hypermethylation may cause inactivation of tumor suppressor genes to promote tumor development (Baylin and Herman, 2000; Ehrlich, 2002; Esteller, 2002). Human research on breast epithelial cells, using DNA methylation analysis, found that PDCD5 mRNA expression was downregulated after exposure to bisphenol A (Fernandez et al., 2012). Our previous studies also demonstrated that the methylation of 5′ CpG Islands contributed to the silencing of the PDCD4 gene at the transcriptional level in glioma tissues, and that treatment of glioma cells with a methyltransferase inhibitor could reverse this abnormal influence (Gao et al., 2009). Therefore, we hypothesized that abnormalities in PDCD5 expression at the transcriptional level in osteoclastoma may be associated with DNA methylation, through post-transcriptional regulation. However, the possible mechanisms mediating the effects of DNA methylation on PDCD5 mRNA expression require further investigation.
Osteoclastoma are benign bone tumors with a tendency for aggravated effects local to the tumor region. However, lung metastasis may occur in 5% of these cases (Errani et al., 2010). The primary therapeutic option for osteoclastoma is surgical treatment, depending on the extent of the disease, because intralesional curettage after incomplete resection is associated with high recurrence rates (up to 50%) (Arbeitsgemeinschaft et al., 2008; Skubitz, 2014). There are several hurdles in the available medical treatments for giant cell tumors, for example, calcitonin therapy, where no progress has been made in the double-blinded case studies for this treatment (de Lange et al., 2006). In addition, steroid injections have caused adverse effects in patients with central giant cell granulomas (EI Hadidi et al., 2015). Furthermore, biological modulation therapy and gene suppressor techniques are expensive and their use must be justified to treat these types of tumors. The recurrence rate after operation is correlated with surgical treatment and the Campanacci grade (Campanacci et al., 1987). Therefore, the statistical relationship between decreased expression of the PDCD5 protein and the pathological features of osteoclastoma was further evaluated for clinical significance, with specific focus on the association between PDCD5 protein expression and Campanacci grade. However, no correlation with recurrence rate was identified. It had been reported that the risk of recurrence for osteoclastoma was correlated with Campanacci grade (Prosser et al., 2005). However, other studies have suggested that surgical treatment was the main factor affecting the recurrence of tumors postoperation, and that Campanacci grade did not correlate with osteoclastoma recurrence (Balke et al., 2008). Based on the short duration of our study, there is no survival analysis of the follow-up data showing that decreased PDCD5 protein expression induced the biological behavior of osteoclastoma. Regarding next steps, Kaplan-Meier analysis and log-rank comparison should be carried out to analyze whether PDCD5 expression status is closely associated with the prognosis of osteoclastoma patients after operation. Therefore, PDCD5 may affect the malignant progression of osteoclastoma, but appears to have no effect on their recurrence. The possible mechanisms underlying the effects of PDCD5 on Campanacci grade remain to be further elucidated.
A study using a stable overexpressing A431 cell model demonstrated that PDCD5 significantly inhibits cell multiplication by promoting G2/M cycle arrest, to induce apoptosis (Li et al., 2018). Therefore, PDCD5 might lead to the apoptosis of osteoclastoma cells. Our study verified that Ki-67 staining, an index of the malignant proliferation of osteoclastoma cells, may be suppressed by PDCD5 and a negative relationship was identified between these factors. Osteoclastomas are characterized by growth of mononuclear stromal cells with a homogeneous presence of numerous multinucleated giant cells (Sobti et al., 2016). Neoplastic stromal cell (NSC) apoptosis has been shown to increase in multiple studies investigating the suppression of NSC-triggered giant cell malignant proliferation in osteoclastoma (Xu et al., 2016). PDCD5 may promote apoptosis of these cells to inhibit osteoclastoma proliferation and differentiation, which requires further experimental study.
In summary, we collectively found that PDCD5 might have an important effect on the malignant progression and transformation of osteoclastoma. Therefore, restoring PDCD5 expression may prove to be an effective treatment strategy for primary osteoclastoma. However, the precise molecular mechanisms underlying adjustment and control of cellular apoptosis and cell phase differentiation by PDCD5 in osteoclastoma require further investigation.
Footnotes
Acknowledgments
We thank Dr. Peng Su (Shandong University Qilu Hospital, Jinan, China) for providing valuable technical support in this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported by research grants from the National Natural Science Foundation of China (81701404 and 81470403), the Key Research and Development Plan and Natural Science Foundation of Shandong Province (2016GSF201141, 2017GSF18165, 2018GGE-2018GSF, ZR2017BH074, and ZR2016HM01) and the China Postdoctoral Science Foundation (2017M610431).