
Abstract
Objective:
Hypertension is a disease caused by both genetic and environmental factors. In the present study, we analyzed the association of the lung cancer adenocarcinoma metastasis-associated gene 1 (MALAT1) gene rs664589 locus single nucleotide polymorphism (SNP) with the risk of essential hypertension and explored its possible mechanisms.
Materials and Methods:
We analyzed the genotype of the MALAT1 gene rs664589 locus in 260 hypertensive patients and 260 healthy controls. The levels of plasma long-chain noncoding RNA (lncRNA) MALAT1, hsa-miR-539-3p, and hsa-miR-485-3p were determined by reverse transcription real-time quantitative PCR (qRT-PCR). The effects of MALAT1 on the expression levels of hsa-miR-539-3p, hsa-miR-485-3p, and bone morphogenetic protein receptor type 2 (BMPR2) were detected by transfection of human umbilical vein endothelial cells.
Results:
The risk of hypertension in subjects carrying the G allele of the MALAT1 gene rs664589 locus was 1.33 times higher than the C allele carriers (95% confidence interval [CI]: 1.15-1.51, p < 0.001). This MALAT1 gene rs664589 locus SNP was significantly associated with the risk of hypertension only in men, subjects with obesity, a history of smoking, and a history of drinking (p < 0.05). lncRNA MALAT1 was downregulated in the plasma of hypertensive patients. In addition, the level of plasma lncRNA MALAT1 was significantly lower in the G allele carriers of the MALAT1 gene than in the C allele carriers (p < 0.001). The lncRNA MALAT1 inhibited the expression of hsa-miR-539-3p and hsa-miR-485-3p and promoted the expression of the BMPR2 protein.
Conclusion:
The G allele of MALAT1 gene rs664589 locus SNP is associated with an increased risk of hypertension. In subjects carrying the G allele, the expression of lncRNA MALAT1 in plasma is significantly decreased, resulting in an abnormally high expression of hsa-miR-539-3p and hsa-miR-485-3p, and inhibition of BMPR2 expression, which might be associated with hypertension; however, further studies in animal models are needed to confirm this hypothesis.
Introduction
Recently, the prevalence of hypertension in China is gradually increasing. The awareness rate, treatment rate, and control rate of hypertensive patients have increased significantly in recent years; however, overall, they are still at low levels: 51.6%, 45.8%, and 16.8%, respectively (Liu and Writing Group of 2010 Chinese Guidelines for the Management of Hypertension, 2011). Prior studies have shown that there is a continuous, independent, and direct positive correlation between blood pressure levels and cardiovascular risk, of which stroke is the most important complication of hypertension in China. Hence, it is extraordinarily important in controlling cardiovascular diseases to control the occurrence and development of hypertension (Houston, 2011; Cappuccio and Miller, 2016; Patel et al., 2017).
Essential hypertension, a polygenic genetic disease, is caused by a combination of genetic and environmental factors, and its pathogenesis has not yet been fully clarified (Pousada et al., 2017). Long-chain noncoding RNA (lncRNA) is the core of research in many fields of genetics. Recent studies show that cellular activities depend on the regulation of lncRNA (Jarroux et al., 2017; Liu et al., 2018).
The lung adenocarcinoma metastasis-associated gene 1 (MALAT1) is a long-chain folded noncoding RNA (originally found in non-small-cell lung cancer) that is associated with human disease, is widely expressed in various mammals, and is located on chromosome 11q13.1 (Gutschner et al., 2013a, 2013b). Previous studies show that MALAT1 has a very important regulatory role in tumor cell growth, metastasis, and invasion, and is closely related to tumor prognosis (Han et al., 2014; Si et al., 2019). Research by Michalik et al. (2014) found that MALAT1 is involved in regulating the proliferation of vascular endothelial cells and thereby reducing cardiovascular production, which may be related to the pathogenesis of hypertension. In addition, Zhuo et al. (2017) found that the MALAT1 gene rs619586 mutation from A to G can directly upregulate the expression of X-box-binding protein 1 (XBP1) by acting as competitive endogenous RNA (ceRNA) of miR-214, thereby inhibiting vascular endothelial cell proliferation and migration in vitro by shortening SM phase transitions, which plays important roles in the pathogenesis of pulmonary hypertension. Recently, there is a growing body of evidence showing that the single nucleotide polymorphism (SNP) of the MALAT1 gene is associated with the occurrence and development of cardiovascular diseases. For example, Hu et al. (2019) showed that the MALAT1 gene rs4102217 locus SNP was associated with the risk of coronary artery disease, and Li et al. (2018a) found that the MALAT1 rs619586 locus gene polymorphism was associated with a susceptibility to congenital heart disease in the Chinese population.
In this study, we selected the rs664589 locus of the noncoding region of the MALAT1 gene to analyze the correlation between this SNP locus and the susceptibility to essential hypertension in the Han Chinese population. In addition, in this study by online target prediction tools we found that hsa-miR-539-3p and hsa-miR-485-3p have targeted binding sites to both lncRNA MALAT1 and bone morphogenetic protein receptor type 2 (BMPR2) gene. Therefore, we speculate that there may be expression regulation networks in lncRNA MALAT1/hsa- miR-539-3p and hsa-miR-485-3p/BMPR2, which could be potential factors for the occurrence of hypertension. We speculate that the SNP at the rs664589 locus in the noncoding region of the MALAT1 gene affects the expression level of MALAT1, which in turn affects its role as a miRNA molecular sponge, affects the expression of target proteins, and may be a potential pathogenesis of hypertension, but further studies are needed to verify this conclusion.
Materials and Methods
Subjects
A cohort of 260 Han Chinese patients with essential hypertension who received treatments in the Affiliated Yantai Yuhuangding Hospital of Qingdao University were selected as subjects (case group) during May 2017 and March 2019, and another cohort of 260 healthy subjects were recruited as a control group. Diagnostic criteria for essential hypertension were: without using antihypertensive drugs, the blood pressure was measured three times on the same day, and sitting systolic blood pressure (SBP) ≥140 mmHg and/or diastolic blood pressure (DBP) ≥90 mmHg (Liu and Writing Group of 2010 Chinese Guidelines for the Management of Hypertension, 2011). Patients with secondary hypertension, diabetes, coronary heart disease, valvular heart disease, and other organic heart disease, cancer, and immune system diseases were excluded. The control subjects had SBP <140 mmHg and DBP <90 mmHg, no diabetes, heart, or other organic damage, coronary heart disease, tumor, or immune system diseases. This study was approved by the Medical Ethics Committee of the Affiliated Yantai Yuhuangding Hospital of Qingdao University, and all participants signed informed consent documents. The recruitment was in accordance with the principles of the World Medical Association Declaration of Helsinki.
Genotyping
Genomic DNA was extracted from 3 mL of venous blood from the subjects using the QIAamp DNA Blood Mini Kit (Qiagen, Venlo, The Netherlands). The primers for the MALAT1 gene rs664589 locus were: 5′-AGT TTT ATT AAA GGG GAG GGG CA-3′ (forward); and 5′-CCC ACC AAT CCC AAC CGT AA-3′ (reverse). A DNA fragment containing the rs664589 locus of the MALAT1 gene was obtained by PCR. The PCR mixture contains 50 ng of genomic DNA, 2.5 mM dNTP 4 μL, 10 μL of PCR buffer, 1 μL each of 10 pM forward and reverse primers, 0.5 U PrimeSTAR HS DNA polymerase (TaKaRa, Dalian, China), and added ddH2O to a final volume of 50 μL. The PCR conditions were: 94°C, 5 min, 35 cycles; 98°C, 10 s; 58°C, 15 s; 72°C, 2 min, and then extension 72°C, 5 min. Then the Sanger sequencing was performed to analyze the genotype of the MALAT1 gene rs664589 locus.
Cell culture
The human umbilical vein endothelial cells (HUVECs) used in this study were purchased from Shanghai Institutes for Biological Sciences (Shanghai, China). Human embryonic kidney (HEK) 293T cells were purchased from American Type Culture Collection (ATCC, Manassas, VA). All cells were cultured in RPMI 1640 medium (GIBCO-BRL) containing penicillin/streptomycin (Invitrogen, Shanghai, China) and 10% fetal bovine serum (Life Technologies, Australia) at 37°C, in an incubator with 5% CO2.
Cell transfection
The sequences of the MALAT1 and the MALAT1 siRNA (sense: 5′-GGC CAA AUG UUG AAG UUA ATT-3′, anti-sense: 5′-UUA ACU UCA ACA UUU GGC CTT-3′) were synthesized by Sangon Biotech (Shanghai, China), and subcloned into pcDNA3.1 (Invitrogen). The plasmid containing the MALAT1, MALAT1 siRNA sequence, was transfected into HUVECs using Lipofectamine 2000 (Invitrogen). After 48 h, the expressions of the MALAT1, hsa-miR-539-3p, hsa-miR-485-3p, and BMPR2 mRNA, were detected by qRT-PCR, and the expression of BMPR2 protein was detected by Western blot. The empty pcDNA3.1 vector was used as the control (NC).
Quantitative real-time polymerase chain reaction (qRT-PCR)
The total RNA was isolated from the plasma and cultured cells using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The extracted RNA was reverse transcribed into cDNA using the PrimeScript RT Master Mix (TaKaRa). The qRT-PCR was performed using the PrimeScript RT Kit and SYBR Premix Ex Taq (TaKaRa) according to the manufacturer's instructions. The primers were synthesized by Ribobio (Guangzhou, China), and GAPDH was used as an internal control. The primer sequences were as follows: MALAT1: 5′-GTG ATG CGA GTT GTT CTC CG-3′ (forward); 5′-CTG GCT GCC TCA ATG CCT AC-3′ (reverse). hsa-miR-539-3p: 5′-TGA TGA GCT GGC AGT GTA TTG T-3′ (forward); 5′-GAC AGC AGT TGC ATG TTA GC-3′ (reverse). hsa-miR-485-3p: 5′-GCT GGC AGT GTA TTG TTA GCT G-3′ (forward); 5′-ATG CAA TAA GAC AGC AGT TGC AT-3′ (reverse). BMPR2 mRNA: 5′-AAT ACT CGC ACT TCC TCA GAA CC-3′ (forward); 5′-AGC ATA GCA AGG CTT CAG ACA G-3′ reverse). U6: 5′-CTC GCT TCG GCA GCA CA-3′ (forward), 5′-TGG TGT CGT GGA GTC G-3′ (reverse). GAPDH: 5′-GTC AAC GGA TTT GGT CTG TAT T-3′ (forward); 5′-AGT CTT CTG GGT GGC AGT GAT-3′ (reverse). The conditions for PCR were: 95°C, 30 s; 95°C, 5 s; and 60°C, 34 s for 40 cycles. The qRT-PCR was conducted using an ABI 7500 real-time PCR system (Applied Biosystems, Foster City, CA). The 2−ΔΔCT method was used to calculate the fold changes of the lncRNA MALAT1, hsa-miR-539-3p, hsa-miR-485-3p relative to U6, and BMPR2 mRNA relative to GAPDH.
Western blot
The expression level of BMPR2 protein in HUVECs after transfection was detected by western blot. The collected HUVECs were first washed with phosphate-buffered saline (PBS; Gibco-Invitrogen, 10010023) and lysed with the RIPA lysis buffer (Beyotime Biotechnology, China). A BCA protein assay method (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) was used to determine the concentration of the proteins. The proteins were separated using a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred to a polyvinylidene fluoride membrane (Bio-Rad, Chicago, IL). The membrane was blocked with 5% milk for 2 h at room temperature, and then incubated with primary antibody (anti-BMPR2; 1:1000, cat. no. 6979) at 4°C overnight. It was then treated with goat anti-rabbit IgG horseradish peroxidase-conjugated secondary antibody (1:1000; cat. no. A0208; Beyotime Institute of Biotechnology, Haimen, China) for 2 h at room temperature. The ECL Chemiluminescence Kit (Millipore, Billerica, MA) was used to visualize the protein bands, and the GAPDH was used as an internal control.
Luciferase reporter gene assay
HEK293 cells were seeded in a 96-well plate and incubated at 37°C for 24 h. A MALAT1-Luc vector with wild-type (MALAT1 WT) or mutant MALAT1 (MALAT1 MT) was established at the binding sites of the hsa-miR-539-3p and hsa-miR-485-3p on the MALAT1. A BMPR2-Luc vector with wild-type (BMPR2 WT) or mutant BMPR2 (BMPR2 MT) was established at the binding sites of the hsa-miR-539-3p and hsa-miR-485-3p on the 3′ untranslated region (UTR) of the BMPR2 gene. The MALAT1 WT, MALAT1 WT+hsa-miR-539-3p mimics, MALAT1 WT+hsa-miR-539-3p inhibitor, MALAT1 WT+hsa-miR-539-3p mimics NC, MALAT1 WT+hsa-miR-539-3p inhibitor NC; MALAT1 MT, MALAT1 MT+hsa-miR-539-3p mimics, MALAT1 MT+hsa-miR-539-3p inhibitor, MALAT1 MT+hsa-miR-539-3p mimics NC, MALAT1 MT+hsa-miR-539-3p inhibitor NC; BMPR2 WT, BMPR2 WT+hsa-miR-539-3p mimics, BMPR2 WT+hsa-miR-539-3p inhibitor, BMPR2 WT+hsa-miR-539-3p mimics NC, BMPR2 WT+hsa-miR-539-3p inhibitor NC; and BMPR2 MT, BMPR2 MT+hsa-miR-539-3p mimics, BMPR2 MT+hsa-miR-539-3p inhibitor, BMPR2 MT+hsa-miR-539-3p mimics NC, and BMPR2 MT+hsa-miR-539-3p inhibitor NC were cotransfected into HEK293 cells using Lipofectamine 2000. After 48 h, the luciferase activity of the cells was measured using a dual luciferase assay system (Promega).
Statistical analyses
All statistical analyses in this study were performed using SPSS 22.0 (SPSS) and GraphPad Prism 7 (GraphPad). Continuous variables were expressed as mean ± standard deviation and comparisons between groups were performed using the independent Student's t-test. The categorical variables were expressed as a percentage [n (%)], and the comparison between groups was performed using the χ2 test. The χ2 test was used to determine if the genotype frequency was consistent with Hardy-Weinberg equilibrium (HWE). The correlation between the MALAT1 gene rs664589 locus SNP and the risk of hypertension was determined based on the distribution of allele frequency and genetic models (dominant, recessive, and additive models). The odds ratio (OR) and 95% confidence interval (CI) were used in an unconditional logistic regression, and adjusted for age, gender, body mass index (BMI), smoking status, drinking status, and other factors. Pearson correlation analysis was used to assess the correlation between lncRNA MALAT1 and microRNA levels. All tests were two-tailed. p < 0.05 was considered statistically significant.
Results
Clinical characteristics
The general clinical characteristics of the case and control groups are shown in Table 1. In this study, we selected 260 patients with hypertension, including 137 males and 123 females, 33-89 years of age, with a mean of 56.01 ± 10.77 years. A total of 260 healthy controls were selected, including 132 males and 128 females, 34-88 years of age, with a mean of 56.34 ± 10.45 years. There were no significant differences in the age, gender, BMI, smoking status, drinking status, and high-density lipoprotein cholesterol between the patients with hypertension and the control group (p > 0.05). However, the SBP, DBP, total cholesterol, total triglycerides, and fasting blood glucose were significantly higher in patients with hypertension than in the control group (p < 0.001).
Comparison of General Clinical Characteristics Between the Case and Control Groups
BMI, body mass index; DBP, diastolic blood pressure; FBG, fasting blood glucose; HDL-C, high-density lipoprotein cholesterol; SBP, systolic blood pressure; SD, standard deviation; TC, total cholesterol; TG, total triglycerides.
MALAT1 gene rs664589 locus SNP was associated with the risk of hypertension
The genotype frequencies of the MALAT1 gene rs664589 locus in the selected control subjects of this study were consistent with HWE (p > 0.05); the patients in the case group were not consistent with HWE (p < 0.05; Table 2). Taking the CC genotype as a reference, we found that the difference in the frequency of the CG genotype between the case group and the control group was not statistically significant (adjusted OR = 1.16, 95% CI: 0.93-1.42, p = 0.18). However, the frequency of the GG genotype was significantly higher in the case group than in the control group (adjusted OR = 1.75, 95% CI: 1.31-2.03, p < 0.001), and the risk of hypertension was significantly increased in both the dominant and recessive models (adjusted OR = 1.28, 95% CI: 1.07-1.52, p = 0.009; adjusted OR = 1.68, 95% CI: 1.27-1.94, p = 0.001; respectively), but no significant change in the risk of hypertension was found in the additive model (adjusted OR = 1.04, 95% CI: 0.91-1.19, p = 0.58). The MALAT1 gene rs664589 locus G allele carriers were 1.33 times more likely to have hypertension than the C allele carriers (95% CI: 1.15-1.51, p < 0.001; Table 2).
Correlation Between Genotype and Allele Frequency of the MALAT1 Gene rs664589 Locus and the Risk of Hypertension
HWE p-value for the case group.
HWE p-value for the control group.
Adjusted for age, gender, BMI, smoking, and drinking statuses.
CI, confidence interval; HWE, Hardy-Weinberg equilibrium; MALAT1, lung adenocarcinoma metastasis-associated gene 1; OR, odds ratio.
Stratified analysis
We then divided the participants of this study into different subpopulations and performed stratified analyses: young subjects (age <60 years), and elderly subjects (age ≥60 years); males and females; nonobese subjects (BMI <24 kg/m2), and obese subjects (BMI ≥24 kg/m2); subjects with a history of smoking, and subjects without a history of smoking; and subjects with a history of drinking, and subjects without a history of drinking (Table 3). No significant correlation between the MALAT1 gene rs664589 locus SNP and the risk of hypertension was detected in the young and elderly subjects (p > 0.05). However, the MALAT1 gene rs664589 locus SNP was significantly associated with the risk of hypertension in males, obese subjects, subjects without a history of smoking, and subjects without a history of drinking (p < 0.05; Table 3).
Stratified Analysis of the Relationship Between Genotypes and Allele Frequencies of the MALAT1 Gene rs664589 Locus and the Risk of Hypertension
Adjusted for age, gender, BMI, smoking, and drinking statuses.
Numbers in bold indicate significant difference.
The correlation between the MALAT1 gene rs664589 locus SNP and systolic and DBPs
We analyzed the correlation between the SBP and DBP in different MALAT1 gene rs664589 locus genotypes. The results showed that the SBP and DBP were significantly higher in patients with the G allele than those with the C allele (p < 0.01; Fig. 1).
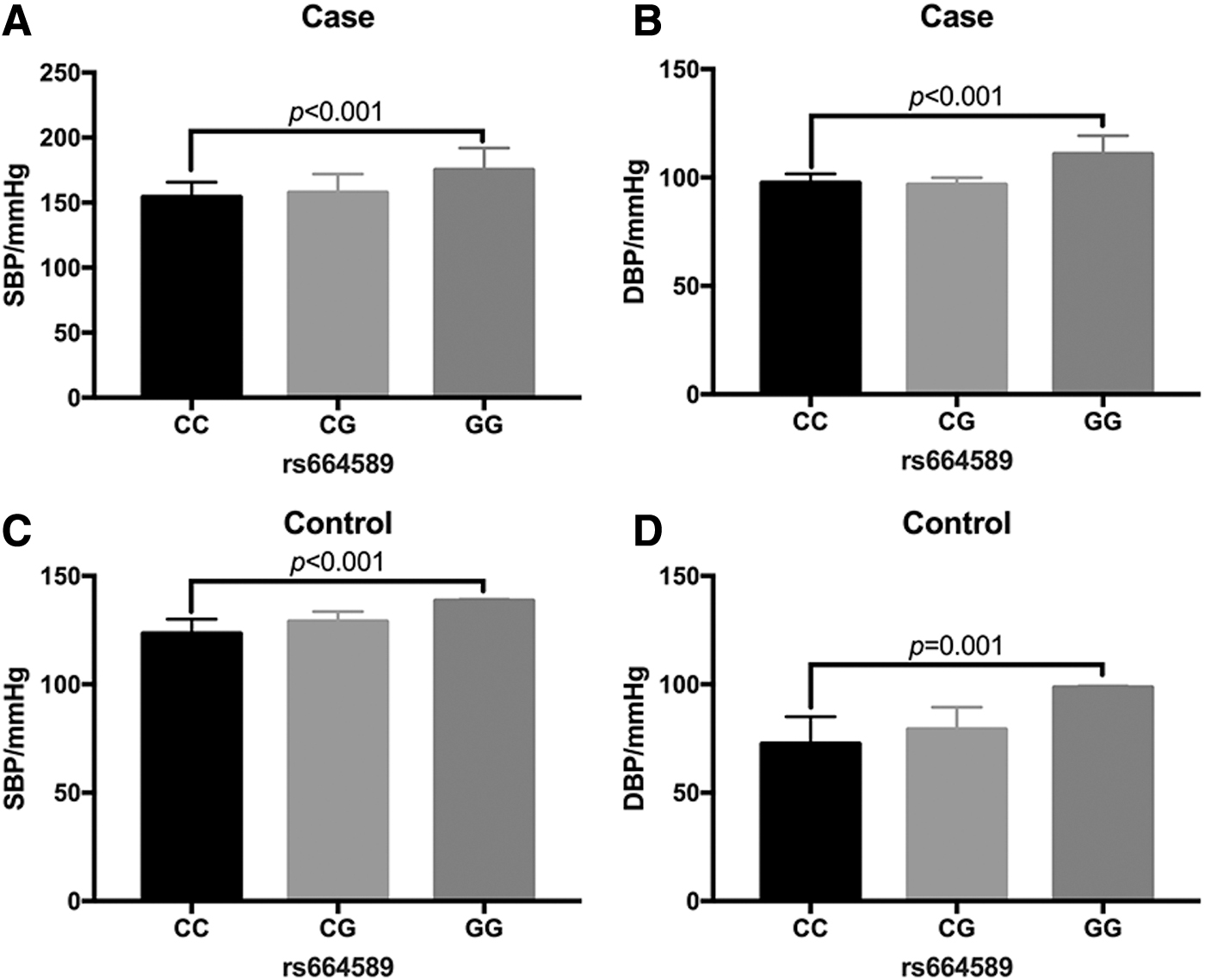
The MALAT1 gene rs664589 locus SNP was associated with systolic and DBPs. Comparison of SBP
Downregulation of the lncRNA MALAT1 in the plasma of patients with hypertension
We detected the expression levels of lncRNA MALAT1, hsa-miR-539-3p, and hsa-miR-485-3p in the plasma of participants by qRT-PCR. We found that lncRNA MALAT1 was lower in the plasma of patients with hypertension compared with that in the controls (Fig. 2A), while the hsa-miR-539-3p and hsa-miR-485-3p were higher in the plasma of patients with hypertension compared with that in the controls (Fig. 2B, C). We further analyzed the correlation between the expression levels of lncRNA MALAT1, and hsa-miR-539-3p and hsa-miR-485-3p in the plasma, and found that there was a significant negative correlation between the expression levels of lncRNA MALAT1 and hsa-miR-539-3p and hsa-miR-485-3p in the plasma of patients with hypertension (Fig. 2D, E), whereas the expression level of lncRNA MALAT1 was also negatively correlated with the expression levels of hsa-miR-539-3p and hsa-miR-485-3p in the control group (Fig. 2F, G).
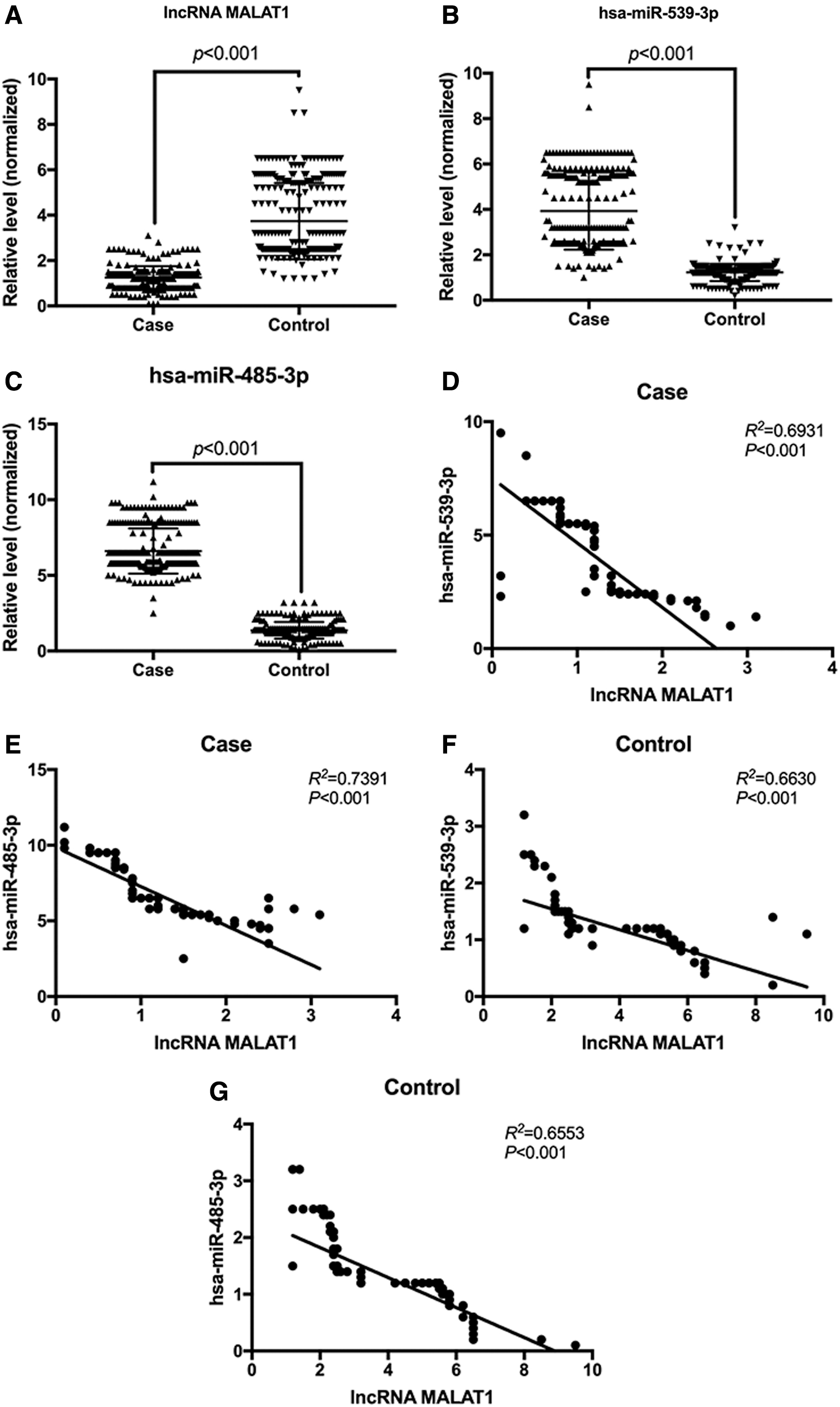
Expression levels of lncRNA MALAT1 and hsa-miR-539-3p and hsa-miR-485-3p in plasma.
The correlation between the MALAT1 gene rs664589 locus SNP and the plasma level of lncRNA MALAT1
qRT-PCR analysis showed that in the patients with hypertension and the healthy controls, the level of plasma lncRNA MALAT1 in subjects carrying the G allele was significantly lower than that in the C allele carriers (p < 0.001; Fig. 3A, B).

The MALAT1 gene rs664589 locus SNP was correlated with the plasma level of lncRNA MALAT1.
The lncRNA MALAT1 inhibited the expression of hsa-miR-539-3p and hsa-miR-485-3p, and promoted the expression of BMPR2 protein
We detected the level of lncRNA MALAT1 in the HUVECs after transfection through qRT-PCR. The results showed that the levels of MALAT1 were significantly higher in HUVECs transfected with MALAT1, whereas the MALAT1 levels were significantly reduced after transfection with MALAT1 siRNA (Fig. 4A). The expression levels of hsa-miR-539-3p and hsa-miR-485-3p in HUVECs were significantly decreased, and the expression levels of BMPR2 mRNA were significantly increased after transfection with MALAT1. However, the expression levels of the hsa-miR-539-3p and hsa-miR-485-3p were significantly elevated, and the expression levels of the BMPR2 mRNA were significantly reduced in HUVECs transfected with MALAT1 siRNA (Fig. 4A). We further determined, by western blot, that MALAT1 promoted the expression of BMPR2 protein (Fig. 4B).
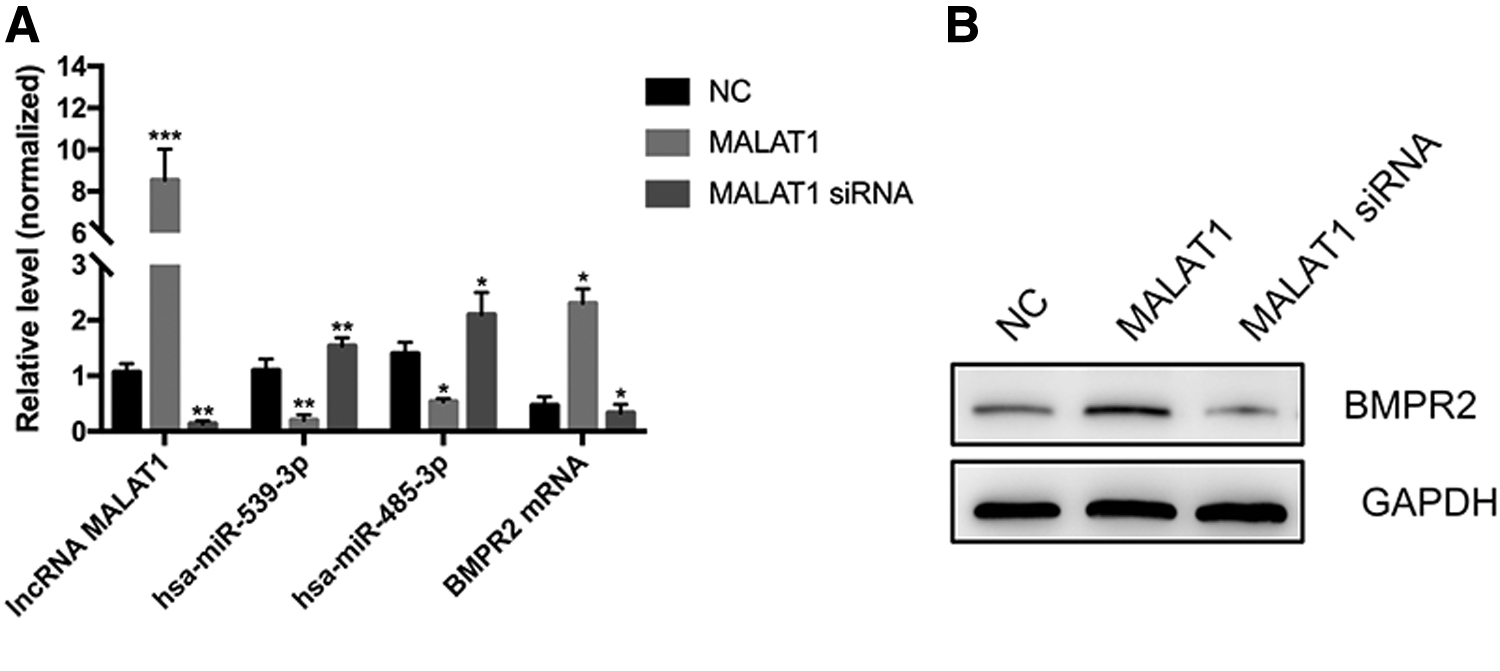
The lncRNA MALAT1 inhibited the expression of hsa-miR-539-3p and hsa-miR-485-3p and promoted the expression of BMPR2 protein.
The hsa-miR-539-3p and hsa-miR-485-3p were targets of MALAT1, and BMPR2 was the target of hsa-miR-539-3p and hsa-miR-485-3p
We found that there were targeted binding sites for hsa-miR-539-3p and hsa-miR-485-3p on MALAT1 (Fig. 5A), and there were targeted binding sites for the hsa-miR-539-3p and hsa-miR-485-3p on the 3′UTR of the BMPR2 through the TargetScan 3.1 online prediction software (Fig. 5B). In 293T cells after cotransfection of hsa-miR-539-3p or hsa-miR-485-3p with MALAT1 WT or MALAT1 MT, the dual luciferase reporter assay revealed that in the MALAT1 group, the dual luciferase activity was significantly decreased in the hsa-miR-539-3p mimic and hsa-miR-485-3p mimic groups compared with that in the hsa-miR-539-3p NC and hsa-miR-485-3p NC groups. However, the dual luciferase activity was significantly increased in the hsa-miR-539-3p inhibitor and hsa-miR-485-3p inhibitor groups compared with that in the hsa-miR-539-3p NC and hsa-miR-485-3p NC groups (p < 0.05; Fig. 5C, D). These findings indicated that hsa-miR-539-3p and hsa-miR-485-3p are targets of MALAT1 WT, not targets of MALAT1 MT.

Results of the dual luciferase reporter gene assay.
Moreover, in 293T cells cotransfected with hsa-miR-539-3p or hsa-miR-485-3p and BMPR2 WT or BMPR2 MT reporter genes, the dual luciferase activity was significantly decreased in the hsa-miR-539-3p mimic and hsa-miR-485-3p mimic groups compared with that in the hsa-miR-539-3p NC and hsa-miR-485-3p NC groups after transfection of BMR2 WT. However, the dual luciferase activity was significantly increased in the hsa-miR-539-3p inhibitor and hsa-miR-485-3p inhibitor groups compared with that in the hsa-miR-539-3p NC and hsa-miR-485-3p NC groups (p < 0.05; Fig. 5E, F). These findings indicated that hsa-miR-539-3p and hsa-miR-485-3p are targets of BMPR2 WT, not targets of MALAT1 MT.
Discussion
In the present study, we found that the MALAT1 gene rs664589 locus SNP was significantly associated with the risk of hypertension, and the G allele gene was susceptible for hypertension. The expression level of lncRNA MALAT1 in the plasma is significantly decreased in the G allele carriers. The expression levels of hsa-miR-539-3p and hsa-miR-485-3p were significantly increased, whereas the expression of BMPR2 was significantly inhibited in the G allele carriers, which is likely to be associated with hypertension, but needs to be confirmed in animal models.
The blood pressure level is causally linked to the risk of morbidity and mortality in cardiovascular and cerebrovascular diseases. An elegant study of 61 worldwide populations (∼1 million people, 40-89 years old) found that the baseline blood pressure ranged from 115/75 to 185/115 mmHg with a mean follow-up of 12 years. The results revealed a continuous, independent, and directly positive correlation between SBP or DBP and stroke, and the risk of coronary heart disease events and cardiovascular deaths (Lewington et al., 2002). For every 20-mmHg increase in SBP or 10-mmHg increase in DBP, the risk of heart and cerebrovascular disease is doubled.
MALAT1 is a lncRNA located at the nucleus of the cell, which is tRNA-like and lacks a sufficient length of open reading frame to form a template for encoding proteins (Brown et al., 2012, 2014). MALAT1 is 8.5 kb in length. Current studies show that MALAT1 is a potential biomarker for tumors (Li et al., 2018b) and plays an important role in tumor formation and progression. Ji et al. (2019) found that MALAT1 regulates the transcriptional and translational levels of the protooncogene RUNX2 in colorectal cancer metastasis. Hu et al. (2017) found that lncRNA MALAT1 regulates autophagy-related chemoresistance through chelation of miR-23b-3p in gastric cancer. Furthermore, it was found that the role of MALAT1 in cardiovascular disease is also very important. A prior study found that the MALAT1 levels in peripheral blood cells of patients with acute myocardial infarction are significantly higher than that in healthy subjects (Yu et al., 2018). Zhu et al. (2019) found that MALAT1/miR-15b-5p/MAPK1 mediates autophagy of endothelial progenitor cells and affects coronary atherosclerotic heart disease through the mTOR signaling pathway. In the present study, we found that the plasma MALAT1 was significantly lower in patients with hypertension than in healthy subjects, suggesting that MALAT1 may be a potential biomarker for hypertension.
SNP mainly refers to polymorphisms in DNA sequences caused by single nucleotide variations at the genomic level, and is a third-generation genetic marker. Current evidence shows that SNP is closely related to susceptibility to disease and drug treatment in different populations. The SNPs on lncRNA have attracted more attention in recent years. The rs664589 is located in the noncoding region of the MALAT1 gene, and this locus SNP is associated with the occurrence of various diseases. For example, Wu et al. (2019) showed that SNPs of the MALAT1 rs664589 polymorphism inhibit the binding of miR-194-5p, leading to the increased risk of colorectal cancer. Wang et al. (2018) found that rs619586 locus of the MALAT1 is associated with the risk of coronary atherosclerotic heart disease. In this study, we found that the SBP and DBP of the G allele carriers of the rs664589 locus of the MALAT1 gene were significantly higher than those of the C allele carriers, and the G allele gene was susceptible for hypertension. The genotype frequency of MALAT1 gene rs664589 locus is not consistent with HWE (p < 0.05) in the case group of the present study, which may be due to the small sample size of the selected population, which cause random deviations between the gene frequency and the expected ratio. Further validation needs to be performed in a large sample size.
Using an online target prediction tool, we found that there was a targeted binding site of hsa-miR-539-3p and hsa-miR-485-3p on the MALAT1. Additionally, there was a targeted binding site of hsa-miR-539-3p and hsa-miR-485-3p on the 3′UTR of the BMPR2 gene. In this study, we also confirmed these target binding sites by the dual luciferase reporter assay. From the results of this study, it is speculated that MALAT1 is a molecular sponge for hsa-miR-539-3p and hsa-miR-485-3p that regulates the expression of the BMPR2 gene. In plasma, we found that the MALAT1 levels were significantly negatively correlated with hsa-miR-539-3p and hsa-miR-485-3p, and it was also confirmed by cell transfection experiments that MALAT1 could negatively regulate the expression of hsa-miR-539-3p and hsa-miR-485-3p, while upregulating the expression of BMPR2. It is well known that BMPR2 is a cell membrane receptor of bone morphogenetic protein. The bone morphogenetic proteins belong to the transforming growth factor-beta family, which play a key role in embryonic development and tissue homeostasis, inhibit proliferation of vascular smooth muscle cells and endothelial cells, and induce cell apoptosis (Diebold et al., 2015; Gomez-Puerto et al., 2019; Harper et al., 2019). Downregulation of BMPR2 might be an important cause of hypertension, but further research is needed to confirm it.
There are some limitations in this study that need further improvement. First, we only found a correlation between the rs664589 locus SNP and the risk of hypertension in a small sample; this needs to be further confirmed in a larger sample size. Second, the population we selected was limited to the Han Chinese population. Further research is needed to determine whether this correlation exists in other populations. Additionally, we speculate that the possible mechanism of this correlation is that the rs664589 locus SNP affects the MALAT1 expression level, and then affects the expression of BMPR2 through hsa-miR-539-3p and hsa-miR-485-3p targeted regulation, which requires an in vivo model for confirmation.
Conclusion
The lncRNA MALAT1 gene rs664589 locus SNP was significantly associated with the risk of hypertension. The G allele gene was susceptible for hypertension, and the expression level of lncRNA MALAT1 in plasma was significantly lower in subjects carrying the G allele. A possible mechanism of the association with hypertension is that low expression levels of MALAT1 may result in abnormally high expression of hsa-miR-539-3p and hsa-miR-485-3p, thereby inhibiting the expression of BMPR2, but further studies are needed in animal models to confirm this speculation.
Authors' Contributions
S.W. conceived and designed the experiments. X.M., J.W., and L.W. performed the experiments. X.M. analyzed the data. S.W. wrote the article.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Platform Project of the National Brain Defense Committee (GN-2018R0011).