
Abstract
Introduction:
The classification of myeloproliferative neoplasms (MPN) is currently based on the genotype. Thus, to achieve better diagnostic and prognostic outcomes, it is necessary to further investigate the genetic spectrum underlying the pathogenesis of MPNs. The rs2736100A>C is a functional single nucleotide polymorphism in the telomerase reverse transcriptase (TERT) gene that has been previously reported to be associated with the risk of MPNs. Herein, we performed a meta-analysis to confirm the relationship between the TERT rs2736100A>C polymorphism and MPN susceptibility.
Materials and Methods:
Studies of case-control design were acquired from online databases with specific inclusion criteria. Odds ratios (ORs) with 95% confidence intervals (95% CI) were estimated to evaluate the association between the TERT rs2736100 polymorphism and MPN susceptibility using different genetic models.
Results:
Ten case-control studies involving 3488 cases and 57,948 controls were examined. Overall, there was a significant association between the TERT rs2736100 polymorphism and the risk of MPNs (allele model [C vs. A]: OR = 1.57 [95% CI: 1.47-1.69]; homozygous model [CC vs. AA]: OR = 3.00 [95% CI: 2.40-3.76]; heterozygous model [AC vs. AA]: OR = 2.17 [95% CI: 1.77-2.66]; dominant model [CC+AC vs. AA]: OR = 2.43 [95% CI: 2.00-2.95]; and recessive model [CC vs. AC+AA]: OR = 1.73 [95% CI: 1.47-2.04]).
Conclusions:
In this meta-analysis, we confirm an association between the TERT rs2736100A>C polymorphism and MPN susceptibility under all genetic models evaluated. The TERT rs2736100A>C allele increases the overall risk of MPN. Further studies are warranted to determine the functional role of the TERT rs2736100 polymorphism in MPN.
Introduction
Myeloproliferative neoplasms (MPNs) are characterized as an aberrant clonal expansion of differentiated hematopoietic cells in bone marrow. This disease is mainly classified into two subtypes depending on the presence of the Ph chromosome. Ph chromosome-positive MPNs are also defined as chronic myelogenous leukemia (CML). Ph chromosome-negative MPNs include three major types: polycythemia vera (PV), primary myelofibrosis (PMF), and essential thrombocytosis (ET) (Abruzzese et al., 2018). PV is a clonal disease caused by mutations in pluripotent hematopoietic stem cells, following with the increase of erythroid cells, granulocytes, megakaryocytes, and B lymphocytes in bone marrow. The PMF is often accompanied by reactive fibrous connective tissue deposition and extramedullary hematopoiesis. The predominant symptoms of PMF are mainly due to the overproduction of megakaryocytes and granulocytes in bone marrow. ET is initiated with a single abnormal clonal proliferation of megakaryocytes and a subsequent increase of platelet count (Helbig, 2018). Current diagnosis of MPN subsets in clinic is basically relied on the newest version of World Health Organization (WHO) guidelines, according to the aforementioned histological evaluation in bone marrow biopsies (Hong and He, 2017).
Although the pathogenesis, diagnosis, and prognosis patterns vary among different MPNs, current understanding of the evolution of different PMNs uniformly begins with a review of their molecular basis. With long-term exploration, many mutations have been proved to be related to MPNs. Besides CML, classical (Ph)-negative MPNs have been determined highly related to three main mutations: Janus kinase 2 (JAK2), calreticulin (CALR), and myeloproliferative leukemia virus oncogene (MPL) (Pardanani et al., 2007).
Telomerase, a DNA-dependent polymerase, contains multiple repeating short sequences at the end of eukaryotic chromosomes. They are essential to prevent the chromosomes from degradation, end-end fusion, and rearrangement. In humans, the telomere repeat is TTAGGG bound to particular proteins (Lu et al., 2013). Two major components of telomerase are telomerase reverse transcriptase (TERT) and telomerase RNA component (TERC). Compared with TERC, TERT is rarely expressed in human somatic tissue. However, the alteration of TERT is common and highly related to the abnormal upregulation of telomerase in most malignancies, including MPNs (Nandakumar and Cech, 2013). TERT rs2736100 locus, located at the intron 2 of TERT, has been widely reported to play an indispensable role in the synthesis of TERT protein and activation of telomerase (Bernardes and Blasco, 2013).
Recently, a growing number of studies show that TERT rs2736100 A>C single nucleotide polymorphism (SNP) is highly related to susceptibility of MPNs (Dahlstrom et al., 2016; Trifa et al., 2016, 2018; Chiang et al., 2017). Yet, a solid consensus has not been reached about the role of TERT rs2736100A>C in MPNs. Herein, a systematic review and meta-analysis should be of great priority to quantitatively analyze and firmly demonstrate the association between TERT rs2736100A>C and susceptibility of MPNs.
Materials and Methods
Literature search strategy
This meta-analysis was conducted according to Preferred Reporting Item for Systematic Reviews and Meta-analyses statement. Literature research within three online literature databases (Embase, PubMed, and Web of Science) was conducted to identify and select relative studies. There was no restriction about language and time endpoint of online search. Following terms were applied to comprehensively identify studies: (1) (TERT) or (telomerase reverse transcriptase) and (2) (disorder, myeloproliferative) or (disorders, myeloproliferative) or (myeloproliferative disorder) or (myeloproliferative neoplasms) and (3) (genetic polymorphisms) or (polymorphisms, genetic) or (genetic polymorphism). References of included studies were conditionally screened, and on-topic articles were included whereas off-topic items were excluded.
Criteria of inclusion and exclusion
The inclusion criteria were applied: (1) case-control studies, (2) patients were diagnosed with MPNs, and (3) TERT rs2736100A>C status was reported. The exclusion criteria were applied: (1) reviews, single case report, or conference abstract; (2) studies not relevant to TERT rs2736100 polymorphism and MPNs; and (3) duplicate or overlapping data. If several publications from the same project were identified simultaneously, the newest version and most comprehensive data would be included.
Data extraction
Two reviewers (T.H. and Q.L.) independently screened the full text and supplementary materials of eligible studies. For each study, the surname of the first author, the year of publication, the geographic location, the sample sizes, and the number of cases with a positive trait of interest and controls were recorded. Also, we extracted data regarding the ethnicities of the patient.
Statistical analysis
Revman version 5.1 (Revman the Cochrane Collaboration; Oxford, England) and Stata version 14 (Stata Corporation, College Station, TX) were applied to perform this meta-analysis. Pooled odds ratios (OR) in different genetic models were presented to evaluate the association between TERT rs2736100A>C and susceptibility of MPNs. The degree of heterogeneity was assessed by χ2 test combined with the I2 method (I2 ≤ 25%, low heterogeneity; 25% <I2 <75%, medium heterogeneity; and 75% ≤ I2, high heterogeneity). Random-effect model would be applied if high heterogeneity; otherwise, fixed-effect model would be applied if medium or low heterogeneity. The p-value chosen for statistical significance was p < 0.05. The funnel plot was applied to precisely estimate the publication bias of eligible studies.
Results
Literature selection
The initial search strategy identified 156 records after systematic literature retrieval. After removal of 29 duplicates and exclusion of 115 irrelevant records, we included 12 articles after review of the title and abstract. The full texts of the remaining articles were carefully screened and assessed. Four pieces of literature were excluded due to the following reasons: reviews, single case report, and repeated data. Eventually, 8 articles with 10 case-control studies were included for meta-analysis according to our specific inclusion and exclusion criteria. The detailed search strategy is shown in Figure 1.
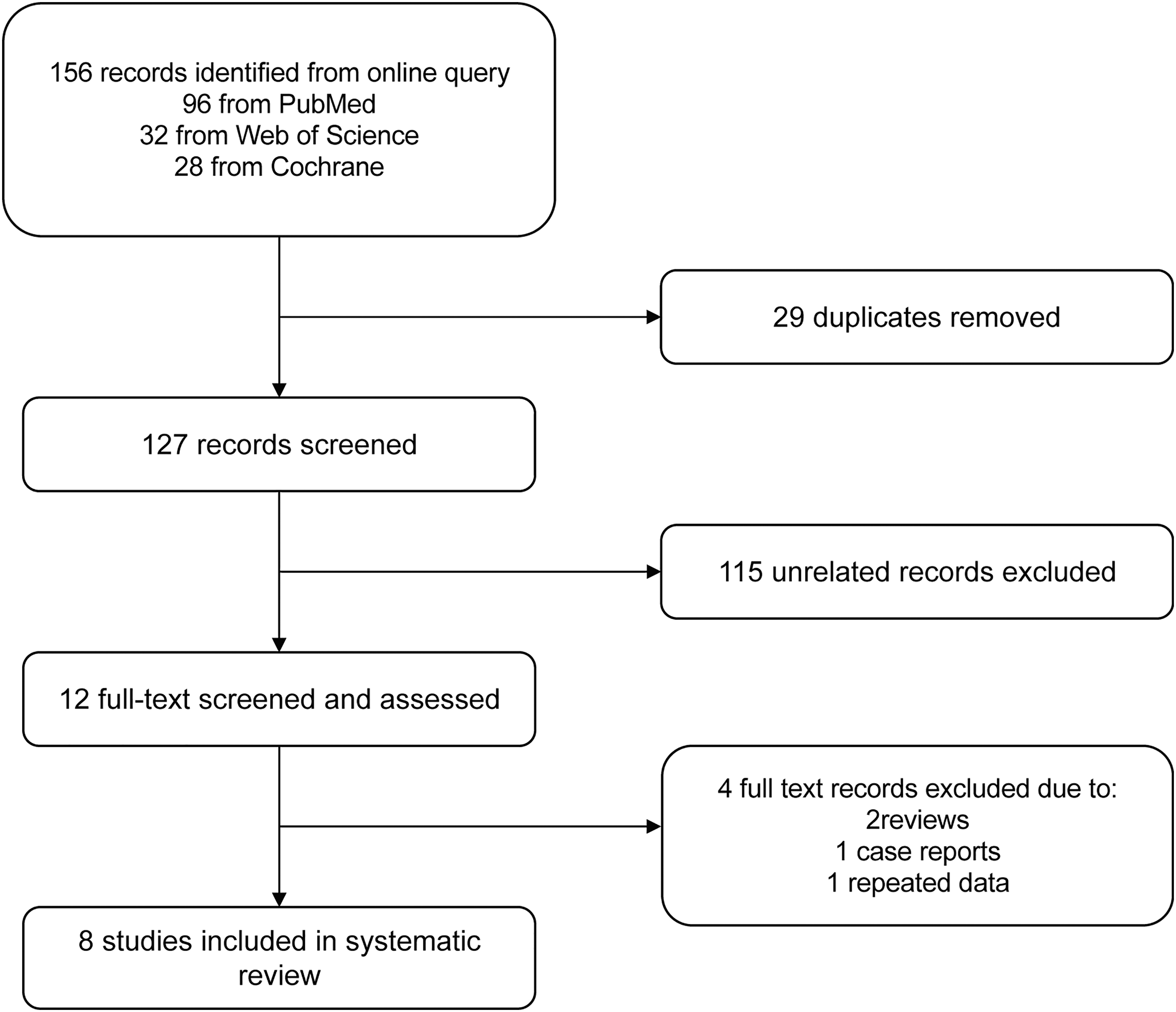
Flow diagram of the search strategy and study selection.
Main characteristics of the included studies
Referring to the included studies, they were all designed for randomized, parallel groups. All the patients were diagnosed as MPN patients based on the 2008 WHO classification. Chiang et al. took samples from 178 Taiwanese MPN patients from MacKay Memorial Hospital and under the approval by the institutional review board of MacKay Memorial Hospital. Trifa et al. included 939 patients with MPN, who were diagnosed between 1984 and 2017 at various hematology units in Romania and 483 controls with comparative ages and genders. Their sequencing was approved by the Iuliu Hatieganu University of Medicine. In total, 101 Chinese MPN patients and 101 matched healthy adults were recruited from Shandong University Hospitals in Dahlstrom's study. Patient cohorts consisted of 584 individuals diagnosed with MPN from Department of Hematology and Stem Cell Transplantation Hospital and 400 healthy Hungarian individuals as controls in Krahling's study. And Tapper et al. included 262 MPN cases and 2674 controls were collected. In total, 717 MPN patients and 202 healthy controls were obtained from Italy and approved by the ethics committee in Jager's study. Oddsson et al. collected 237 Icelanders diagnosed with MPN and 34,128 Icelandic controls. Masafumi also selected 201 patients and 366 healthy people as control. Finally, we collected data from these 10 case-control studies including 3488 patients with MPNs and 57,948 healthy participants. The sample groups with different genetic backgrounds were included (Asian and Caucasian). The main characteristics of included studies are given in Table 1.
Baseline Characteristics of Qualified Studies in this Meta-Analysis
Pooled analysis and heterogeneity detection
The association between TERT rs2736100A>C and susceptibility of MPN was estimated with pooled ORs (with 95% confidence interval [CI]). All available data from eligible studies were pooled into the meta-analysis. The overall result showed that TERT rs2736100A>C was significantly related to susceptibility of MPNs. The C allele of TERT rs2736100 was positively related to increased risk of MPNs (allele model [C vs. A]: OR = 1.57 [95% CI: 1.47-1.69] through fixed model, I2 = 37%, p = 0.11; Fig. 2). Furthermore, the association between TERT rs2736100 polymorphism and MPN susceptibility was analyzed among all genetic-model subgroups according to the genotype of patients and healthy controls. The results showed that TERT rs2736100 polymorphism dramatically increased the risk of MPNs (homozygous model [CC vs. AA]: OR = 3.00 [95% CI: 2.40-3.76] through fixed model, I2 = 0%, p = 0.88; heterozygous model [AC vs. AA]: OR = 2.17 [95% CI: 1.77-2.66] through fixed model, I2 = 0%, p = 1.00; dominant model [CC+AC vs. AA]: OR = 2.43 [95% CI: 2.00-2.95] through fixed model, I2 = 0%, p = 0.98; and recessive model [CC vs. AC+AA]: OR = 1.73 [95% CI: 1.47-2.04] through fixed model, I2 = 0%, p = 0.68; Fig. 3).

Forest plot of the association between TERT rs2736100A>C polymorphism and susceptibility of MPNs in the allele model (C vs. A). MPNs, myeloproliferative neoplasms; TERT, telomerase reverse transcriptase.
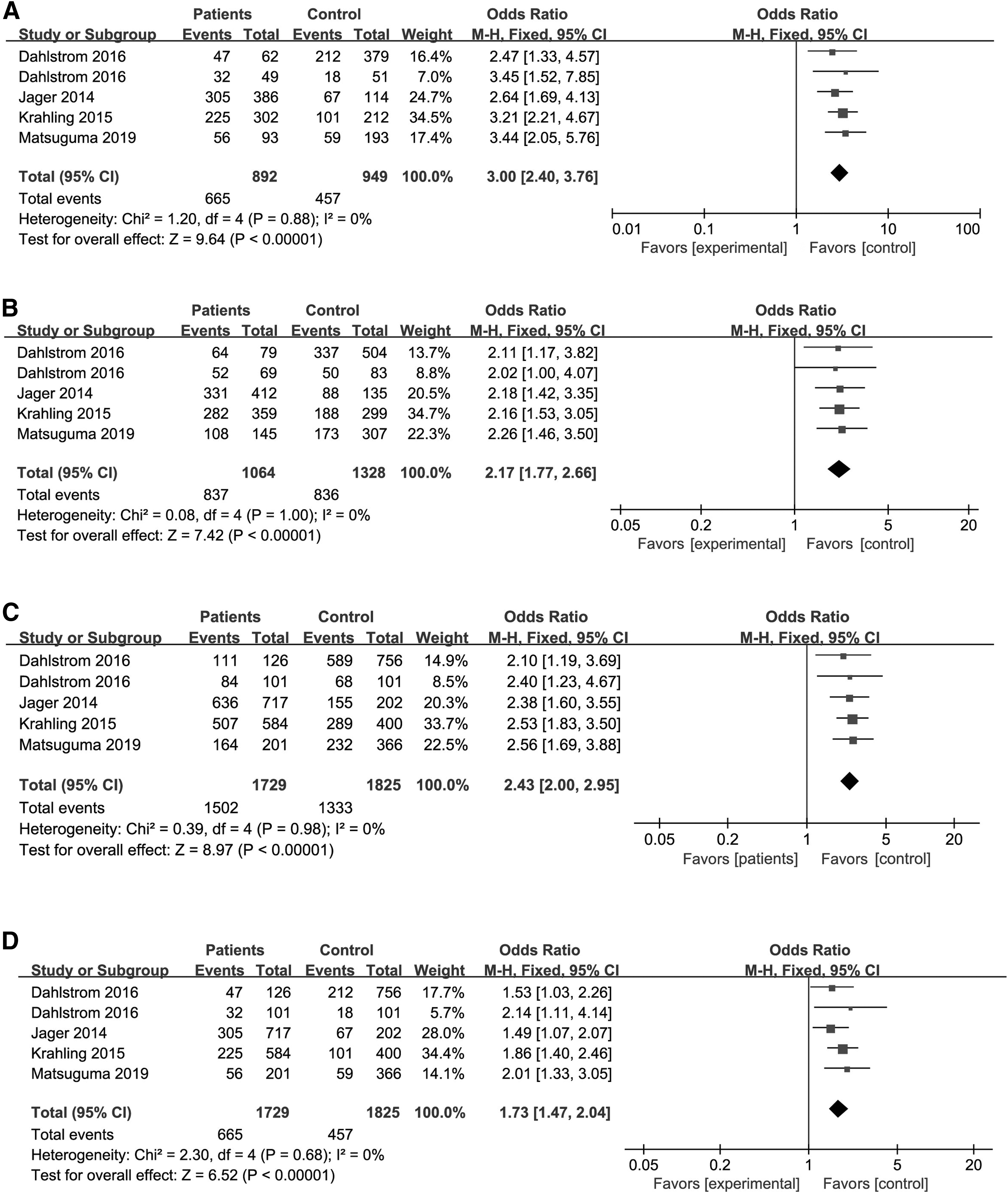
Forest plot of the association between the TERT rs2736100A>C polymorphism and susceptibility of MPNs in homozygous model
Sensitivity analysis
To judge the contribution of variation caused by an individual study in the allele model, the results were pooled with sensitivity analysis. Individual study was excluded from the analysis and pooled OR (with 95% CI) was calculated for each combination. There was no significant change after removal of any individual study. In other words, none of the included studies had significant variation to cause unreliable overall results. Therefore, the results of our meta-analysis were stable and reliable (Fig. 4).
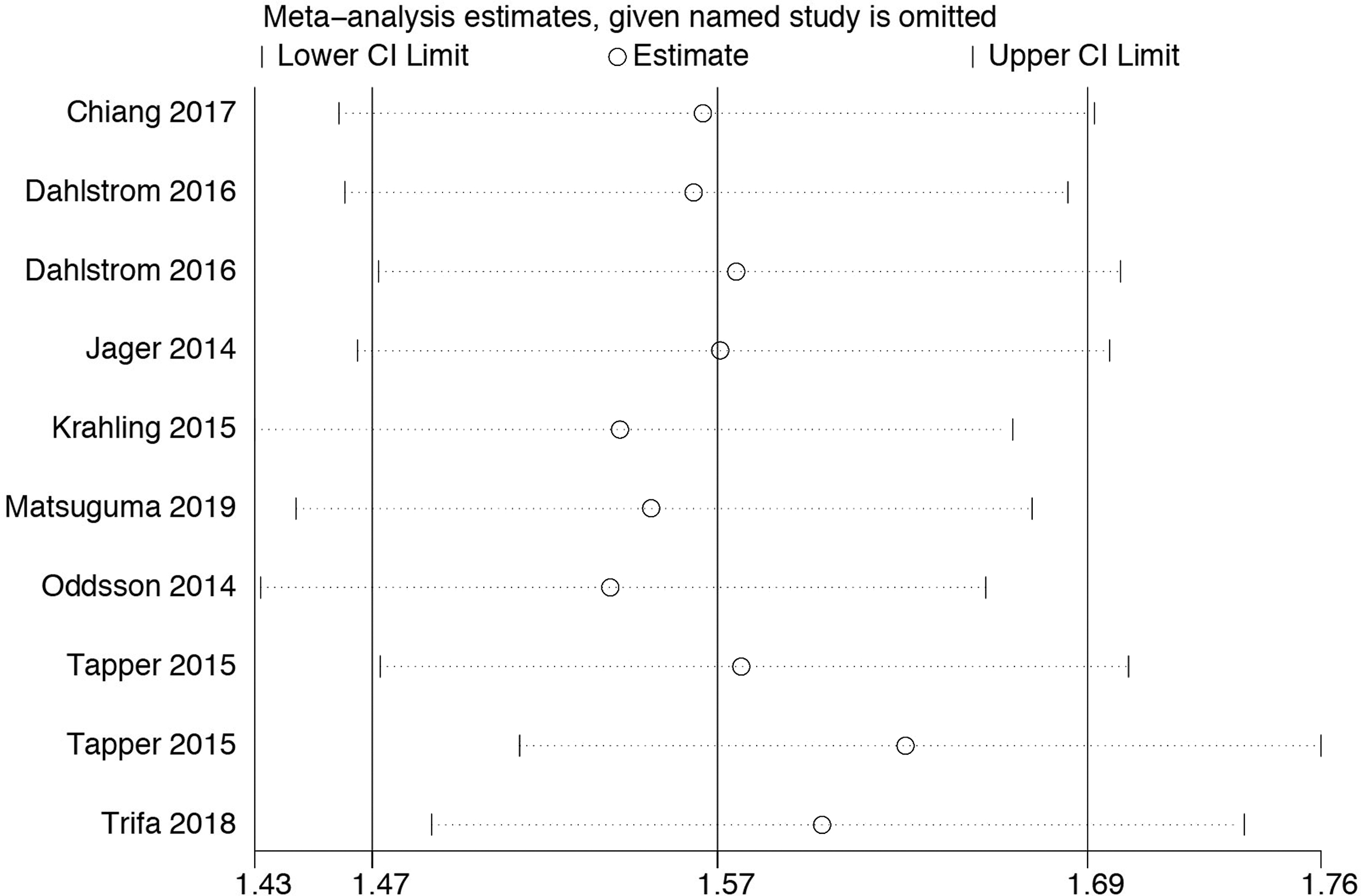
Sensitivity analysis of TERT rs2736100A>C polymorphism and susceptibility of MPNs data.
Publication bias
We applied the funnel plot to evaluate whether a publication bias existed in our meta-analysis. Our funnel plot did not indicate any publication bias in the allele model. And the visual estimation of the funnel plot showed clear symmetry (Fig. 5).
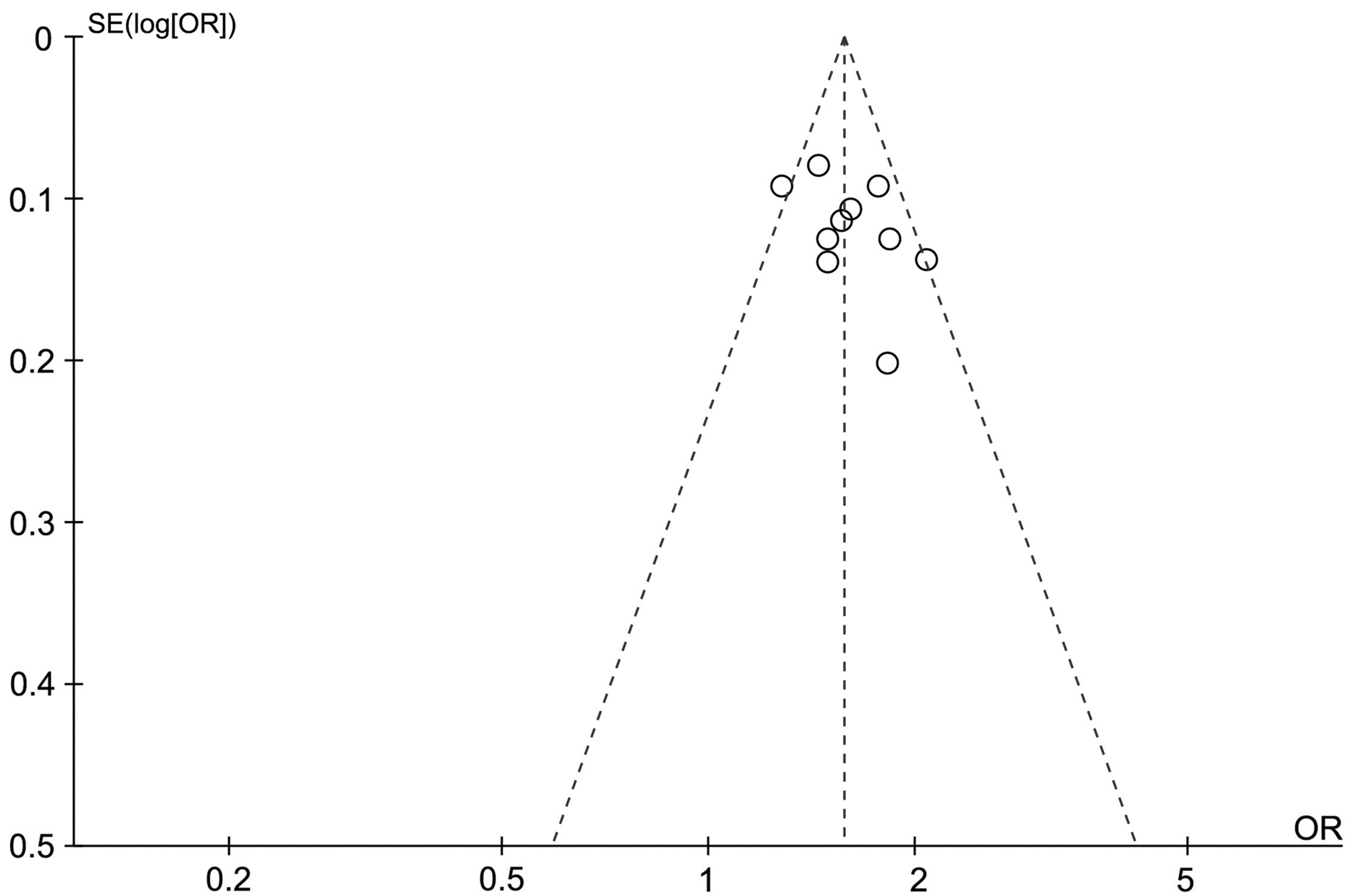
Funnel plot of TERT rs2736100A>C polymorphism and susceptibility of MPNs data for publication bias.
Discussion
Recent studies have reported that TERT rs2736100A>C SNP might be positively associated with cancer susceptibility. This innovative concept provides a new molecular basis for the genetic susceptibility of different malignancies, particularly MPNs (Tapper et al., 2015). However, current results about the association between TERT rs2736100A>C SNP and susceptibility of MPNs are controversial and heterogeneous. Several issues are necessary to be addressed. In this study, we analyze pooled data from eligible case-control studies to confirm the predictive role of TERT rs2736100A>C polymorphism in susceptibility of MPNs. Our results validate that TERT rs2736100A>C allele is significantly related to susceptibility of MPNs in all populations. And the subgroup analysis demonstrates that TERT rs2736100A>C increases the risk of MPNs in all genetic models, and TERT rs2736100 C genotype carriers present a higher risk of MPNs than other genotypes. Thus, TERT rs2736100A allele is a susceptible factor for MPNs.
To our knowledge, telomerase leads telomeres to shorten within each successive round of cell division. When it reached a critical telomere length, the cells enter replicative senescence stage (Wilson et al., 2014). The TERT gene is one of the most important components of telomerase that is located at 5p 15.33 locus (McKay et al., 2008). It encodes the functional unit of telomerase complex that is essential for maintaining telomere length. It has been revealed that TERT overexpression presents in most malignant cells (including MPNs) (Luo et al., 2019), whereas TERT is rarely expressed in normal histiocytes (excluding germ cells and somatic cells with high replication potential). The aberrant TERT expression may directly prompt the dysfunction of telomerase, which has been proved to be highly related to oncogenesis and oncomodulation in different cancers (Hao et al., 2019).
Apart from the impact on telomerase and telomere length, TERT dysregulation can significantly increase cellular activities through activation of different oncogenesis signaling pathways. For example, first, the pRb/E2F signaling pathway is an important pathway mediating cell cycle arrest and is closely related to cell senescence and tumor formation (Xiang et al., 2002). Multiple previous studies have shown that overexpression of TERT can induce hyperphosphorylation of Rb and upregulation of E2F transcriptional activity. Second, regarding Wnt/β-catenin pathway, the extracellular signal is transmitted to the cell through the activation process of the intracellular domain. It has been found that β-catenin protein occupies the TERT promoter, which can lead to TERT overexpression directly (Fernandez and Johnson, 2018). Moreover, TERT interacts with β-catenin to promote the development of tumors by inducing epithelial-mesenchymal transition (Qin et al., 2016). Finally, PI3K/AKT pathway is crucial in cell proliferation and differentiation. The dysregulation of any parts in PI3K/AKT can elicit tumorigenesis. Previous researches have concluded that PI3K/AKT signaling activates transcription of TERT by phosphorylation of sMad1, which will increase the transcription bindings at TERT promoter (Che et al., 2019). Undoubtedly, not only the pivotal relevance between TERT and telomerase but also the biological characteristics of TERT in diversified pathways make it a compelling candidate gene for cancer susceptibility.
As such an important component to telomerase and involved in multiple oncogenesis pathways, the contributive role of TERT variants to cancer susceptibility should be highlighted. Abnormal gene variants play a critical role in the transformation of somatic cells into malignant cells throughout the aberrant activation of telomerase and other related signaling pathways. Diversified variants have been reported to be associated with the risk of cancers, including TERT rs2736100 polymorphism. In 2008, McKay et al. carried out a study about the genetic susceptibility of lung cancer and TERT rs2736100 was detected as a pathogenetic factor. Since then, a growing number of genome wide association studies have shown that TERT rs2736100 polymorphism is associated with the susceptibility of multiple types of cancers, such as glioma, colorectal cancer, testicular germ cell cancer, bladder cancer, and gastric cancer. (Shete et al., 2009; Turnbull et al., 2010; Gago-Dominguez et al., 2011; Zhang et al., 2017).
To probe the underlying mechanism of pathogenesis role of TERT rs2736100 polymorphism, elucidation of the relationship between telomerase activity and TERT rs2736100 polymorphism is of necessity. Many studies reported that telomerase activity is relatively high in the bone marrow of patients with MPN patients. It has been extensively explored that TERT rs2736100A>C is responsible for the activated telomerase and longer telomeres by the direct regulation on TERT expression. This direct regulation might be initiated by increasing the activity of TERT enhancer (Krahling et al., 2016). Intriguingly, it has also been found that telomere shortening is present in myeloma cells of MPN patients, which may result from excessive proliferation of these cells (Bernard et al., 2009). Regardless of the effect of TERT rs2736100A>C on telomerase, various studies have reported that TERT rs2736100A>C is highly associated with aberrant clonal expansion of differentiated hematopoietic cells in bone marrow, which is consistent with the histological characteristics of MPNs (Bouillon et al., 2018).
There are several limitations to this meta-analysis that should be concerned. First, MPNs are mainly classified as PV, PMF, and ET. The effect of TERT rs2736100A>C on different histological types of MPNs may be diverse. It is worthwhile to further explore the relationship between TERT rs2736100A>C on different subtypes of MPNs in future studies. Second, the age and gender factors are not involved in our analysis owing to the lack of available data. To our knowledge, the morbidity and mortality of MPNs can be influenced by multiple factors, especially age and gender. We believe that TERT rs2736100A>C has a distinct effect on the susceptibility of MPNs if we take their age and gender into consideration. Third, we merely focus on the association between TERT rs2736100A>C and susceptibility of MPNs, whereas the concomitant TERT variants (e.g., rs401681 and rs2736098) and variants of other genes (e.g., JAK2, CALR, MECOM, HBS1L1, SH2B3, TET2, CHEK2, and ATM) might have interactive effect with TERT rs2736100A>C on susceptibility of MPNs (Hinds et al., 2016) (Tapper et al., 2015).
Conclusion
In conclusion, the association between TERT rs2736100A>C polymorphism and susceptibility of MPNs is dramatic in all genetic models. TERT rs2736100A>C allele increases the overall risk of MPNs. In the future, further studies are warranted to determine the functional role of TERT rs2736100 polymorphism in MPNs.
Statement of Ethics
All the research meets the ethical guidelines.
Footnotes
Authors' Contributions
T.H. and Q.L. designed the analysis. T.H. and M.L. identified the proper publications and collected the data. Tao pooled the data and performed statistical analysis. M.L. adjusted article format. All authors analyzed the data and revised the article. All authors reviewed and approved the final report.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.