
Abstract
Background:
Chronic heart failure (CHF) is a major health burden worldwide, but there are a lack of effective methods for its early diagnosis and prognostic evaluation. Circular RNAs (circRNA), as a class, have been found to regulate gene expression and are implicated in multiple types of diseases. The circRNA, CDR1as, is reported to regulate gene transcription by acting as a microRNA inhibitor. However, the role of CDR1as in CHF remains to be elucidated. The aim of this study was to investigate the role and mechanism of action of the circRNA, CDR1as, in CHF.
Methods:
A total of 30 patients with CHF and 30 healthy persons were included in the study. The levels of CDR1as, miR-135a, and miR-135b in the plasma of all subjects were quantified by qRT-PCR. ELISA was used to detect the level of HMOX1 in plasma. The effect of CDR1as was investigated in human cardiomyocytes, including HCMs and AC16 cells.
Results:
CDR1as was upregulated in the plasma of patients with CHF and is a potential diagnostic marker of CHF. The levels of miR-135a and miR-135b were downregulated in the plasma of patients with CHF. The plasma level of HMOX1 in patients with CHF was significantly higher compared with the control group and was highly correlated with cardiac function in CHF patients. CDR1as was shown to act as a sponge for miR-135a and miR-135b and regulated the proliferation and apoptosis of human cardiomyocytes through the miR-135a/HMOX1 and miR-135b/HMOX1 signaling axes.
Conclusion:
CDR1as is a potential biomarker of CHF that is mechanistically involved in the disease pathogenesis and participates in regulating the occurrence and development of CHF through the miR-135a/HMOX1 and miR-135b/HMOX1 signaling axes.
Introduction
Chronic heart failure (CHF) is a common complication of cardiovascular disease. The incidence of CHF is increasing year by year, making it a health threat worldwide (Abuzaanona and Lanfear, 2017). Early correct diagnosis and status assessment can help patients select the appropriate therapeutic strategies and reduce mortality, but there is still a lack of effective biomarkers for early diagnosis and prognostic evaluation (Fu et al., 2016).
In eukaryotic cells, protein-coding RNAs (mRNAs) account for ∼2% of the genome, and the rest are noncoding RNAs (ncRNAs) (Dou et al., 2016). ncRNAs are multifunctional RNAs that modulate gene expression at both the transcriptional and posttranscriptional levels (Mattick and Makunin, 2006). Recently, with the application and development of RNA sequencing (RNA-seq) technology, circular RNAs (circRNAs) have been found to be widely present in many biological cells and to regulate gene expression (Salzman et al., 2012). circRNAs are covalently closed RNAs with cell-specific or tissue-specific expression patterns (Sanger et al., 1976; Chen and Yang, 2015) that have been implicated in multiple types of diseases. Unlike linear RNA, circRNAs lack 5′ and 3′ ends, so they are protected from exonuclease cleavage, and are highly stable. In addition, circRNA exhibit specific expression patterns in several types of diseases, making them useful as biomarkers for disease diagnosis and prognosis (Huang et al., 2017; Li et al., 2018b; Wang et al., 2018).
CDR1as gene is located at 139865339-139866824 on human chromosome Xq27.1 (Zhao et al., 2019) and is formed by trans-splicing of the CDR1 gene after transcription. CDR1as exists in the cytoplasmic processing bodies (P-bodies) and regulates gene transcription by acting as a natural microRNA inhibitor (Hansen et al., 2013; Memczak et al., 2013). Overexpression of CDR1as leads to developmental defects in the zebrafish midbrain, and inhibition of miR-7 expression also induces a similar phenotype, whereas overexpression of miR-7 partially rescued these defects (Xu et al., 2015). Li et al. (2018a) showed that knockdown of CDR1as significantly inhibited osteoblast differentiation of periodontal ligament stem cells in vitro and in vivo, which is similar with the changes induced by miR-7 overexpression. As the first identified circRNA with biological function in mammalian cells, CDR1as is conservatively and stably expressed in mammalian cells (Yao et al., 2018). However, the role of CDR1as in the occurrence and development of CHF remains to be elucidated.
MicroRNAs are endogenous single-strand noncoding RNAs of 18-24 nucleotides in length (Lu and Rothenberg, 2018). Recently, studies demonstrated that circRNAs often function as endogenous competitive RNA for miRNAs (miRNA sponges). By binding with miRNAs, circRNAs participate in regulating the expression of critical factors in cellular activities, and is widely involved in regulating the occurrence and development of diseases such as atherosclerosis and pulmonary fibrosis (Burd et al., 2010; Hansen et al., 2013; Memczak et al., 2013; Yao et al., 2018).
Heme oxygenase (HMOX) is a key enzyme that catalyzes the degradation of heme and is present in three isoforms, HMOX1, HMOX2, and HMOX-3. Because carbon monoxide (CO) and bilirubin produced by the HMOX1 degradation of heme play an important role in the process of CHF, HMOX-1 is considered to be involved in the development of CHF (Barbagallo et al., 2013).
In the current study, to identify effective prevention and treatment methods for CHF, we analyzed the levels of CDR1as, its potential target miRNAs, and HMOX1 in the plasma of CHF patients, and explored the possible mechanism of CDR1as in the occurrence and development of CHF through an in vitro cell model.
Materials and Methods
Subjects
From December 2018 to April 2019, 30 patients with CHF and 30 healthy individuals at the Shanghai General Hospital were enrolled in the study. All patients with CHF had chronic and stable symptoms of heart failure (Bank et al., 2012). The inclusion criteria of subjects enrolled in the present study were as follows: (1) subjects were over 18 years of age and had been treated for heart failure for more than 1 month; (2) subjects met the diagnostic criteria for CHF or had left ventricular enlargement and decreased left ventricular ejection fraction; and (3) subjects had underlying diseases that clearly cause CHF. Patients with the following characteristics were excluded from the study: (1) patients suffering from unstable angina pectoris, acute non-ST elevation myocardial infarction, or acute ST elevation myocardial infarction within 1 month before the study; (2) patients with a malignant tumor or a severe infectious disease; (3) patients suffering from acute decompensated heart failure; (4) patients with a history of organ transplant; and (5) patients with blood creatinine over 2.5 mg/mL; (6) patients with immune system diseases; (7) patients with diabetes; and (8) patients with uncorrected valvular heart disease. There were no statistically significant differences in age, sex, body mass index, smoking and drinking, and aspartate aminotransferase or alanine aminotransferase levels in the 30 CHF patients included in the study (p > 0.05). The general information of the enrolled CHF patients and the control group is shown in Table 1. All subjects signed informed consent, and this study was approved by the Medical Ethics Committee of the Shanghai General Hospital.
Comparison of Physical Data Between Chronic Heart Failure Patients and the Control Group
CHF, chronic heart failure; BMI, body mass index; AST, aspartate aminotransferase; ALT, alanine aminotransferase; LVEF, left ventricular ejection fraction; BNP, B-type brain natriuretic peptide; SD, standard deviation.
Cell culture
The HEK-293T, HCM, and AC16 cell lines were purchased from the Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences (Shanghai, China). All cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 mg/mL streptomycin (Gibco-BRL, Grand Island, NY) in a 5% CO2 humidified incubator at 37°C.
Cell transfection
The CDR1as overexpression vector pcDNA-CDR1as, blank control vector (mock), small interfering RNA (si-CDR1as) targeting CDR1as, miR-135a mimic, and miR-135b mimic were chemically synthesized by GenePharma (Shanghai, China).
HCMs and AC16 cells were seeded in 24-well plates at a density of 5 × 104 cells/well 24 h before transfection. When cultured cells reached 50% confluence, DNA and RNA molecule delivery was conducted by Lipofectamine 2000 according to the commercial instructions (Thermo Fisher Scientific, CA). HCM and AC16 cell samples were harvested at 24, 48, 72, and 96 h posttransfection for further experiments.
RNA extraction and quantitative real-time PCR
The plasma was separated from 5 mL of collected peripheral venous blood from subjects and stored at −80°C until use. Total RNA was extracted from subject plasma using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The isolated RNA was reverse transcribed into cDNA using PrimeScript RT Master Mix (TaKaRa, Dalian, China). qRT-PCR was performed by the PrimeScript RT Kit and SYBR Premix Ex Taq (TaKaRa). The primer sequences were as follows: CDR1as forward: 5′-GAC CTG GAG GCC ATT GGA AG-3′, CDR1as-reverse: 5′-AGC AAT TAC TGG TCT TCC ACC A-3′; miR-135a forward: 5′-GGC CTC GCT GTT CTC TAT GG-3′, miR-135a reverse: 5′-GCC ACG GCT CCA ATC CCT AT-3′; miR-135b forward: 5′-TGG CCT ATG GCT TTT CAT TCC T-3′, miR-135b reverse: 5′-AGC TCG CCC CTC ACT GTA-3′; U6 forward: 5′-TGA CAC GCA AAT TCG TGA AGC GTT C-3′, U6 reverse: 5′-CCA GTC TCA GGG TCC GAG GTA TTC-3′; and GAPDH forward: 5′-GGG TGA TGC AGG TGC TAC TT-3′, GAPDH reverse: 5′-GGC AGG TTT CTC AAG ACG GA-3′. The PCR conditions were as follows: 95°C, 30 s; and 95°C, 5 s; 58°C, 34 s for 40 cycles. qRT-PCR was performed on an ABI 7500 real-time PCR system (Applied Biosystems, Foster City, CA). The expression levels of CDR1as, miR-135a, and miR-135b were calculated using the 2−ΔΔCT method. U6 and GAPDH were used as internal controls for miRNA and CDR1as, respectively.
Enzyme-linked immunosorbent assay
A human HMOX1 ELISA Kit (ENZO Life Sciences, Plymouth Meeting, PA) was used to detect HMOX-1 levels in plasma samples according to the user's manual. The concentration of HMOX-1 was determined by the standard curve method, and each sample measurement was repeated three times.
MTT assay
Cell growth was measured after adding 0.5 mg/mL MTT solution (Sigma-Aldrich, Merck KGaA) to HCMs and AC16 cells harvested at the indicated time points. After 4 h of incubation, the medium was changed to 100 μL of dimethyl sulfoxide (DMSO; Sigma-Aldrich; Merck KGaA), and the absorbance was recorded at 450 nm (OD450) using microplate readers.
Dual luciferase assay
The constructs, pGL3-CDR1as-WT (with miR-135a binding site), pGL3-CDR1as-MUT, pGL3-CDR1as-wt (with miR-135b binding site), and pGL3-CDR1as-mut, were synthesized by GenePharma. Experimental compounds were transfected into HEK-293T cells using Lipofectamine 2000. Cells were harvested at 48 h posttransfection for the dual luciferase reporter assay following the manufacturer's instructions.
Apoptosis assay
HCMs and AC16 cells were collected 24 h after transfection. The Annexin V-FITC Apoptosis Detection Kit (Beyotime, Shanghai, China) was used to detect apoptosis according to the user's manual. Annexin V and PI staining was performed on HCMs and AC16 cells collected at 24 h posttransfection. The apoptosis rate was detected by flow cytometry (FACScan; BD Biosciences, San Jose, CA).
Western blot analysis
The concentration of isolated protein was quantified using the BCA Protein Assay Kit (Thermo Scientific, Rockford, IL). Total protein was separated in 15% SDS-PAGE gels and transferred onto PVDF membranes for protein detection. Subsequently, the PVDF membranes were blocked with 5% nonfat milk for 2 h at room temperature and then incubated with the corresponding primary antibodies (anti-HMOX1: ab137749; anti-β-actin: ab8226; Abcam, Cambridge, MA) overnight at 4°C. The protein level of β-actin was used as a loading control. Protein levels were visually recorded by an enhanced chemiluminescence (ECL) reagent (Thermo Fisher Scientific).
Statistical analysis
Statistical analysis in this study was carried out using SPSS version 22.0 (SPSS, Inc., Chicago, IL). Continuous variables are expressed as the mean ± SD, while categorical variables are expressed as n (%), and Student's t-test and chi-square test were used for the statistical analysis, respectively. The Spearman's correlation coefficient was adopted to evaluate the correlation among CDR1as, miR-135a, and miR-135b, and the relationship among miR-135a, miR-135b, and plasma HMOX1 levels. The receiver operating characteristic curve (ROC) was used to determine the diagnostic value of CDR1as, miR-135a, and miR-135b in CHF. p < 0.05 indicated a significant difference.
Results
CDR1as was upregulated in the plasma of CHF patients
The levels of plasma CDR1as in enrolled subjects were determined by qRT-PCR, and the results showed that the plasma CDR1as levels in CHF patients were significantly higher than those in the control group (p < 0.05, Fig. 1A). In addition, the higher the levels of CDR1as in the plasma of CHF patients, the higher the NYHA class of the patients (p < 0.001, Fig. 1B). Correlation analysis showed that plasma CDR1as levels were negatively correlated with LVEF (r = −0.90; p < 0.001) but positively correlated with plasma BNP levels (r = 0.85; p < 0.001) (Fig. 1C, D). The area under the ROC was 0.84 (p < 0.001), suggesting that the level of plasma CDR1as could be used as a potential diagnostic marker of CHF (Fig. 1E). In addition, there was no significant difference in plasma levels of CDR1as between males and females in CHF patients and controls group (p > 0.05, Supplementary Fig. S1).
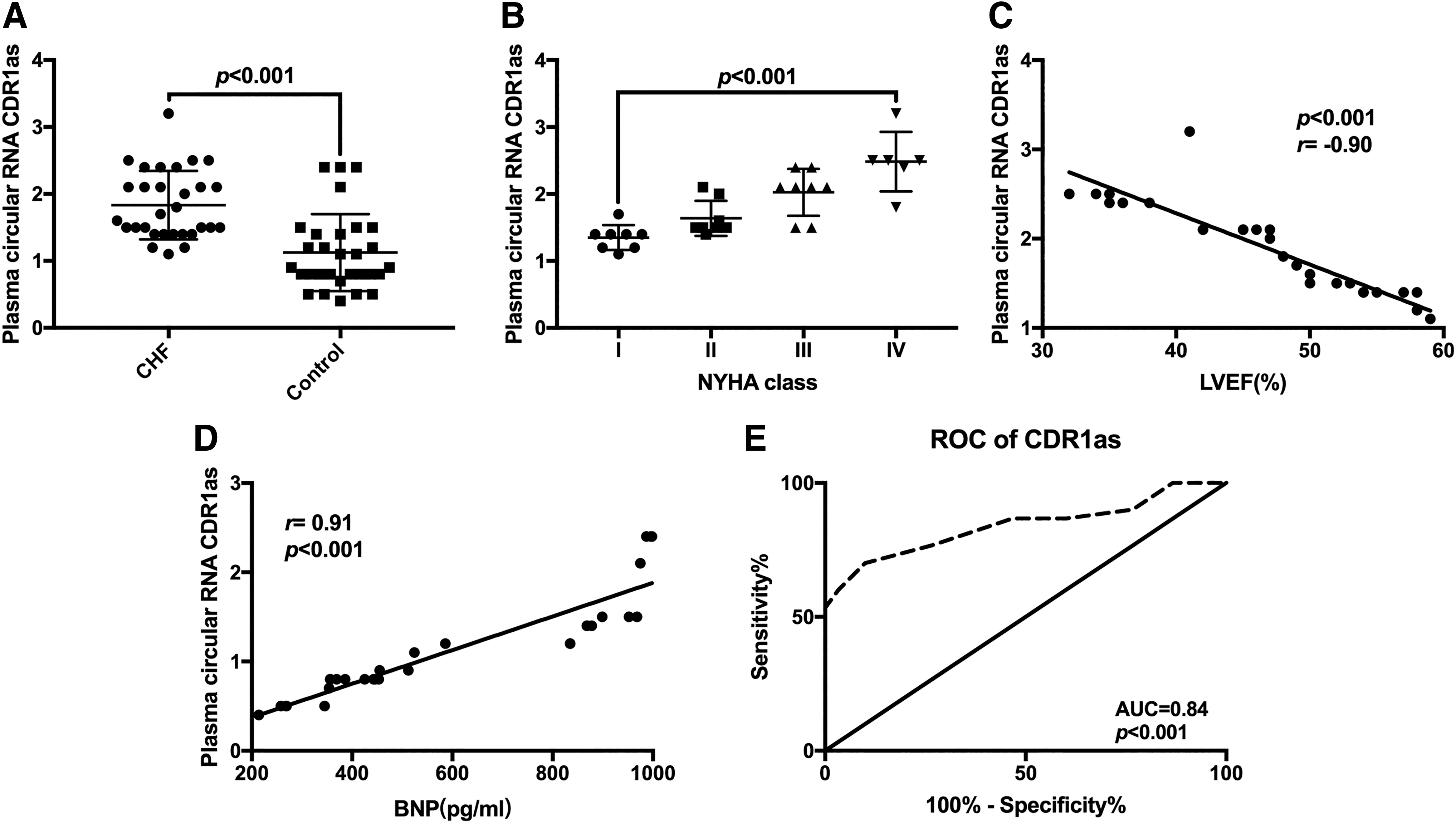
CDR1as was upregulated in the plasma of CHF patients.
The levels of miR-135a and miR-135b were decreased in the plasma of CHF patients
The plasma from patients with CHF (n = 30) and controls (n = 30) was collected to quantify the levels of miR-135a and miR-135b. As shown in Figure 2A and B, the expression of miR-135a and miR-135b was downregulated in the plasma from patients with CHF. The higher the levels of miR-135a and miR-135b in plasma, the lower the NYHA class of CHF patients (Fig. 2C, D). Similarly, there was a negative correlation between the levels of miR-135a and miR-135b in plasma and LVEF (r = 0.55, r = 0.69; p < 0.001, p < 0.001) and the amount of BNP in plasma (r = −0.83, r = −0.55; p < 0.001, p < 0.001) (Fig. 2E-H). The ROC showed that the AUCs of miR-135a and miR-135b were 0.82 (95% CI: 0.71-0.93, p < 0.01) and 0.66 (95% CI: 0.52-0.80, p < 0.01), respectively, which indicated that the expression levels of miR-135a and miR-135b in plasma are potential markers for the diagnosis of CHF (Fig. 2I).
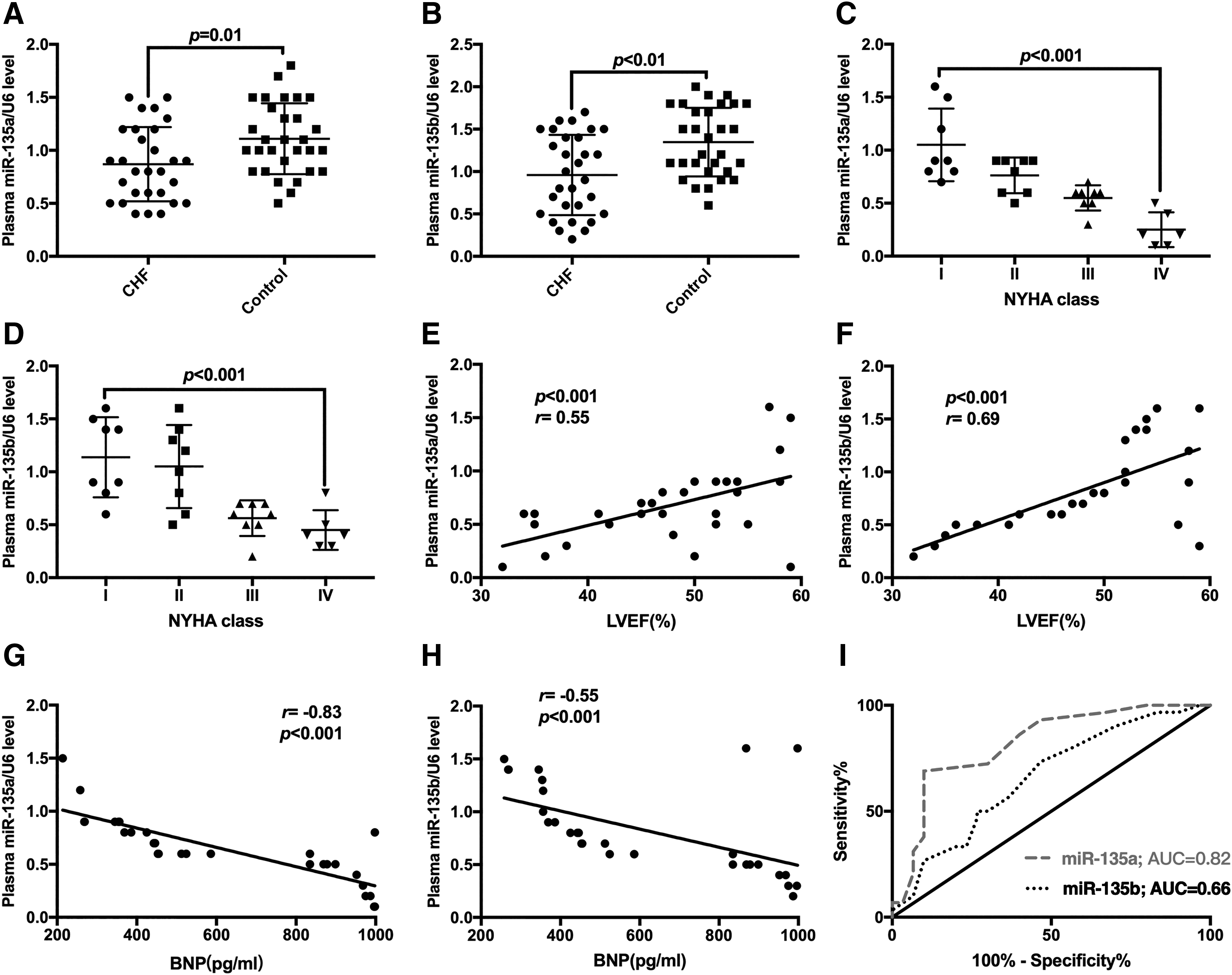
The expression of miR-135a and miR-135b was reduced in the plasma of patients with CHF.
HMOX1 was highly expressed in the plasma of patients with CHF
ELISA analysis revealed that the level of HMOX1 in the plasma of CHF patients was significantly higher compared with the control group (p < 0.05) (Fig. 3A). In addition, the higher the NYHA class, the higher the plasma level of HMOX1 in CHF patients (p < 0.01) (Fig. 3B). As presented in Figure 3C and D, there was a negative correlation between plasma HMOX1 level and LVEF (r = −0.64, p < 0.01), but a positive correlation with BNP levels (r = 0.81, p < 0.001). The ROC showed that the AUC of HMOX1 was 0.85 (95% CI: 0.76-0.95, p < 0.01), implying that plasma HMOX1 levels were a potential biomarker for the diagnosis of CHF (Fig. 3E).
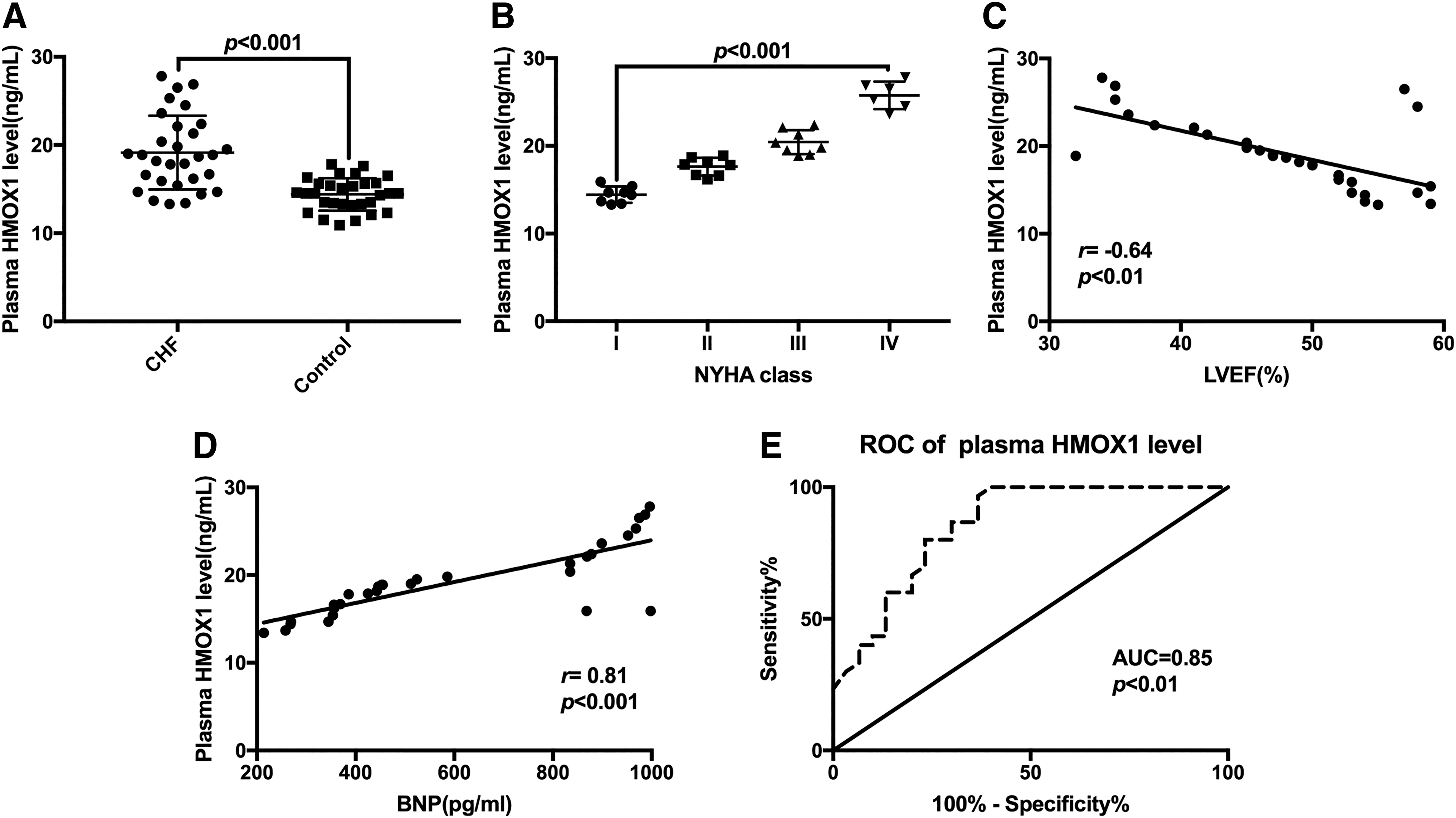
The level of plasma HMOX1 in CHF patients was significantly increased.
CDR1as acted as a sponge for miR-135a and miR-135b
As shown in Figure 4A and B, there was a significant negative correlation between CDR1as levels and miR-135a or miR-135b levels in patients with CHF (r = −0.72, r = −0.88; p < 0.01, p < 0.01). Moreover, the expression level of CDR1as positively correlated with HMOX1 levels in the plasma of CHF patients (r = 0.84; p < 0.01, Fig. 4C). Additionally, the HMOX1 level was significantly negatively correlated with miR-135a and miR-135b (r = −0.57, r = −0.67; p < 0.01, p < 0.01, Fig. 4D, E).
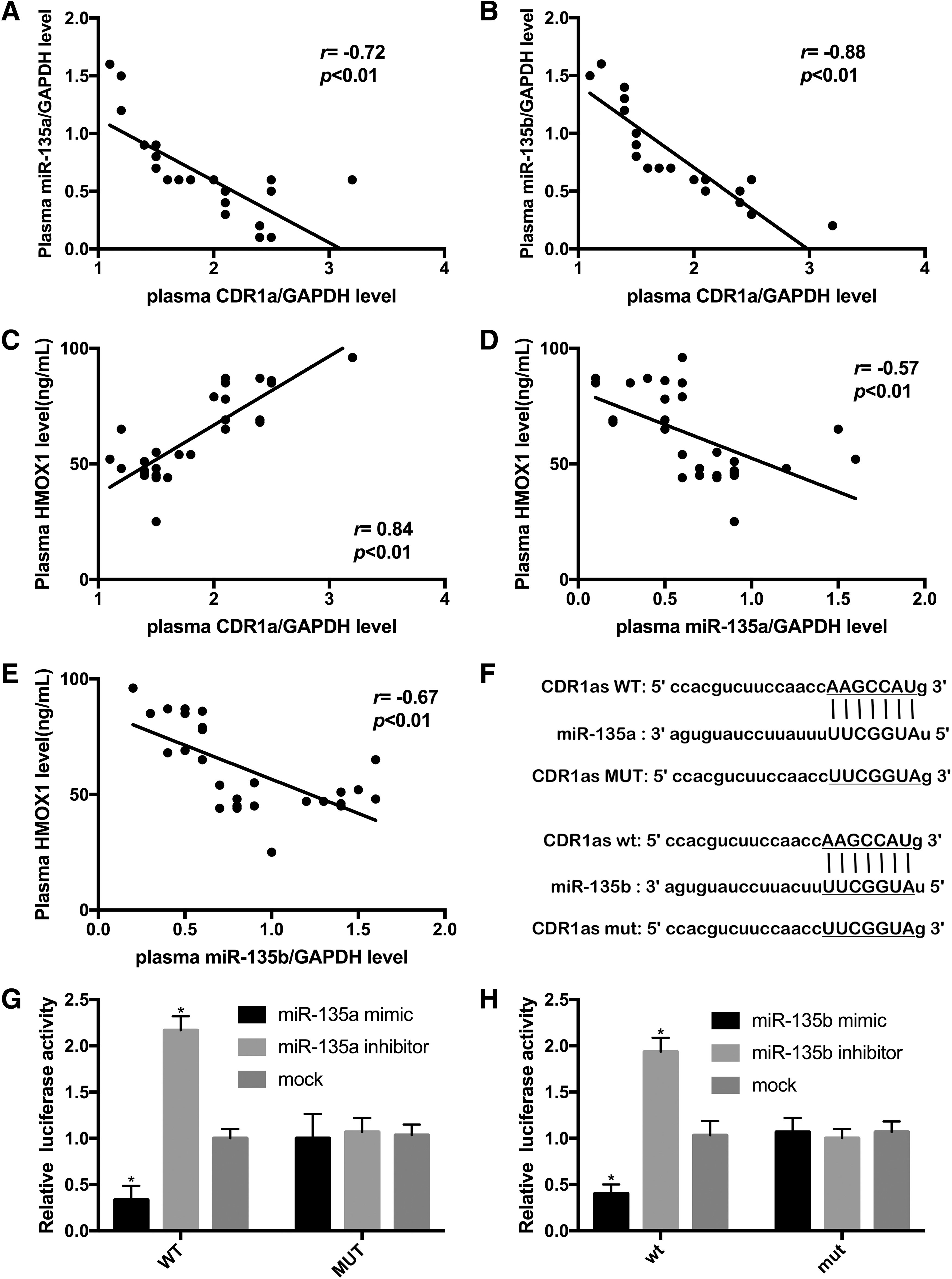
CDR1as acted as a molecular sponge for miR-135a and miR-135b.
Bioinformatic analysis predicted that CDR1as contains the seed sequences of miR-135a and miR-135b (Fig. 4F). To further characterize the binding between CDR1as, miR-135a, and miR-135b, dual luciferase reporter assays were conducted. As expected, cotransfection of CDR1as-WT and miR-135a mimic and cotransfection of CDR1as-wt and miR-135b mimic significantly reduced the luciferase activity. The addition of miR-135a and miR-135b to cells transfected with CDR1as-WT or CDR1as-wt greatly increased the luciferase activity (Fig. 4G, H). Taken together, these results demonstrated that CDR1as was a molecular sponge for miR-135a and miR-135b.
CDR1as regulated the proliferation of human cardiomyocytes through the CDR1as/miR-135a/HMOX1 axis and CDR1as/miR-135b/HMOX1 axis
Based on the above results, we speculated that CDR1as may be involved in the development of CHF through the miR-135a/HMOX1 and miR-135b/HMOX1 signaling axes. The MTT assay showed that compared with the mock group, overexpression of CDR1as markedly promoted the proliferation of HCMs and AC16 cells, whereas knockdown of CDR1as inhibited the growth of HCMs and AC16 cells. Notably, ectopic transfection of miR-135a mimic and miR-135b mimic restored the effect of CDR1as on the proliferation of HCMs and AC16 cells (Fig. 5A, B). Similarly, the protein level of HMOX1 in HCMs and AC16 cells was increased by overexpression of CDR1as, but transfection of miR-135a and miR-135b reversed the effect of CDR1as on the HMOX1 expression (Fig. 5C, D).
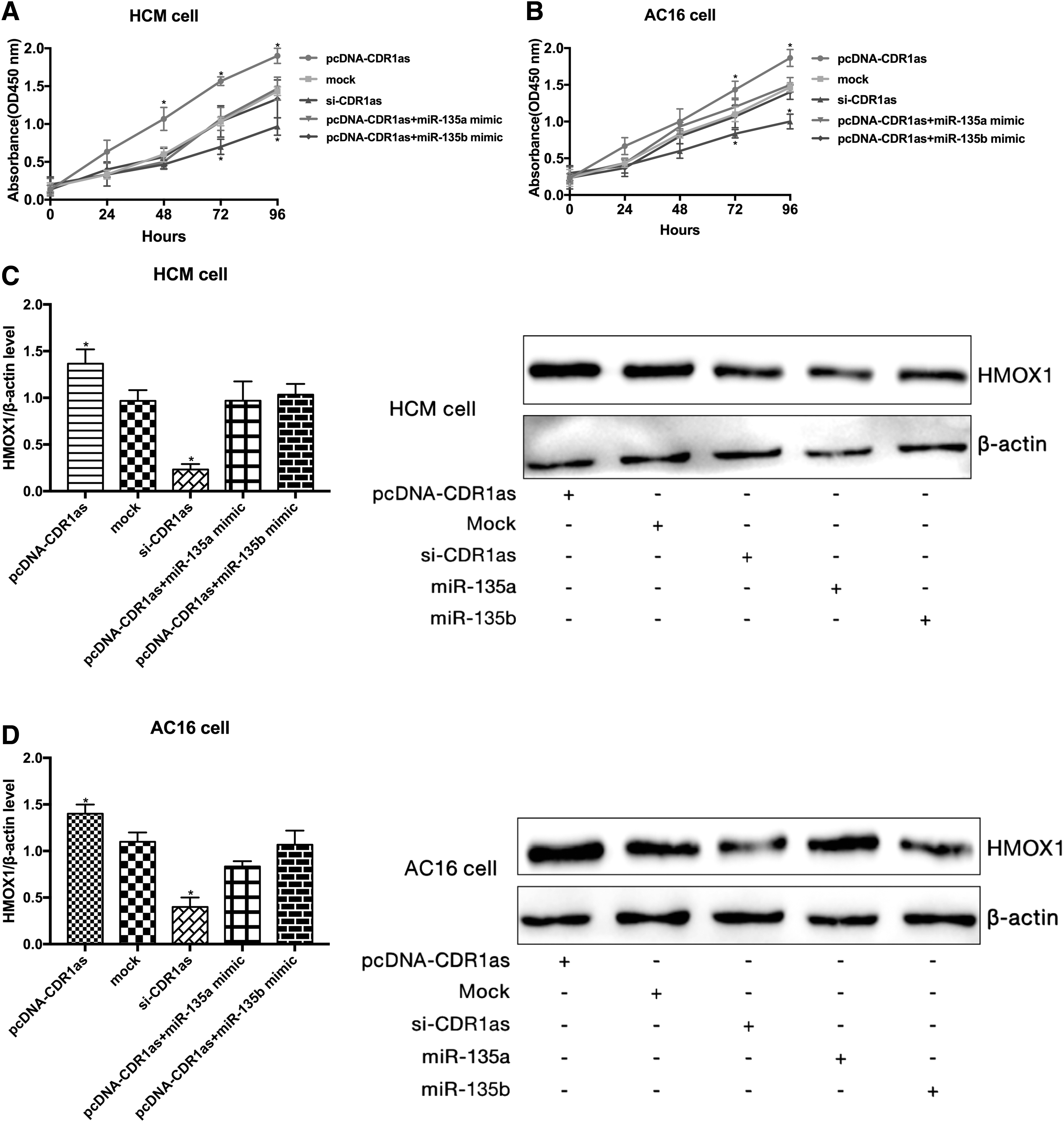
CDR1as modulated the proliferation of human cardiomyocytes.
CDR1as regulated human cardiomyocyte apoptosis through the miR-135a/HMOX1 and miR-135b/HMOX1 signaling axes
CDR1as overexpression significantly inhibited HCM and AC16 cell apoptosis, and knockdown of CDR1as significantly promoted HCM and AC16 cell apoptosis. In addition, miR-135a mimic or miR-135b mimic transfection reversed the effect of CDR1as overexpression on the apoptosis of HCM and AC16 cells (Fig. 6A, B). As shown in Figure 6C and D, ectopic expression of CDR1as suppressed the endogenous protein levels of Bax and Bcl-2, and miR-135a mimic or miR-135b mimic could restore the effect of CDR1as overexpression on Bax and Bcl-2 protein expression in HCMs and AC16 cells.
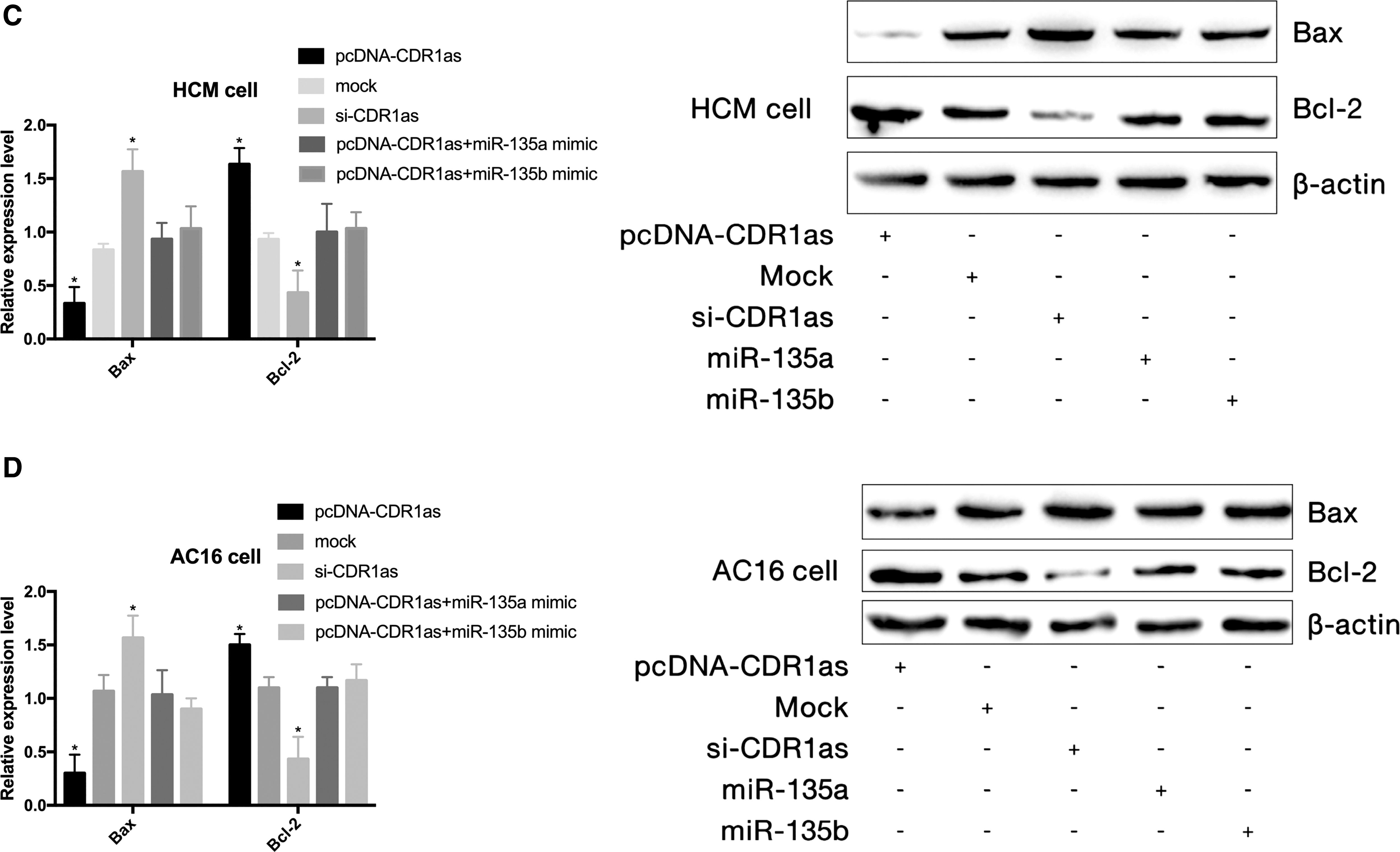
CDR1as is involved in the regulation of human cardiomyocyte apoptosis.
Discussion
In this study, we found that the level of CDR1as was upregulated in the plasma of patients with CHF and correlated with NYHA class, suggesting that CDR1as could be used as a potential diagnostic marker of CHF. Through bioinformatic analyses, we found that CDR1as and the 3′ untranslated region (UTR) of HMOX1 contained target sites of miR-135a and miR-135b. Dual luciferase assays showed that CDR1as could interact with miR-135a and miR-135b to function as a molecular sponge for miR-135a and miR-135b in HCMs and AC16 cells. Finally, with functional experiments, we proved that CDR1as exerted a regulatory effect on the proliferation and apoptosis of HCMs and AC16 cells through the miR-135a/HMOX1 and miR-135b/HMOX1 axes.
Growing evidence indicates that circRNA can affect gene expression through different pathways, thereby regulating the occurrence and development of diseases (Qu et al., 2015; Salzman, 2016; Ebbesen et al., 2017). In this study, we found that circRNA CDR1as was highly expressed in the plasma of patients with CHF. ROC analysis showed that the AUC of plasma CDR1as levels was as high as 0.84, suggesting that CDR1as in plasma is a potential biomarker for CHF diagnosis.
Through complementary binding with the 3′UTR of their target mRNA molecules, microRNAs mediate target mRNA degradation or translational inhibition to negatively regulate the biological effects dominated by the target gene (Tutar, 2014; Lu and Rothenberg, 2018).
MicroRNAs are involved in many important cellular pathways and pathophysiological processes (Fabian et al., 2010; Zhang et al., 2017). In this study, bioinformatics analysis predicted that CDR1as contained target binding sites of miR-135a and miR-135b. Further dual-luciferase assays demonstrated that CDR1as was a molecular sponge for both miR-135a and miR-135b. In addition, miR-135a and miR-135b were downregulated in the plasma of CHF patients. Combined with the AUC values of miR-135a and miR-135b in the ROC, these results indicate that miR-135a and miR-135b levels in plasma are potential diagnostic markers of CHF.
HMOX1 is a key enzyme in the process of heme metabolism. HMOX1 is expressed at low levels under physiological conditions. However, inflammatory factors, oxidative stress, ultraviolet radiation, ischemia, hypoxia, and heme can greatly promote the expression of HMOX1 (Moreno-Navarrete et al., 2017; Waza et al., 2018). HMOX1 and its metabolites have the effects of expanding blood vessels, regulating cell proliferation, antioxidative stress, anti-inflammation, and antiapoptosis (Brouard et al., 2000; Lee and Chau, 2002). In the present study, we found that HMOX1 was highly expressed in the plasma of CHF patients, and the amount of HMOX1 was negatively correlated with cardiac function. Taken together with the analysis of ROC, these results indicated that HMOX1 could be used as a potential biomarker for CHF diagnosis.
HMOX1 was predicted to be a potential target gene of miR-135a and miR-135b. Therefore, we speculated that CDR1as may participate in the occurrence and development of CHF by regulating the miR-135a/HMOX1 and miR-135b/HMOX1 axes. Experimental results showed that overexpression of CDR1as not only elevated HMOX1 expression but also promoted proliferation and inhibited apoptosis of HCMs and AC16 cells. Notably, miR-135a or miR-135b mimic transfection obviously reversed the effect of CDR1as overexpression on HMOX1 protein levels and the proliferation and apoptosis of HCMs and AC16 cells. The above data suggested that CDR1as regulated the proliferation and apoptosis of human cardiomyocytes through the miR-135a/HMOX1 and miR-135b/HMOX1 signaling axes and may be involved in the occurrence and development of CHF.
We preliminarily investigated the role of CDR1as in CHF in the present study. CDR1as, miR-135a, miR-135b, and HMOX1 were identified as potential biomarkers of CHF diagnosis. CDR1as may regulate the occurrence and development of CHF through the miR-135a/HMOX1 and miR-135b/HMOX1 signaling axes, but this conclusion needs to be validated further in vivo. Additionally, the study had the following limitations. Taking the plasma CDR1as levels of the CHF patients and controls as a reference, the minimum sample sizes required for CHF patients and control groups were 9 and 9, respectively. Obviously, the 30 CHF patients and 30 control groups selected in this study was in line with the requirements. But the sample size of 30 cases is still small, which may have impacted the objectivity of the statistical analysis. Moreover, there may be multiple circRNAs involved in the occurrence of CHF, and further research is needed to explore novel regulatory signaling axes. Finally, due to the role of CDR1as, miR-135a, and miR-135b in cardiomyocyte apoptosis, the specific role of CDR1as, miR-135a, and miR-135b in mitochondrial dysfunction will be further explored in the future.
Authors' Contributions
Cheng Chen and Hua Shen designed and performed the experiments and analyzed the data. Qiuping Huang and Qing Li participated in the clinical data collection and analysis. Cheng Chen was responsible for writing the article. All the authors approved the final article before submission.
Author Disclosure Statement
There are no competing interests associated with the article.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.