
Abstract
Background:
Allergic diseases, including asthma, atopic dermatitis, allergic rhinitis, and food allergies, are caused by both environmental and genetic factors. The allergic condition, where genetic factors make up the largest proportion (up to 95%), is asthma.
Aim:
To identify polymorphisms and mutations in potentially disease-causing genes in a family affected with allergic asthma.
Methods:
Whole exome sequencing of the index patient was performed via next-generation sequencing. Variants in known allergy-associated susceptibility genes were identified by comparison with the reference genome GRChr37.
Results:
Seven common polymorphisms and three rare mutations were identified in the allergy-susceptibility genes of the index patient. Only four of these variants co-segregated with a second patient in the same family. These variants occurred in the TENS1, NPSR1, RAD50, and IL6R genes.
Discussion:
The variants observed in TENS1 and NPSR1 are relatively common (minor allele frequency, MAF ∼0.4), whereas the mutation in RAD50 is rare (MAF 0.0035). The mutation identified in IL6R (S409P) has never been found before. IL6R encodes an important receptor of the inflammatory system. The mutation occurs in the intracellular domain within a tyrosine-based motif, which is required for sorting of the IL6R protein to the basolateral side of polarized cells. It is likely that this rare mutation contributes—together with the other variants—to the predisposition to asthma and other allergic diseases.
Introduction
The incidence of allergic diseases has dramatically increased in the industrialized world during the past decades. The term allergy defines a hypersensitive inflammatory immune response to normally harmless environmental substances and includes allergic asthma, atopic dermatitis (eczema), allergic rhinitis (hay fever), and food allergy. It is generally accepted that these atopic hypersensitivities are mediated by antibodies of the immunoglobulin E (IgE) class. The development of an allergy is a very complex process and involves both environmental and genetic contributors. Today it is estimated that the contribution of a genetic predisposition makes up, depending on the individual study, 35-95% for asthma, 33-91% for allergic rhinitis, and 71-84% for atopic dermatitis (Ober and Yao, 2011; Portelli et al., 2015). It has also been realized that asthma, allergic rhinitis, and atopic dermatitis represent clinically related diseases. Thus, the majority of people with asthma also have allergic rhinitis (∼80%) and many patients with allergic rhinitis also suffer from asthma (19-38%).
Several approaches have been used to identify genes that might predispose a person to an allergic disease (Ober and Yao, 2011; Portelli et al., 2015). (1) Candidate gene association studies search for an enrichment of single nucleotide polymorphisms (SNPs) or other variants in patients with a disease compared with normal controls. For this approach, genes are selected based on their known function and putative involvement in the pathogenesis. Naturally, no novel genes or pathways can be detected by this approach. (2) Genome-wide linkage studies analyze the co-segregation of a disease locus in families with several affected members. The likelihood of co-segregation is expressed as LOD score and summed up across families. To achieve statistical significance, this approach requires large sample sizes. Moreover, the approach has a poor resolution since the disease locus usually spans a broad region on a chromosome containing several dozens of genes. (3) Genome-wide association studies (GWASs) extend the approach described earlier by analyzing a high number of SNPs from a large cohort of patients represented on a high-throughput genotyping platform as they are found on microarrays provided by Affimatrix or Illumina. Although this approach has a good resolution, it will only allow the identification of common risk alleles. In fact, it has been noticed that the identified polymorphisms can explain only a small proportion of the heritability of asthma and other allergies, a phenomenon that was termed “missing heritability” (Ober and Yao, 2011). One explanation might be that the data sets used for GWAS do not include very rare variants (mutations that occur with a frequency <5%) or structural variants of genes with different copy numbers. (4) Next-generation sequencing is used to identify and resequence rare variants associated with allergies. It is thought that rare variants might have a larger impact than common variants on the risk to acquire an allergy (Cirulli and Goldstein, 2010). Together, the four approaches have fostered the idea that asthma and other atopic allergies might be caused by one or several rare mutations in a genetic background of several common risk alleles that predispose to the disease.
In this study, we used next-generation sequencing to identify common polymorphisms and rare variants in candidate genes, which have previously been identified by the approaches listed earlier, in a family affected with allergic asthma.
Materials and Methods
Genome analysis
Genomic DNA was isolated from peripheral blood leukocytes using the Wizard genomic DNA purification kit from Promega (Madison, WI). Written consent was obtained from all probands to use the samples for genetic testing and to publish the results in accordance with the guidelines of the Swiss Society of Medical Genetics. The DNA was fragmented with transposase and provided with dual-indexed adapters. A library was prepared with the SureSelect QXT V5 kit from Agilent. The library was sequenced on an Illumina HiSeq 2500 platform performing 2 × 125 cycles. Paired sequence reads were mapped on the human reference genome GRCh37.75 using the Burrows-Wheeler Alignment Tool (Li and Durbin, 2010). Variant calling was performed with the program LoFreq-Star 2. Finally, the variants were filtered against the reference library of humans with no medical impact (ftp://ftp.ncbi.nlm.nih.gov/pub/clinvar/vcf_GRCh37/) and analyzed with the Integrative Genomics Viewer 2.3.92. All variants were verified by Sanger cycle sequencing on an ABI 3730 platform. Further analysis of the sequences was performed with the program package MacVector 17 (Apex, NC).
Cloning of mutated IL6R
Total RNA was prepared from leukocytes of the index patient with the QIAamp RNA Blood Mini kit from Qiagen (Hilden, Germany). The RNA was transcribed into cDNA using random hexamer primers and Improm-II Reverse Transcriptase from Promega. The full-length sequence for IL6R was amplified by polymerase chain reaction through 35 cycles (20′′ at 98°C, 15′′ at 65°C, 40′′ at 72°C) with KAPA high-fidelity DNA polymerase (Roche Diagnostics, Switzerland) employing two synthetic primers, TGC
Expression in cell culture
Wild-type and mutated cDNA constructs were transfected into HEK293 cells (CRL-1573, American Type Culture Collection, Manassas, VA) using Lipofectamine and Opti-MEM (Invitrogen, San Diego, CA). Transfected cells were grown under an atmosphere of 5% CO2 in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum, nonessential amino acids, penicillin (100 U/mL), and streptomycin (100 mg/mL). For induction of signaling, the cells were starved for 4 h in medium without any serum and then stimulated for 30 min with recombinant human IL6 (200-06; PeproTech Ltd., London, UK). After stimulation, the cell monolayers were directly dissolved in hot sodium dodecyl sulfate (SDS) sample buffer and the solubilized proteins were resolved on a 12.5% SDS polyacrylamide gel. The separated polypeptides were transferred to a nitrocellulose membrane by semidry blotting (TransBlot SD, BioRad). Unspecific sites of the membrane were blocked with 3% milk powder in phosphate buffered saline before the membrane was reacted with a rabbit monoclonal antibody against phospho-STAT3 (D3A7; Cell Signaling Technology, Inc., Danvers, MA). This antibody specifically recognizes phosphorylated Tyr-705 in human STAT3 (79.86 kDa). After three washing cycles, the membrane was incubated with alkaline phosphatase-conjugated secondary antibodies against rabbit IgG (Jackson Laboratories, West Grove, PA). Bound antibodies were visualized by reaction with 5-bromo-4-chloro-3-indolyl-phosphate and nitro blue tetrazolium substrate. To demonstrate equal loading, the blot was subsequently stained with a mouse monoclonal antibody against human GAPDH (Thermo Fisher Scientific).
Results
A family with two patients suffering from allergic asthma was analyzed in this study. The index patient II:3 (Fig. 1) who is currently 62 years old had asthma since his early childhood. Nocturnal asthma was treated with Oxis and Pulmicort, and sudden attacks with Ventolin. The patient also suffered from allergic rhinitis with reaction against house dust, mold, and hazelnut, and was treated with Nasacort. Furthermore, he showed mild psoriasis on elbows, knees, and back, which was treated with Dermovate. Intestinal involvement was noticed with intolerance of gluten and lactose. IgE was elevated in the blood. The patient did not suffer from comorbidities and did not show other symptoms typical of an autoimmune disease such as fatigue, joint or muscle pain, intermittent fever, or weight loss. It is likely that he had inherited the predisposition to allergies from his deceased father I:1 (Fig. 1) who suffered from urticaria and less clearly defined allergies (sneezing in the morning, skin problems, and intestinal involvement). The son III:3 of the index patient had severe asthma since early childhood and suffered from psoriasis and intestinal problems. None of the other relatives showed any signs of asthma, but the younger brother of the index patient complained about strong allergic rhinitis and intestinal problems (Fig. 1).

Pedigree of the family with two asthma patients. The index patient II:3 and his son III:3, who suffer from atopic asthma and other allergies, are indicated by black boxes. It is likely that the index patient has inherited the predisposition to allergies from his deceased father who suffered from several less clearly described allergic conditions, as indicated by the gray boxes. The brother II:5 of the index patient does not have asthma but suffers from severe allergic rhinitis and intestinal problems. Circles, females; square boxes, males; diagonal slash, deceased; white symbols, not affected.
To investigate the putative cause of these allergic diseases, whole exome sequencing (WES) was performed with genomic DNA from the index patient. A representative library of exonic sequences was prepared with SureSelect V5 from Agilent and analyzed by next-generation sequencing on an Illumina platform. A total of 30,673,483 reads were obtained representing 3824 Mb of genomic sequence. Mapping on the human reference genome GRCh37 showed that 65.3% of the sequence reads mapped on targets with only 3.98% duplicates. Sequence variants were called with the program LoFreq-Star, which identified 39,308 SNPs and 2944 InDels. Filtering against a library of all known variants with no medical impact reduced the number to 8826. Special attention was paid to genes that have previously been identified as risk alleles for asthma (Ober and Yao, 2011; Portelli et al., 2015), atopic dermatitis (Kiyohara et al., 2008), psoriasis (Tsoi et al., 2012), allergic rhinitis (Andiappan et al., 2013), inflammation (Milhavet et al., 2008), and inflammatory bowel disease (Huang et al., 2017). In this way, nearly 250 allergy-susceptibility genes were individually inspected with the Integrative Genomics Viewer 2.3.92 for polymorphisms and InDels in exons and flanking regions. Finally, our procedure yielded a list of seven common (CTLA4, TNS1, TLR1, IL13, ADRB2, NPSR1, and IL4R) and three rare missense variants (IL6R, RAD50, and NLRP7) as shown in Table 1.
Variants Found in the Index Patient
MAF, minor allele frequency; SNP, single nucleotide polymorphism.
The relatives of the index patient were checked for the presence of the same variants. This analysis demonstrated that only the mutations in IL6R and RAD50 as well as the polymorphisms in NPSR1 and TNS1 were shared by the son of the index patient, who also suffered from allergic asthma (Table 2). The variants in TNS1 (minor allele frequency, MAF 0.346) and NPSR1 (MAF 0.446) are common, but the variant in NPSR1 occurred in homozygous form in both the index patient and the son. The mutation in RAD50 is rare (MAF 0.0035) and the mutation in IL6R has never been described before (MAF <0.000008). The latter mutation occurs in the intracellular domain of IL6R and leads to an amino acid substitution in the sequence 406-Pro-Pro-Tyr-Ser/Pro-Leu-Gly-Gln-412 (Fig. 2). This sequence is known to act as signal for sorting of IL6R to the basolateral side of polarized cells (Martens et al., 2000).

Sequence of the rare mutation in IL6R. The mutation identified by whole exome sequencing was verified by Sanger cycle sequencing. Part of the original chromatogram is displayed together with the genomic sequence and the predicted amino acid sequence. The heterozygous mutation is indicated by an arrow and reveals a T > C transition, which leads to the substitution of serine by proline in the intracellular domain of IL6R.
Segregation of Variants in the Family
The signaling activity of mutated IL6R was tested in a cell culture experiment employing phosphorylated STAT3, the major downstream target of IL6R, as a measure for signaling. For this experiment, the cDNA sequences for the wild-type and the mutated receptor were cloned into an expression vector and transfected into HEK293 cells. After stimulation of the cells with recombinant IL6, the cell layer proteins of the cultures were dissolved in SDS sample buffer, separated on a polyacrylamide gel and transferred to a nitrocellulose membrane. The membrane was stained with antibodies against phosphorylated STAT3 (Fig. 3). Before stimulation, the cells were found to contain very low levels of phosphorylated STAT3 that did not yield any band on the Western blot (not shown). After stimulation with 30 nM of the ligand, a band of 80 kDa was observed, which is characteristic of phosphorylated STAT3. The intensity of this band became stronger when the cells were stimulated with 100 nM IL6. A similar band of 80 kDa was observed when the experiment was performed with cells, which had been transfected with the mutated receptor. Comparison of band intensities did not reveal significant differences between samples containing the wild-type receptor and those containing the mutated receptor (Fig. 3). These results demonstrate that the mutated receptor is capable of transmitting the IL6 signal to the interior of the HEK293 cells with an efficiency similar to the wild-type receptor.
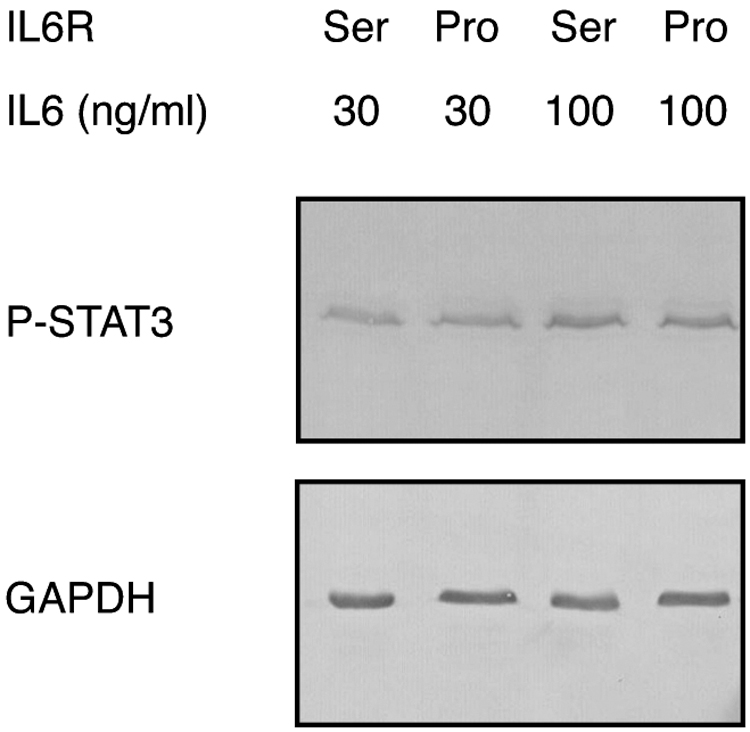
Signaling by mutated IL6R. HEK293 cells were transfected in parallel with constructs coding for the wild-type IL6R sequence (Ser) or the mutated IL6R sequence (Pro) as indicated. The cells were stimulated with 30 or 100 ng/mL of recombinant human IL6. After stimulation, the proteins of the cell monolayers were collected and processed for Western blotting with an antibody against phosphorylated STAT3 (P-STAT3). Subsequently, the same blot was reacted with an antibody against GAPDH as a loading control. Both the wild-type and the mutated receptor are capable of downstream signaling.
Discussion
Today it is well accepted that the etiology of allergies involves both environmental and genetic factors. The allergic condition, where genetic factors contribute the largest part with up to 95%, is asthma (Ober and Yao, 2011).
In this study, we have investigated a family with two asthma patients for the presence of potentially disease-causing polymorphisms and mutations. To this end, we sequenced all exons of the index patient and compared the sequences with a list of variants previously reported in patients with allergies. We found 10 variants in susceptibility genes for allergic diseases, but only 4 of these variants co-segregated in the second patient of the same family, namely the variants in TNS1, NPSR1, RAD50, and IL6R.
Tensin-1 is a cytoskeletal protein involved in the formation of cell-substrate junctions (Lo et al., 1994; Chen et al., 2002). A sequence variant in this protein might, therefore, be involved in the dysfunction of the barrier at the surface of the lung or skin where an allergen might breach through the epithelial barrier. A significant association of tensin-1 with lung function and respiratory disease has been described in a meta-analysis of the SpiroMeta Consortium (Repapi et al., 2010). The V1604I variant (MAF 0.346) has also been found in a GWAS for asthma and lower airway disease (Imboden et al., 2012). NPSR1 is a G-protein coupled receptor involved in calcium signaling (Zhang and Tao, 2019). It is expressed in the epithelia of bronchi and the gastrointestinal tract as well as in the brain. There are numerous reports in the literature about the involvement of the N107I variant of NPRS1 in the pathogenesis of asthma and other IgE-mediated diseases (Laitinen et al., 2004; Andiappan et al., 2013; Acevedo et al., 2017). One group (Vendelin et al., 2005) reported that the expression of NPSR1 with the N107I variant is about 10-fold higher in the bronchi of asthmatic patients when compared with normal controls. Another group showed that the N107I variant leads to an increase in the potency of the receptor (Bernier et al., 2006). With a MAF of 0.446, this variant occurs frequently in the western population, but both of our patients contained the variant in homozygous form (frequency 0.199). RAD50 is a DNA repair protein required to repair double-strand breaks (Syed and Tainer, 2018). The RAD50 gene has repeatedly emerged from searches for susceptibility genes associated with asthma and atopic dermatitis (Akhabir and Sanford, 2011; Raby, 2019). Interestingly, RAD50 was the only gene that could be replicated in a study of childhood asthma and 50 reported asthma risk genes (Murk et al., 2011). Finally, IL6R is the receptor for the important inflammatory cytokine IL6.
The IL6 signaling system is highly complex and involves IL6, IL6R, and gp130 (Wolf et al., 2014; Garbers et al., 2015; Schaper and Rose-John, 2015). IL6R occurs in two different forms, a membrane-bound form and a soluble form, which is generated by cleavage and shedding from the membrane-bound form. Both forms bind the ligand IL6 and present it to the transducer gp130, which represents the active signaling part of the system. Upon interaction with IL6R and its cytokine, Janus kinases (JAKs) are recruited to the intracellular tail of gp130 and downstream signaling is achieved by the JAK/STAT pathway. The gene for IL6R is specifically expressed in leukocytes (monocytes) and hepatocytes, whereas the gene for gp130 is ubiquitously expressed in most cells of the body. Interestingly, signaling through the soluble form of IL6R (called trans-signaling) accounts for the proinflammatory activities of IL6, whereas signaling through the membrane-bound form (called classical signaling) mediates the anti-inflammatory and regenerative activities of IL6 (Garbers et al., 2015; Schaper and Rose-John, 2015). Opposing effects are, therefore, provoked by IL6, depending on the specific way of signaling. It is the balance between trans-signaling and classical signaling that determines the final outcome in a particular tissue or an entire organism.
IL6R has been implicated in the etiology of many chronic diseases, including allergic asthma. A nonsynonymous variant of IL6R, D358A (rs8192284), has been described, which is associated with an increased susceptibility to asthma (Rafiq et al., 2007; Reich et al., 2007). This polymorphism occurs in the extracellular domain of the receptor close to the site, where IL6R is cleaved from the membrane, and leads to an increase in the levels of soluble IL6R (Ferreira et al., 2013). Although numerous other polymorphisms have been found in the gene for IL6R, the mutation S409P has never been described before. Only a variant (rs1277848589) at the adjacent position 1:154465199 has been found, which changes the TCT codon for serine into TTT for phenylalanine (S409F). This mutation is extremely rare (1/125568, MAF 0.000008) and is listed as deleterious. Since the data set of the Exome Aggregation Consortium (ExAC) includes >120,000 different exomes and since the S409P variant found in our study does not occur in this data set, its MAF must be <0.000008. Thus, the variant qualifies for a rare mutation that could—together with other variants—contribute to the onset of allergic asthma. Nevertheless, it is difficult to provide final proof that this variant is not simply a benign polymorphism. For instance, it will not be feasible to check a cohort of asthma patients for the occurrence of the S409P variant since this mutation is simply too rare. A logical approach would be to introduce the variant into the genome of a mouse and study its effect in an animal experiment. However, the IL6/IL6R system of the mouse differs substantially from that of humans and consequently, data from mice cannot directly be transferred to humans. For example, human IL6 binds to the mouse IL6R, but mouse IL6 does not bind to the human receptor (van Dam et al., 1993).
A clue to its role might be obtained from theoretical considerations using the prediction tool SIFT. The S409P mutation occurs in the intracellular domain of the receptor. Mutations at adjacent sites, such as position 406 (rs150127972 P406R), 411 (rs780683821 G411R), 413 (rs1214944137 L413R), and 415 (rs142732420 P415L), are all listed as deleterious, suggesting that this part of the protein is crucial for normal function. Another clue might come from an amino acid sequence alignment of IL6R from humans and other vertebrates (Fig. 4). Such an alignment demonstrates that serine at position 409 is conserved among different species, including human, dog, cow, horse, panda, and dolphin. However, a few rodents, such as rat, cavia, and chinchilla, contain a proline residue at this position as found in our variant. In this context, we should remember that the human and the mouse IL6/IL6R system differ substantially from each other (van Dam et al., 1993). Moreover, all rodents with a proline at the critical position also contain a nearby deletion of five residues (Fig. 4). This fact could mean—but does not prove—that the substitution of serine by proline might be tolerated in the three-dimensional (3D) structure of IL6R only if it occurs in combination with an adjacent deletion, which is not the case in our asthma patients. Knowledge of the 3D structure of IL6R could eventually help to solve this question but so far, only the 3D structure of the extracellular domain is available (Varghese et al., 2002).

Multiple sequence alignment of the C-terminal domain of IL6R from nine different vertebrates. Similar residues are boxed; the TM is indicated by a black frame. The position of the novel mutation is shown by an arrow. It occurs within an important motif (Y motif) used for sorting of IL6R to the basolateral side of polarized cells. TM, transmembrane domain.
In the IL6R/gp130 complex, IL6R appears to be involved in binding of the ligand, whereas gp130 is involved in transmitting the signal to the interior of the cell by the JAK/STAT pathway (Garbers et al., 2015). Interestingly, a tyrosine residue is found at position 408 next to the S409P variant in the sequence 408-YSLG-411. Short linear sequence motifs with a tyrosine residue are frequently found in the intracellular domain of transmembrane proteins, where they often play a role in the turnover and subcellular sorting of the proteins (Pandey, 2009). They are specifically recognized by protein complexes of the cellular sorting machinery such as AP-1 and targeted to specific membrane compartments. In fact, one research group showed that the YSLG motif of IL6R is responsible for sorting of the receptor to the basolateral side of polarized epithelial cells (Martens et al., 2000). When the YSLG motif was deleted, the receptor was sorted to the apical side of the cells. It is, therefore, likely that the S409P mutation, which occurs next to the crucial tyrosine-408, will affect the correct sorting of IL6R to the basolateral side. The mutation identified in our two patients might, therefore, disturb intracellular sorting of IL6R. This might not be of importance in nonpolarized cells such as monocytes, but it might have severe consequences in polarized cells such as hepatocytes.
We have performed a preliminary experiment and demonstrated that the S409P variant of IL6R is capable of transmitting the IL6 signal to the interior of HEK293 cells with an efficiency similar to the wild-type receptor. However, this experiment does not allow any conclusions about the actual outcome within an entire organism where the activities of IL6 are determined by the balance between trans-signaling (through soluble IL6R) and classical signaling (through the membrane bound receptor). Moreover, there is evidence that IL6R is found only on the basolateral plasma membrane of hepatocytes, where it faces the blood stream and where it is susceptible to circulating IL6 (Martens et al., 2000). The HEK293 cells used in our experiment are not polarized and will, therefore, not reveal the effects of disturbed receptor sorting on the extent of inflammation. Thus, further studies are needed to investigate the relevance of the S409P variant and its potential involvement in the predisposition to asthma and other allergic diseases.
As one of the reviewers pointed out, there is the possibility that the IL6R mutation of the family might also be involved in the onset of psoriasis. Both the index patient II:3 and his son III:3 show signs of psoriasis and both carry the S409P variant. Psoriasis is considered to be an autoimmune disease that commonly affects the skin of the elbows, knees, and scalp. Although the exact causes of psoriasis are not known, the development of the disease has a clear genetic basis. The index patient suffers from mild psoriasis vulgaris at elbows and knees, which is successively treated with Dermovate. His son III:3 suffers from scalp psoriasis or seborrheic psoriasis that is treated with ketoconazole (a final differentiation between scalp psoriasis and seborrheic psoriasis has not yet been performed because this would have required extended histology of skin biopsies). However, the daughter III:4 also suffers from scalp psoriasis or seborrheic psoriasis diagnosed by the same dermatologist. Although we cannot exclude the possibility that the S409P variant might be involved in the onset of psoriasis, a direct contribution seems unlikely since the daughter does not carry the S409P mutation in IL6R (Table 2).
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.