
Abstract
Objective:
Prostate cancer (PCa) is one of the leading causes of death by solid tumors in men, and autophagy plays an important role in tumor progression and treatment. This study was designed to construct and evaluate an autophagy-related prognosis model of PCa with long noncoding RNAs (lncRNAs).
Materials and Methods:
RNA sequencing data and clinical features were obtained from The Cancer Genome Atlas (TCGA) prostate adenocarcinoma (PRAD) dataset. These data were analyzed by Pearson correlation and Lasso regression to enrich for six autophagy-related prognostic lncRNAs that were used to build a progression-free interval (PFI) prognostic model. In addition, survival and receiver operating characteristic (ROC) curve analyses were performed to evaluate the PFI's prognostic effect. Moreover, we used a nomogram analysis combined with other clinical features to improve its accuracy. Finally, we verified the biological function of LINC00342 using cell-based assays.
Results:
Six differentially expressed autophagy-related lncRNAs (LINC00342, AC092171.4, PCAT7, AC048341.2, FGF14-AS2, and AC005180.2) were identified between PCa and adjacent noncancerous tissues from the TCGA-PRAD dataset. A novel risk score model was established based on these six autophagy-related lncRNAs. PCa patients were then stratified into high- or low-risk groups in terms of their PFI; and the area under the ROC curve (AUC) determined to be significant. Moreover, we developed a nomogram with other clinical features that could be used for clinical decision-making. Lastly, we verified the oncogenic role of LINC00342 in promoting autophagy.
Conclusions:
We developed a novel six autophagy-related lncRNA signature for prediction of PFI in PCa. These lncRNAs may help establish individualized biomarkers and therapeutic targets for patients with PCa. Moreover, we also found an oncogenic role for LINC00342 in autophagy in PCa.
Introduction
In developed countries, prostate cancer (PCa) is the most commonly detected solid tumor with high morbidity in men (Siegel et al., 2015, 2020). Surgery, endocrine therapy, and radiation therapy can have substantial therapeutic effects in most PCa cases (Teo et al., 2019). Prostate-specific antigen (PSA) test is still the most commonly used test to monitor PCa recurrence. However, PCa is a heterogeneous tumor, and the limited specificity and sensitivity of PSA fail to meet the individualized treatment in different patients. Therefore, there is an urgent need to identify novel prognostic methods.
The human genome comprises ∼98% noncoding RNAs (ncRNAs) and 2% protein-coding RNAs (Anastasiadou et al., 2018). Research on ncRNAs has provided new insights into cancer biology. MicroRNAs (miRNAs) and long noncoding RNAs (lncRNAs) are important members of ncRNAs. Moreover, their clinical value and biological functions in cancer have been widely reported. For example, Lin et al. (2017) found that miR-30d promotes tumor growth and angiogenesis through the MYPT1/c-JUN/VEGFA pathway and predicts aggressive prognosis in PCa. Another study revealed that miR-489-3p enhances proliferation and glycolytic potential by regulating SIX1 expression in melanoma (Yang et al., 2020).
In addition, lncRNAs also play important roles in tumorigenesis and progression. For instance, THAP9-AS1 promotes the progression of pancreatic cancer (PC) by targeting miR-484 and combining with YAP and is associated with the poor clinical outcomes (Li et al., 2020). Besides, the lncRNA like HOXB-AS3 can directly regulate ribosomal RNA transcription to drive the exacerbation in mutated NPM1 leukemia (Papaioannou et al., 2019).
Autophagy is a process of recycling proteins and intracellular components in cells and is triggered by survival stress. Autophagy plays a dual role in tumorigenesis and progression (Levy et al., 2017). However, autophagy usually promotes cancer progression in established tumors. Transcription factor EB, induced by transforming growth factor beta, facilitates progression in PC by activating autophagy and predicts unfavorable outcomes in patients (He et al., 2019). In addition, dCTP pyrophosphatase-1 promotes progression in PCa by inducing autophagy (Lu et al., 2018).
LncRNAs also play an important role in regulating autophagy. The lncRNA MALAT1 can sponge off miR-204, thereby activating autophagy to boost the proliferation of gastric cancer (Shao et al., 2020). Moreover, lncSNHG5 stimulates autophagy during the development of resistance to chemotherapy through the miR-32/DNAJB9 axis in acute myeloid leukemia (Wang et al., 2020a). Nevertheless, there are no reports of prognostic models based on autophagy-related lncRNAs in PCa. In this study, we screened the autophagy-related lncRNAs in The Cancer Genome Atlas (TCGA)-prostate adenocarcinoma (PRAD) cohort and developed a novel risk score model for assessing progression-free interval (PFI). In addition, we verified the biological function of the most abundant oncogenic lncRNA, LINC00342, in PCa using cell-based assays in vitro.
Materials and Methods
Ethics statement
This study was approved by the institutional Ethics Committee of The First Affiliated Hospital of Jinan University, China.
Data acquisition and processing
RNA sequencing (RNA-seq) data (level 3), including FPKM (fragments per kilobase of exon model per million mapped fragments) and counts, and corresponding clinical information for 551 samples with PCa were obtained from the TCGA using the UCSC Xena browser (https://xenabrowser.net/). The FPKM values of RNA-seq data for 499 PCa samples and 52 adjacent tissues were processed by normalizeBetweenArrays function from the limma package. GRCh38.p13 from the Ensembl human genome browser (www.ensembl.org/index.html) was used to retrieve the required expression information of the protein-coding genes (mRNAs) and lncRNAs from RNA-seq data. The collected clinicopathological data included patients' age at initial pathological diagnosis, stage, TNM classification, PFI status, and number of days of PFI. A total of 222 autophagy-related genes from the Human Autophagy Database (HADb) (www.autophagy.lu/) were obtained for subsequent analyses.
LncRNA differential expression and correlation analyses
The expression profiles of lncRNAs and autophagy genes were determined using RNA-seq. Pearson correlation was used to calculate the correlation between autophagy-related genes and lncRNAs. The DESeq2 package was used to determine the differentially expressed lncRNAs based on an absolute log2foldchange (log2FC) ≥1 and an adjusted p-value <0.05. Furthermore, Pearson correlation was used to calculate the correlation between the autophagy-related genes and differentially expressed lncRNAs, and the lncRNAs with the correlation coefficient |R2| > 0.5 and p < 0.05 were considered to be autophagy-related lncRNAs. The autophagy-related lncRNAs were considered as candidate genes for further analysis.
Construction and assessment of the autophagy-related gene-based prognostic signature
The least absolute shrinkage and selection operator (LASSO) analysis with L1 regularization was considered as a variable shrinkage and selection method for linear regression. The candidate autophagy-related lncRNAs were incorporated into LASSO regression with a 10-fold cross-validation to optimize the prognostic multi‑lncRNA signature and establish an autophagy-related gene-based prognostic signature using the plmnet R package. The risk score formula was established as follows:
Furthermore, the risk score for each patient was calculated by combining the expression values of prognostic autophagy-related lncRNAs weighted by their coefficients, and the patients were subdivided into high-risk and low-risk groups based on the median risk score. The Kaplan-Meier (KM) method with log-rank test was performed to assess the difference in PFI between high- and low-risk groups using survminer and survival packages. The time-dependent receiver operating characteristic (ROC) curves of PFI and area under the ROC curve (AUC) were used to evaluate the performance of the signature in 1, 3, and 5 years using the survivalROC package.
Analysis of the autophagy-related lncRNA signature and clinical characteristics
The ROC curve was also used to examine the relationship between the prediction accuracy of the prognostic model and patients' clinical characteristics, including age, Gleason score, and so on. Univariate and multivariate Cox regression analyses were performed to describe six autophagy-related lncRNAs affecting PFI in patients, which were presented in the form of hazard ratios and corresponding 95% confidence intervals. A nomogram using the rms package and concordance index (C-index) with 1000 bootstrap resampling was utilized to evaluate the accuracy of the prognostic signature. A higher C-index (0.5-1.0) demonstrates greater discrimination ability of the signature.
Cell lines, cell culture, and treatment
The benign prostate hyperplasia cell line (BPH-1) and PCa cell lines (C4-2, LNCaP, PC-3, DU145, and 22Rv1) were purchased from the ATCC (Manassas, VA). All cell lines were cultured in RPMI-1640 medium (Gibco, Thermo Fisher Scientific) with 10% fetal bovine serum (Gibco, Thermo Fisher Scientific) and 1% double antibiotics (streptomycin and penicillin; Gibco, Thermo Fisher Scientific) at 37°C and 5% CO2. To detect the autophagy-related markers, PC-3 and 22Rv1 cells were treated with Earle's Balanced Salt Solution (EBSS; Gibco, Thermo Fisher Scientific) with or without chloroquine (CQ; Sigma) for 8 h before the next experiment.
RNA interference and cell transfection
Small interfering RNAs (siRNAs) targeting LINC00342 were obtained from Hipobio Company (Zhejiang, China), and Lipofectamine 3000 (Invitrogen) reagent was used to transfect these siRNAs into PC-3 and 22Rv1 cells. The siRNA sequences targeting LINC00342 are shown in Supplementary Table S1.
Cell viability and cell colony formation assay
The proliferation and colony formation abilities of PC-3 and 22Rv1 cells transfected with siRNAs were examined using Cell Counting Kit-8 (CCK-8) reagents (Meilunbio, China) and colony formation assays, respectively. Detailed procedures are available in our previous report (Zhou et al., 2019). Each experiment was repeated three independent times.
RNA extraction, reverse transcription, and quantitative real-time polymerase chain reaction
Total RNA from PC-3 and 22Rv1 cells was extracted using TRIzol reagent (Invitrogen), and the concentration and quality of each RNA sample were tested using NanoDrop (Thermo Fisher Scientific). Total RNA was reverse-transcribed to cDNA using the PrimeScript™ RT Reagent Kit with gDNA Eraser (RR0471, Takara). The SYBR Green Real-time PCR Master Mix (QPK-201, TOYOBO) was used to detect gene expression using the CFX Connect qPCR Detection System (Bio-Rad). Relative gene expression levels were calculated using the 2−ΔΔct method. ACTB was used as an internal reference for normalization. Differences between groups were analyzed using the unpaired two-tailed Student's t test, and the data were showed as mean ± standard deviation. The polymerase chain reaction (PCR) primers for β-actin and LINC00342 are listed in Supplementary Table S1.
Protein extraction and western blotting
PC-3 and 22Rv1 cells were lysed using radioimmunoprecipitation assay (RIPA) buffer with 1% phenylmethylsulfonyl fluoride (PMSF) on ice for 20 min, followed by detection of each sample concentration using bicinchoninic acid (BCA) protein assay. Primary antibodies against GAPDH (Proteintech), p62 (Santa Cruz), and MAP1LC3A (LC3, Novus) were used at a dilution of 1:1000. Detailed western blotting procedures were performed as described in our previous report (Zhou et al., 2019).
Statistical analyses
R language (version 4.0.2) was used to perform all the statistical analyses in the study. The heat maps were constructed using the pheatmap package (version 1.0.12). Differences were considered statistically significant at p < 0.05.
Results
Differentially expressed autophagy-related lncRNAs in PCa
The flowchart of this study is shown in Figure 1. We removed three replicated PCa sequencing data and obtained a final RNA-seq data containing 496 PCa tissues and 52 pericancerous tissues from the UCSC. A total of 14,057 lncRNAs were annotated and retrieved by GRCh38.p13 from the Ensembl human genome browser. As shown in Figure 2A, differentially expressed analysis identified 718 upregulated and 1212 downregulated lncRNAs with |logFC| ≥1 and adjusted p < 0.05, respectively (Fig. 2A). Furthermore, Pearson correlation analysis between the differentially expressed lncRNAs and autophagy-related genes was conducted to select 89 autophagy-related lncRNAs with |R| > 0.5 and p < 0.05.
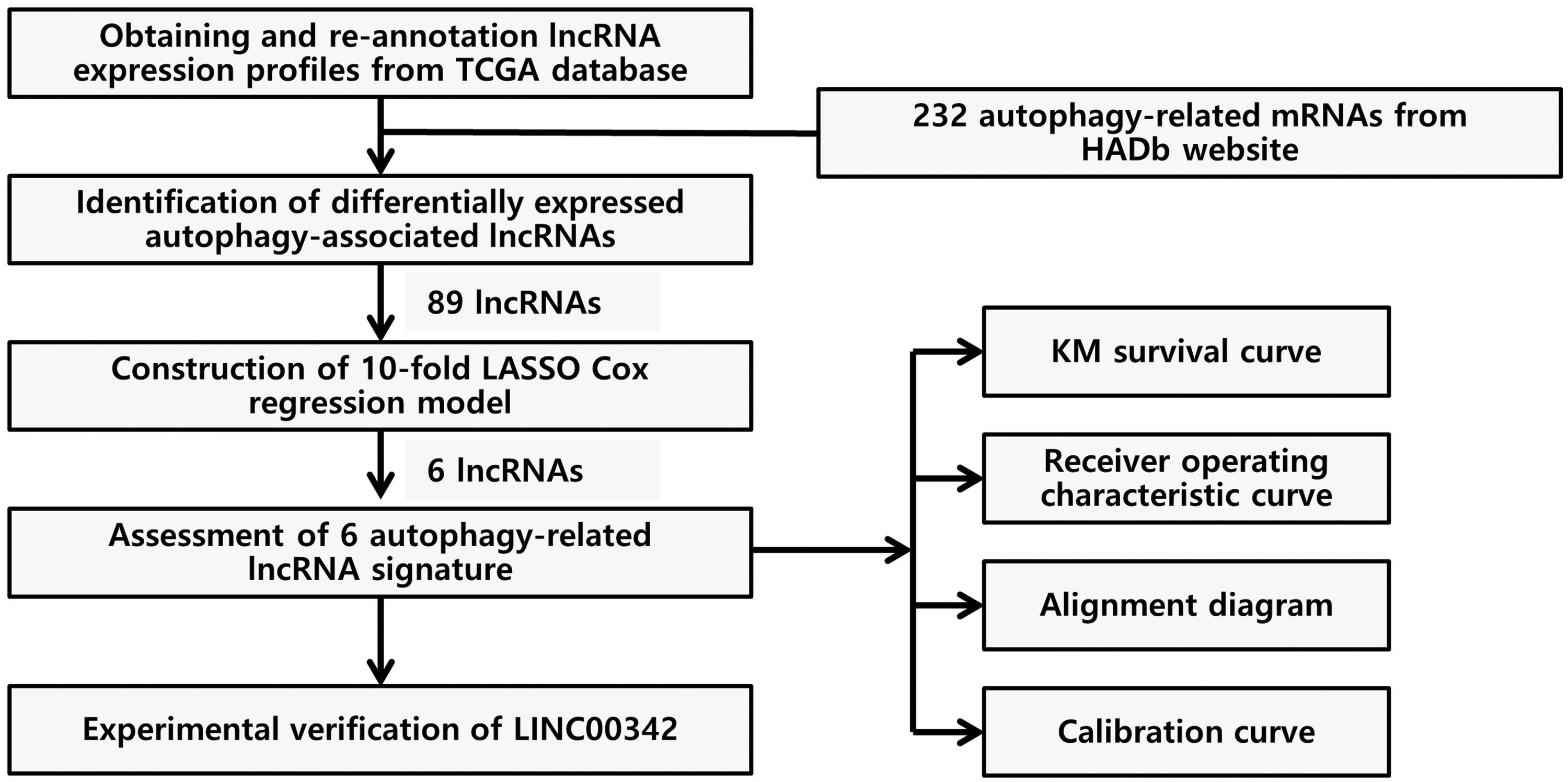
The study flowchart. HADb, human autophagy database; LASSO, least absolute shrinkage and selection operator; lncRNA, long noncoding RNA; KM, Kaplan-Meier; TCGA-PRAD, The Cancer Genome Atlas-prostate adenocarcinoma.
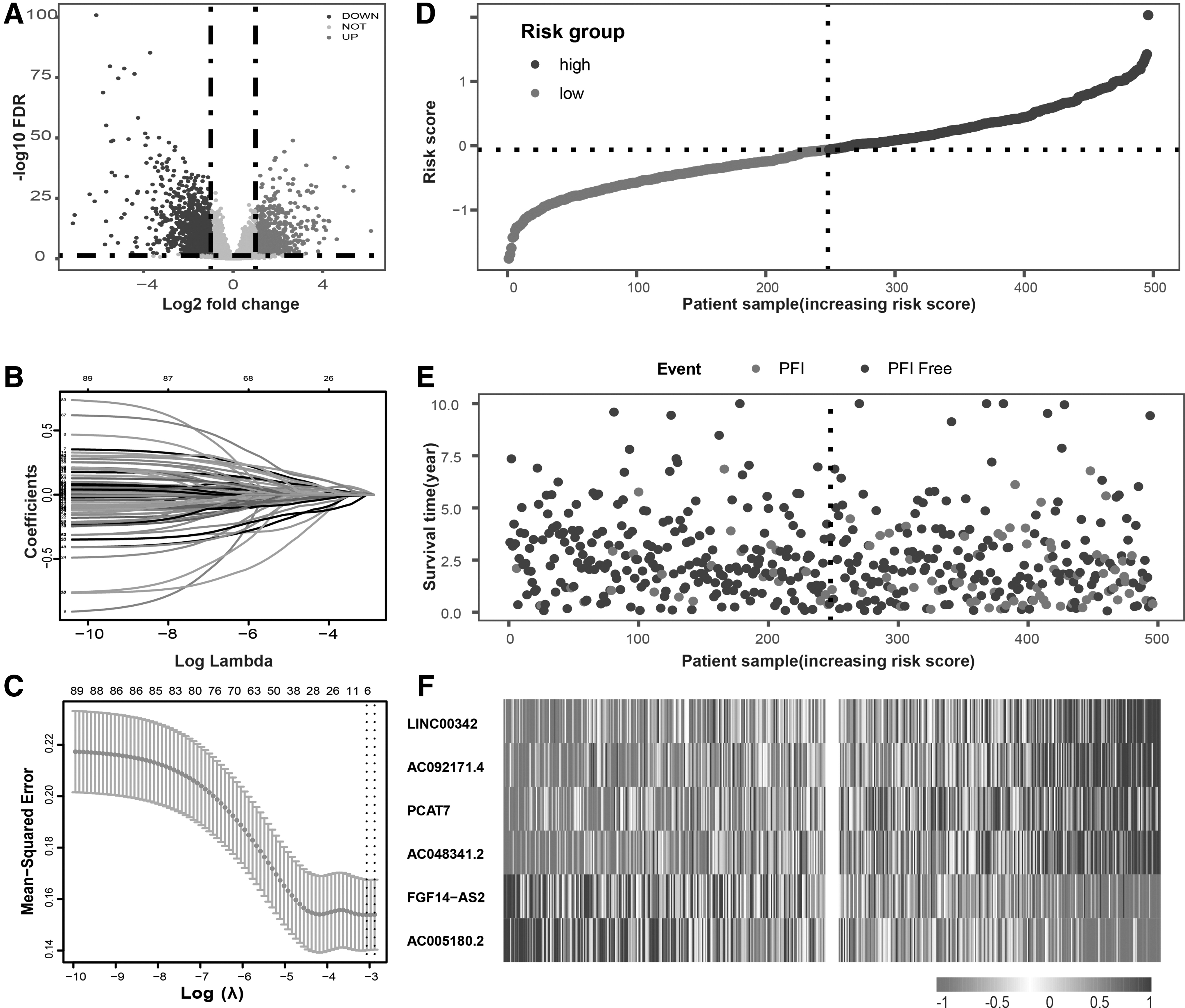
Differentially expressed autophagy-related lncRNAs and construction of prognosis model in patients with PCa.
Identifying a prognostic autophagy-related lncRNA signature
LASSO regression was applied to construct a prognostic signature to select lncRNAs with prognostic values from 89 autophagy-related lncRNAs. Six autophagy-related lncRNAs and coefficients were identified after 10-fold cross-validation LASSO regression with lambda 0.0465517, including LINC00342, AC092171.4, PCAT7, AC048341.2, FGF14-AS2, and AC005180.2 (Fig. 2B, C). The expression levels of the six autophagy-related lncRNAs and their coefficients were integrated to establish a signature formula. The formula was as follows: Risk score = 0.004055*LINC00342 + 0.002756*AC092171.4 + 0.004313*PCAT7 + 0.001299*AC048341.2 + (−0.00045)*FGF14-AS2 + (−0.000558)*AC005180.2. Based on the signature, the risk scores were calculated, and 496 PCa patients with PFI were subdivided into the high- and low-risk groups according to the median value (0.2087). The patients were sorted by the risk score (Fig. 2D), and the survival status and heatmap for the expression levels of the six autophagy-related lncRNAs are shown in Figure 2D-F. Patients with a higher risk score had a shorter PFI than those with low-risk score. In addition, the heatmap demonstrated noticeable differences in the high- and low-risk PCa patients. Higher expression levels of risk factors (FGF14-AS2 and AC005180.2) were observed in high-risk patients, while higher levels of protective factors (LINC00342, AC092171.4, PCAT7, and AC048341.2) were observed in low-risk patients. KM curves based on different risk score groups are shown in Supplementary Figure S1. Besides, the correlation between risk score model associated lncRNAs and clinical features is displayed in Supplementary Table S2.
Assessment of the autophagy-related lncRNA signature
Next, the KM survival curve demonstrated that the PFI status of the high-risk group was significantly lower compared with low-risk group (p < 0.0001, Fig. 3A). Based on the risk scores of the six-lncRNA signature, the AUCs corresponding to 1, 3, and 5 years of survival were 0.681, 0.66, and 0.683, respectively (Fig. 3B). Furthermore, a nomogram was constructed to estimate the 1-, 3-, and 5-year survival probabilities. Nomograms are generally used to provide the clinician with a quantitative method to predict a patient's probability of survival based on the points assigned for each prognostic factor. In this study, the prognostic factors included risk score, age, Gleason score, and T stage (Fig. 3C). The concordance index (C-index) value for the nomogram was 0.748. In addition, the calibration curve analysis for 3- and 5-year survival showed nomogram-predicted recurrence along the X-axis, observed outcome on the Y-axis, and best prediction represented along the cater-cornered (Fig. 3D).
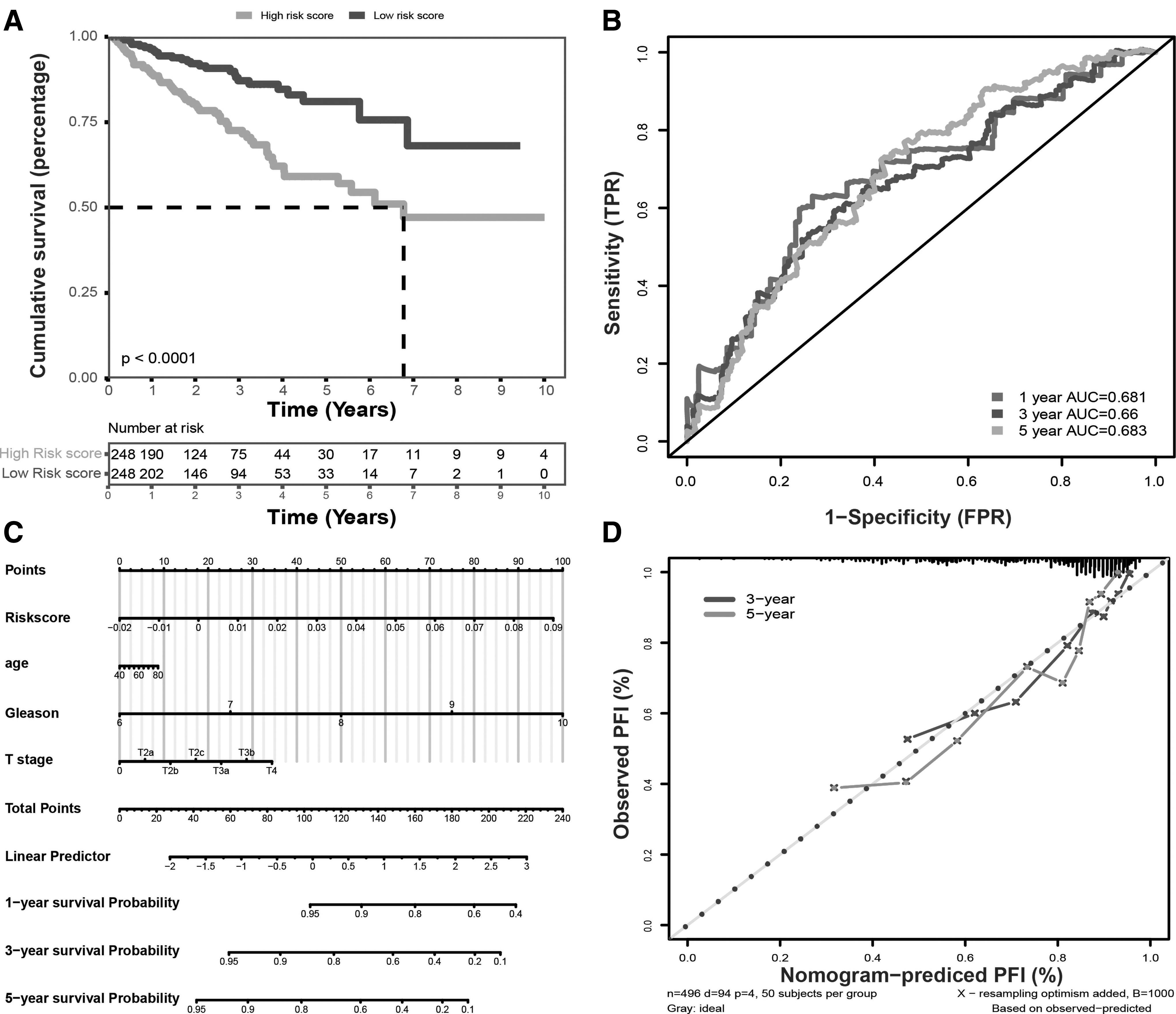
Assessment of the autophagy-related lncRNA model.
Knockdown of LINC00342 suppressed proliferation and autophagy in PCa cells
By analyzing the position of LINC00342 in the human genome from the NCBI (https://www.ncbi.nlm.nih.gov/) and LNCipedia (https://lncipedia.org/) databases, we identified that it was located on chromosome 2q11.1 and transcribed from the antisense stand comprising three exons and two intergenic introns (Fig. 4A). Moreover, LINC00342 was significantly upregulated in the TCGA-PRAD dataset (Fig. 4B). We then examined the basal expression of LINC00342 in benign prostate and PCa cell lines. LINC00342 was highly expressed in PCa cell lines, especially in PC-3 and 22Rv1 cells than in benign prostate cells (Fig. 4C; LNCaP: 53.65 ± 0.6696, p < 0.0001; C4-2:13.27 ± 0.5256, p < 0.0001; PC-3: 252.2 ± 15.88, p < 0.0001; DU145: 213.7 ± 6.654, p < 0.0001; 22RV1, 229.9 ± 17.59, p = 0.0002). Therefore, we designed three siRNAs specifically targeting LINC00342 and transfected PC-3 and 22Rv1 cells with negative control (NC) oligo RNAs and these three specific siRNAs. Quantitative real-time PCR (qPCR) assay showed that transfection with siRNA-1 and siRNA-2 significantly knocked down the expression of LINC00342 by 60%-70% than that with NC siRNA (Fig. 4D; 22Rv1-si-1: 0.44 ± 0.062, p = 0.0032; 22Rv1-si-2: 0.41 ± 0.052, p = 0.0019; 22Rv1-si-3: 0.48 ± 0.023, p = 0.0015; PC-3-si-1: 0.36 ± 0.050, p = 0.0015; PC-3-si-2: 0.30 ± 0.078, p = 0.0022; PC-3-si-3: 0.75 ± 0.064, n = 0.0529). Thus, we used these two siRNAs for subsequent experiments. Next, in the CCK-8 assay, we found that silencing LINC00342 by siRNAs remarkably inhibited the proliferative ability of PC-3 and 22Rv1 cells than that with NC siRNA (Fig. 4E). Moreover, decreased expression of LINC00342 significantly suppressed the cell colony formation ability in both PC-3 and 22Rv1 cells than that with basal expression (Fig. 4F). As we previously confirmed that LINC00342 is closely related to autophagy, we examined the variable expression of autophagic markers, including LC3 and p62 after knocking down LINC00342 in PC-3 and 22Rv1 cells. Western blotting analysis confirmed that downregulation of LINC00342 significantly reduced the conversion of LC3-II/LC3-I and blocked the degradation of p62 in PC-3 and 22Rv1 cells cultured in EBSS with or without CQ for 8 h than that in NC group (Fig. 4G).

Downregulation of LINC00342 attenuates proliferation and autophagy in PCa cells.
Discussion
In men, PCa is the most common and progressive tumor that requires an effective prognostic indicator for further treatment (Siegel et al., 2015, 2020; Porzycki and Ciszkowicz, 2020). The end of PFI is characterized by a new disease event, which could be a primary tumor, local recurrence, or distant metastasis (Liu et al., 2018). In the TCGA-PRAD dataset, PFI is the most reliable clinical end point. Due to the heterogeneous mechanism of PCa progression, existing methods for prognosis prediction are insufficient. There are increasing reports that autophagy plays an important role in PCa progression and therapeutic resistance (Xu et al., 2020). For example, hypoxia regulator HIF‑1α can strengthen cell mobility and chemosensitivity by regulating hypoxia‑induced autophagy through the miR‑224‑3p/ATG5 axis in glioblastoma and astrocytoma (Huang et al., 2019). Li et al. (2019) showed that high expression of HERC3 accelerated degradation of SMAD7 in an autophagy-dependent manner, inducing epithelial-mesenchymal transition and chemoresistance in glioblastoma.
Molecular signatures of DNA, mRNA, and noncoding RNA have been used to predict the prognosis in many tumors (Li et al., 2017; Shen et al., 2018; Wang et al., 2020). For example, Li et al. reported that 5 hmC DNA modification biomarkers in circulating cell-free DNA (cfDNA) are better than the markers from tumor biopsies in colorectal and gastric cancers. In addition, Shen et al. showed that a new classifier combining miRNA expression with Ki-67 levels will have a better prognostic accuracy in cutaneous T cell lymphoma. Moreover, lncRNA is a key member of ncRNAs, and its prognostic value has been widely reported. However, the role of autophagy-related lncRNAs in predicting the PFI in PCa remains unclear.
In this study, we identified prognostic autophagy-related lncRNAs to establish a reliable model for predicting the PFI in patients with PCa. First, we identified differentially expressed lncRNAs between PCa and cancer adjacent tissues in the TCGA-PRAD dataset. Second, we analyzed their prognostic value and Pearson correlation with autophagy-related mRNAs and identified 89 prognostic autophagy-related lncRNAs.
Furthermore, LASSO regression was applied to construct a prognostic signature based on these 89 prognostic autophagy-related lncRNAs. Of these, six autophagy-related lncRNAs were included to construct a risk score model. This model assigns a shorter PFI for the higher risk score group. To improve the accuracy of the model, we used a nomogram analysis combined with other clinical features. Nomograms are a common tool used in daily clinical practice. Using our data, we created a nomogram containing risk score, age, Gleason score, and T stage as variables. The calibration diagram confirms the reliable prediction of our nomogram. This visual scoring system may provide the physicians a choice to design a more personalized treatment plan for the patients with PCa.
LINC00342, AC092171.4, PCAT7, AC048341.2, FGF14-AS2, and AC005180.2 were identified in our risk score model. Some of these are reported in different tumors. For instance, Sun et al. (2020) found that AC092171.4 can promote progression in hepatocellular carcinoma through the microRNA-1271/GRB2 axis. Various studies have verified that PCAT7 promotes the progression of numerous types of cancers, such as nasopharyngeal carcinoma, nonsmall cell lung cancer, and PCa (Liu et al., 2017a, 2017b; Lang et al., 2020). In contrast, the lncRNA FGF14-AS2 functions as a suppressor in many cancers, including colorectal cancer and breast cancer (Yang et al., 2019; Hou et al., 2020; Jin et al., 2020). LINC00342 has been widely studied in different types of cancer. It promotes progression and predicts poor outcomes in nonsmall cell lung cancer (Chen et al., 2019; Tang et al., 2019). In addition, LINC00342 also acts as a tumorigenic factor in colon cancer and infantile hemangioma through the competing endogenous (ceRNA) regulatory network (Liu et al., 2019; Miao et al., 2020).
We selected LINC00342 that is upregulated in our data for further experimental validation to verify its functions in PCa as its high expression level is positively correlated with poor prognosis. LINC00342 is an intergenic lncRNA with increased expression levels in PCa tissues and cell lines than adjacent tissues and benign hyperplastic cells. Knockdown of LINC00342 reduced cell viability and autophagy levels in PC-3 and 22RV1 cells than that in NC group. These data suggested that the oncogenic effect of LINC00342 may relate to autophagy. Moreover, the mechanism of action of LINC00342 in autophagy adjustment is unknown. Most of the lncRNAs function as miRNA sponge. In addition, lncRNAs can directly interact with proteins. According to the LNCipedia and published articles, the most likely role of LINC00342 in PCa is through the ceRNAs and autophagy-related protein interaction; however, this is a preliminary evidence. We plan to conduct further in vitro and in vivo studies to identify the mechanism of action of LINC00342 in PCa.
Conclusion
In summary, we developed a novel autophagy-related lncRNA risk score model to predict the PFI in PCa that may help in establishing individualized treatment. Moreover, we preliminarily tested the oncogenic effects of LINC00342 in PCa. However, a large-scale, multicenter cohort study is needed to validate this autophagy-lncRNA prognostic model in PCa.
Ethics Approval
The experimental procedures in this study were approved by the Ethics Committee of The First Affiliated Hospital of Jinan University, China.
Availability of Data and Materials
The datasets used and/or analyzed in this study are available from the corresponding author upon reasonable request.
Footnotes
Authors' Contributions
The author(s) read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Leading Specialist Construction Project Department of Urology, The First Affiliated Hospital, Jinan University (No. 711006), the Science and Technology Planning Project of Guangdong (No. 2020A1414010348), and the Science and Technology Project of Zhongshan City (No. 2019B1063, 2020B1073).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.