
Abstract
Objective:
Diabetic nephropathy (DN), the most severe complication of diabetes mellitus, is characterized by albuminuria and progressive loss of kidney function. Dapagliflozin (DAP), a sodium-glucose cotransporter inhibitor, is an oral medication that improves blood glucose control in diabetic patients. However, the effects and mechanisms of DAP on DN remain unclear.
Materials and Methods:
The effect of DAP was based on a retrospective cohort study of patients who underwent 2-year surveillance, and the concentration of urine albumin-to-creatinine ratio, glomerular filtration rate, and serum creatinine were collected after treatment with DAP. To investigate the underlying mechanisms through which DAP reduces urinary albumin excretion, we used RNA-sequencing (RNA-seq) to analyze gene expression in human kidney 2 (HK-2) cells treated with DAP.
Results:
The retrospective cohort analysis indicated that DAP could reduce the excretion rate of urinary albumin in patients with type 2 diabetes and renal impairment. The results of the RNA-seq experiments showed 349 differentially expressed genes between DAP-treated HK-2 cells and control cells. Gene ontology annotation enrichment analysis showed that DAP mainly affected the expression of integral component of membrane- and cell junction-related genes, while the Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis showed that DAP primarily downregulated the expression of gene clusters associated with cyclic adenosine monophosphate, mitogen-activated protein kinase, and cyclic guanosine monophosphate-protein kinase G signaling pathways, which play critical roles in the progression of DN.
Conclusion:
Our results shed light on the mechanism by which DAP controls DN progression and provide a theoretical basis for the clinical treatment of DN.
Introduction
Albuminuria, declining glomerular filtration rate (GFR), and arterial hypertension are symptoms of diabetic nephropathy (DN), which affects 20-40% of patients with type 1 and type 2 diabetes (Muskiet et al., 2019). DN clinically manifests as macrovascular and microvascular complications, which are the most common causes of end-stage renal disease (ESRD) (Flyvbjerg, 2017). The molecular mechanisms underlying DN remain unclear. However, it is well established that hemodynamic and metabolic pathway changes in diacylglycerol (DAG)-protein kinase C (PKC), advanced glycation end products, polyol, and hexosamine pathways, are involved in the pathogenesis of DN (Liu et al., 2015; Shimizu et al., 2017; Chen and Fang, 2018). In addition, numerous studies verified that cyclic adenosine monophosphate (cAMP), mitogen-activated protein kinase (MAPK), and cyclic guanosine monophosphate-protein kinase G (cGMP-PKG) signaling pathways play critical roles in the development of DN (Mora and Navarro, 2004; Inada et al., 2008; Cao and Cooper, 2011).
Currently, controlling hyperglycemia, hypertension, and dyslipidemia to stabilize the disease and minimize complications is the main strategy for DN treatment (Fineberg et al., 2013). However, management of these risk factors is not sufficient to completely prevent DN progression (Gilbert, 2019). Dapagliflozin (DAP) is an inhibitor of sodium-glucose cotransporter-2 (SGLT-2), which is responsible for the reabsorption of most of the glucose filtered by the kidney. In recent clinical trials, DAP was initially shown to control blood glucose levels in patients with diabetes and could significantly reduce the risk of DN and cardiovascular or renal failure in patients with chronic kidney disease (Heerspink et al., 2020). In a phase 3 clinical trial, DAP treatment was associated with a significant improvement of renal function (defined as a sustained decrease in glomerular filtration rate of >50%) and a 39% reduction in the combined end-point risk of ESRD, renal death, and cardiovascular death in diabetic patients. The risk of all-cause death in patients with chronic kidney disease treated with DAP was also reduced by 31% compared with placebo treatment (Mathieu et al., 2018). However, the protective mechanisms of DAP on DN and other functions of DAP in renal tubular epithelial cells are still unclear.
RNA sequencing (RNA-seq) is the latest transcriptome analysis method developed using a new generation of sequencing technology (Wang et al., 2009). To investigate the mechanisms of DAP in the treatment of DN, we used the RNA-seq method to explore the gene expression profile and cellular function after DAP treatment in renal tubular epithelial cells. We found a total of 349 differentially expressed genes (DEGs) between DAP-treated human kidney 2 (HK-2) cells and control cells. Gene ontology (GO) annotation enrichment analysis showed that DAP mainly affected the expression of genes related to integral component of membrane and cell junction, whereas Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis showed that DAP affected the expression of gene clusters associated with cAMP, MAPK, and cGMP-PKG signaling pathways. We also confirmed that DAP could reduce the urinary albumin excretion rate in patients with type 2 diabetes with renal impairment. Our results provide new insights into the mechanism of DAP in reducing urinary albumin excretion in patients with DN.
Materials and Methods
Study design and data collection
The retrospective cohort study included patients who underwent 2-year surveillance at Sir Run Run Shaw Hospital, School of Medicine, Zhejiang University, from October 2017 to November 2019. The inclusion criteria for the cohort were as follows: patients with confirmed type 2 diabetes mellitus diagnosed according to the World Health Organization (WHO) (Mustaly-Kalimi et al., 2018) and Chinese official guidelines. A total of 47 subjects who had been diagnosed with DN and received DAP treatment were included. Subjects with urinary protein elevation caused by urinary tract infection or fever were excluded. The patients were followed up at least once and at most three times, ranging from 2 to 60 weeks. The glucose-lowering regimen was adjusted according to the experience of the doctors. Some patients were administered only DAP during the course of their treatment, and some switched from sulfonylurea drugs to DAP. The initial dose of DAP was 5 or 10 mg/day.
We collected data on the following variables from the hospital electronic medical record systems: demographic characteristics (age, gender, and body mass index, BMI), diabetes parameters (blood glucose concentration, mean duration of diabetes mellitus, and insulin resistance), and changes in physiological variables including the urine albumin-to-creatinine ratio (UACR), estimated GFR (eGFR), serum creatinine (SCr), fasting blood glucose, and glycosylated hemoglobin.
Cells and reagents
The human renal tubular epithelial cell line (HK-2; ATCC No. CRL-2190) was purchased from American Type Culture Collection (ATCC, Manassas, VA) and cultured in DMEM high-glucose (25 mM) medium supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL) in a 5% CO2 incubator at 37°C. DAP (BMS-512148) was purchased from MedCheExpress (Shanghai, China). DAP was dissolved in dimethyl sulfoxide (DMSO) for storage at 10 mM.
Cell treatment
HK-2 cells were seeded onto a 12-well plate at a density of 2 × 105/mL and incubated overnight. To avoid the potential interference of albumin present in FBS, cells were maintained in a serum-free high-glucose medium for 24 h and treated with or without 10 μM DAP for 24 h according to the previous studies (Han et al., 2018; Huang et al., 2019; Xu et al., 2021). The control group received an equal percentage of DMSO. Three replicate wells were included in each group.
RNA extraction, library construction, and sequencing
Total RNA was isolated from HK-2 cells treated with DMSO or DAP using TRIzol reagent according to the manufacturer's protocol (Invitrogen Life Technologies, Shanghai, China). The quality and concentration of the RNAs were determined using a nanodrop spectrophotometer (Thermo Scientific Technology, Shanghai, China). RNA integrity was evaluated using a Bioanalyzer 2100 (Agilent, CA) with RIN number >7.0, and confirmed by electrophoresis on a denaturing agarose gel. Poly (A) RNA was purified from 1 μg total RNA using Dynabeads Oligo (dT)25-61005 (Thermo Fisher Technology, CA) using two rounds of purification, and then fragmented into small pieces using a magnesium RNA fragmentation module (NEB) at 94°C for 7 min. The cleaved RNA fragments were reverse transcribed to cDNA using SuperScript™ II Reverse Transcriptase (Invitrogen Life Technologies, cat. no. 1896649), which were then used to synthesize U-labeled second-stranded DNAs with Escherichia coli DNA polymerase I (NEB), RNase H (NEB), and dUTP solution (Thermo Fisher Technology). An A-base was then added to the blunt ends of each strand, preparing them for ligation to the indexed adapters. Each adapter contains a T-base overhang to ligate the adapter to the A-tailed fragmented DNA. Single- or dual-index adapters were ligated to the fragments, and size selection was performed using AMPureXP beads. After heat-labile uracil-DNA glycosylase (UDG) enzyme (NEB) treatment of the U-labeled second-stranded DNAs, the ligated products were amplified by polymerase chain reaction (PCR) under the following conditions: initial denaturation at 95°C for 3 min; eight cycles of denaturation at 98°C for 15 s, annealing at 60°C for 15 s, and extension at 72°C for 30 s; and a final extension at 72°C for 5 min. The average insert size of the final cDNA library was 300 ± 50 bp. Finally, we performed 2 × 150 bp paired-end sequencing (PE150) on an Illumina Novaseq 6000 (LC-Bio Technology, Hangzhou, China) following the vendor's recommended protocol.
Functional and pathway enrichment analysis of DEGs
The DEGs were selected based on fold change >2 or fold change <0.5 and p-value <0.05, using R package edgeR (https://bioconductor.org/packages/release/bioc/html/edgeR.html) or DESeq2 (www.bioconductor.org/packages/release/bioc/html/DESeq2.html). Functional and pathway analyses of DEGs were performed using the Database for Annotation, Visualization, and Integrated Discovery. GO enrichment analysis of DEGs was performed using a previously described equation (Wang et al., 2018). An online database (www. genome. jp/kegg) was used to annotate and visualize the functions of DEGs and the signaling pathways involved (KEGG pathway enrichment). Gene expression and pathway analyses were performed by LC-Bio Technology.
Real-time quantitative PCR analysis
Total mRNA was extracted from DAP-treated and control HK-2 cells, and 1 μg of total RNA per sample was used for cDNA synthesis. Glyceraldehyde phosphate dehydrogenase was selected for normalization of genes of interest using the 2−ΔΔct method. Candidate genes for real-time quantitative PCR (RT-qPCR) are shown in Supplementary Table S1.
Statistical analysis
Distributions of categorical variables are presented as numbers and percentages and compared using the chi-squared test. Continuous variables are expressed as mean ± SEM and were compared using the independent-samples t-test. Binary multivariable logistic regression was performed to evaluate the potential association between the variables, and odds ratios (ORs) and 95% confidence intervals (CIs) were calculated at the same time. A one- or two-way ANOVA was used to determine the significance of differences between groups in RT-PCR analysis. Statistical analysis was conducted using SPSS version 19.0. Differences were considered statistically significant at p < 0.05.
Results
The UACR was decreased in diabetic patients after DAP treatment
In this study, we enrolled 47 patients, including 36 males and 11 females, aged 23 to 77 years, with an average of 48.21 ± 13.63 years. The average duration of diabetes was 8.62 ± 5.44 years. The median time of the first follow-up was 2 (0.5, 8) months. The patients were treated with 5 or 10 mg/day DAP to control their blood glucose levels. To confirm the effect of DAP on renal function in diabetic patients, we measured the levels of UACR, eGFR, and SCr. As shown in Figure 1A, the UACR decreased in 41 (87.2%) patients at the first follow-up after treatment with DAP. The level of UACR was decreased in 26 (96.3%) patients with 30-300 mg/kg UACR levels after DAP treatment. Among them, 11 (40.7%) patients had normal UACR levels after treatment with DAP, whereas 75.0% of patients with UACR levels >300 mg/kg had an average reduction of 5.3% (p > 0.05) in UACR levels after treatment. The levels of UACR continuously decreased in patients after DAP treatment (Fig. 1B). However, the levels of eGFR and SCr were not significantly affected by DAP administration (Fig. 1B). Our results demonstrated that DAP treatment significantly reduced UACR.
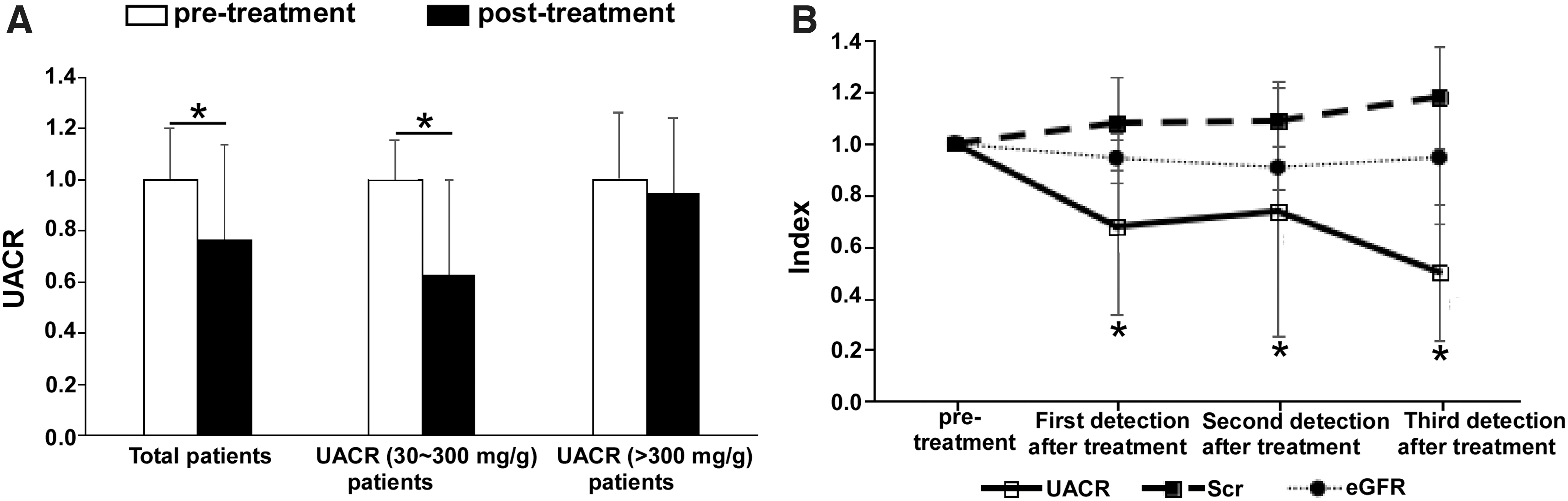
The UACR levels in diabetic patients were decreased after dapagliflozin treatment.
DAP treatment affects gene expression profiles of HK-2 cells
To investigate the underlying mechanisms of DAP in reducing the levels of UACR in diabetic patients, we examined gene expression in HK-2 cells after treatment with DAP using RNA-seq technology. The quality control of the RNA-seq data is shown in Supplementary Table S2. A total of 349 genes showed significant differential expression after DAP treatment (Fig. 2A, B). Among these genes, 280 genes were downregulated and 69 genes were upregulated (Fig. 2A, B). A hierarchical cluster of DEGs is partially shown in a heat map showing the top 100 DEGs (Fig. 2C). Surprisingly, a significant number of inflammatory genes were downregulated in the DAP treatment group, especially the protein activator of interferon-induced protein kinase (PPKAR2A-AS1), fibroblast growth factor receptor like 1 (FGFRL1), and G protein-coupled receptor associated sorting protein 1 (GPRASP1). This indicated that DAP might inhibit the inflammatory response of HK-2 cells by downregulating the expression of PPKAR2A-AS1, FGFRL1, and GPRASP1.
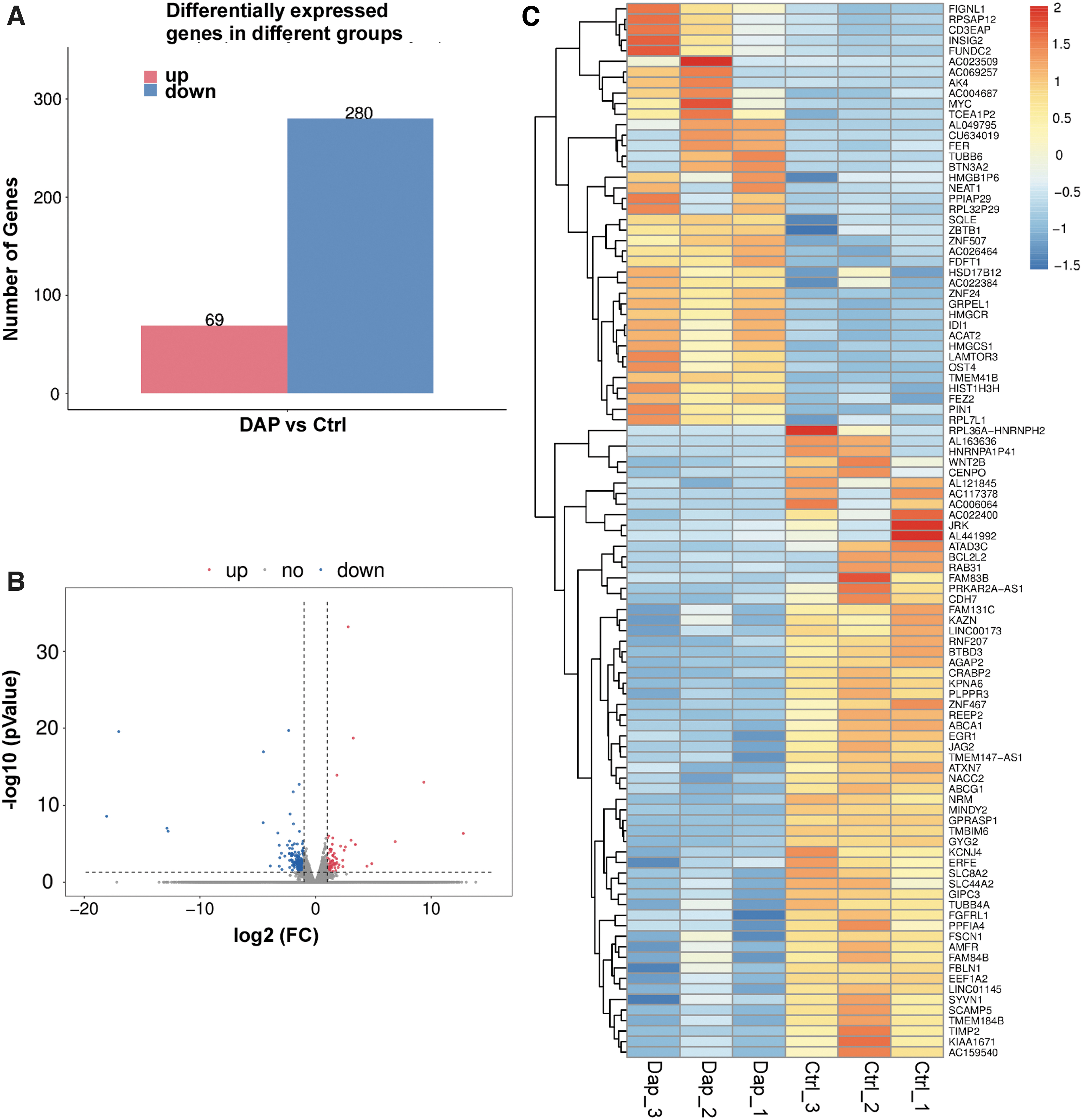
Dapagliflozin treatment affects HK-2 gene expression profiles. Histogram representation of the number of differentially expressed genes
GO classification of DEGs affected by DAP
To further investigate the biological function of DAP in HK-2 cells, a total of 349 DEGs between the control and DAP treatment groups were classified by GO terms, including biological process, cellular component, and molecular function. For biological processes, regulation of transcription, multicellular organism development, and cell differentiation were the most abundant subcategories. In terms of the cellular component category, a large number of DEGs were categorized into cell membrane, cell junction, and extracellular space. In terms of molecular function category, the DEGs were mainly classified into protein/metal ion/DNA binding and DNA-binding transcription factor activity (Fig. 3A). The top 20 GO-enriched genes with the most significant p-values are displayed in a scatter plot representing the rich factor (ratio of the number of DEGs to the total number of genes in the GO term or pathway) and the number of DEGs (Fig. 3B). The results showed that DAP significantly regulated the expression of genes associated with cell junction and integral component of membrane (Fig. 3B).
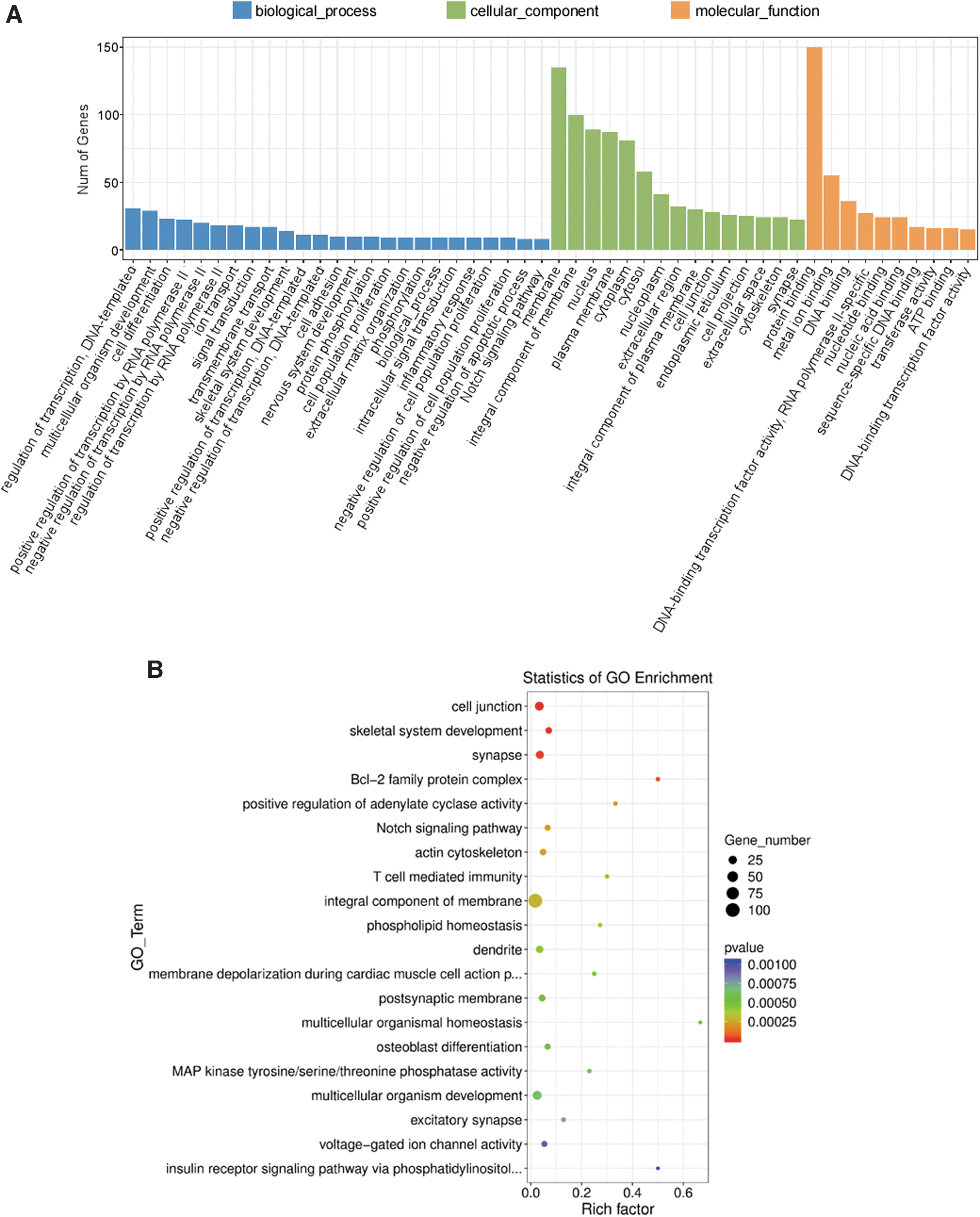
GO terms classification of DEGs in dapagliflozin-treated HK-2 cells.
DAP treatment affects gene expression profiles of cell junction and integral component of membrane
Next, we analyzed the DEGs in cell junction and integral component of membrane groups. The FPKM value, fold change, and p-value of DEGs associated with cell integral component of membrane are given in Supplementary Table S3. As shown in Figure 4A and B, a total of 100 DEGs were clustered into the integral component of membrane group, in which the expression of 89 genes was significantly downregulated and that of 11 genes was upregulated upon DAP treatment. A total of 30 DEGs were clustered into the cell junction group, in which FER and CLDN10 expression was upregulated, and the expression of 28 genes, including DAG1, KCNA2, CABP1, and EPPK1, was downregulated upon DAP treatment (Fig. 4C, D). The FPKM value, fold change, and p-value of DEGs associated with cell junction are given in Supplementary Table S4. Our results showed that DAP could downregulate the DEGs associated with the cell junction and integral component of the membrane.
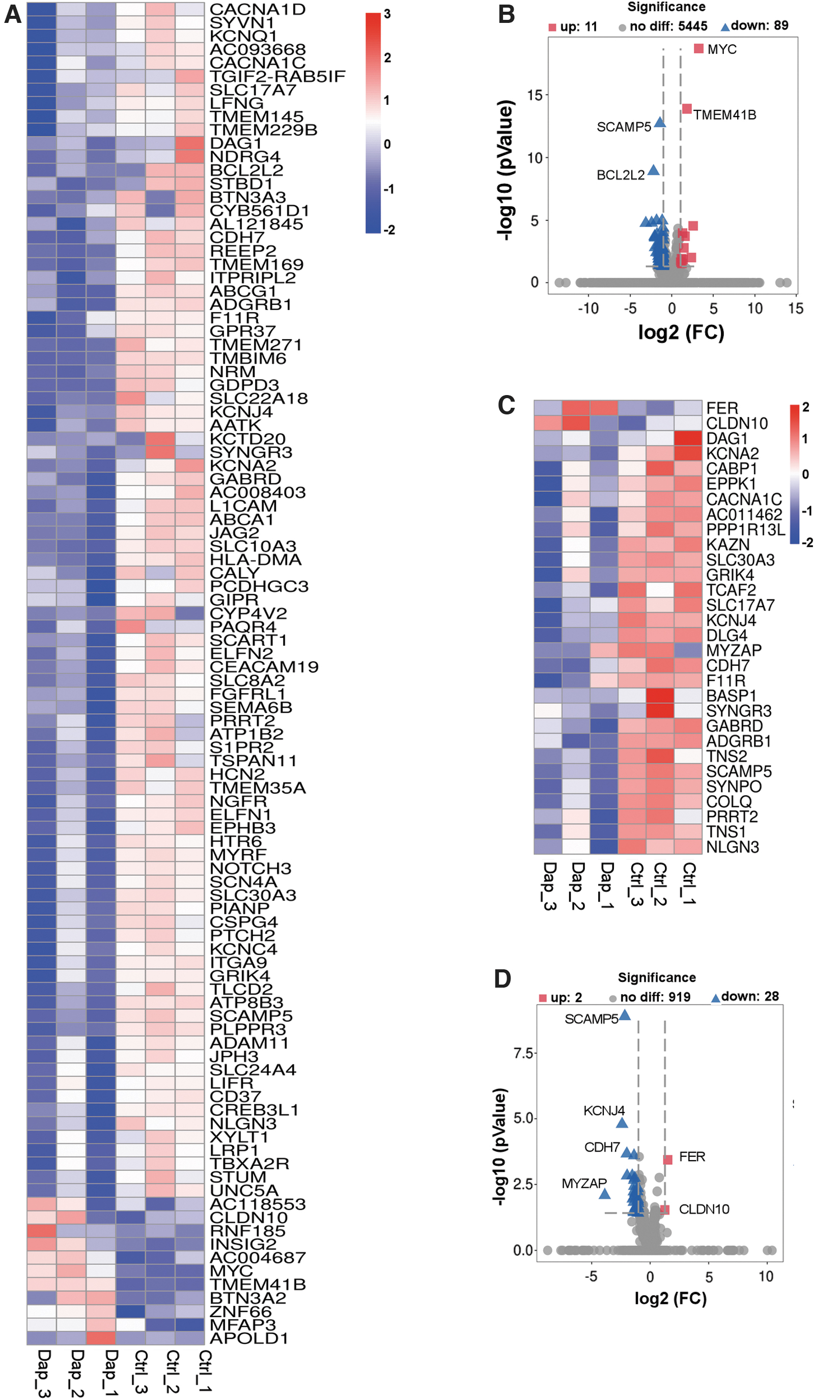
Dapagliflozin treatment affects gene expression profiles of cell junction and integral component of membrane. Heatmap showing the DEGs related to integral component of membrane
KEGG pathway analysis of DEGs affected by DAP in HK-2 cells
Next, we performed KEGG pathway enrichment analysis on the DEGs affected by DAP in HK-2 cells. The data of KEGG enrichment analysis are shown in Supplementary Table S5, and the top 20 KEGG pathways with the most significant p-values were plotted in a scatter plot (Fig. 5). The results indicated that the cAMP, MAPK, cGMP-PKG, and Hippo signaling pathways were the most significant pathways (Fig. 5A). Furthermore, several pathways were associated with cellular physiological functions, protein digestion and absorption, cell adhesion, and aldosterone synthesis and secretion.
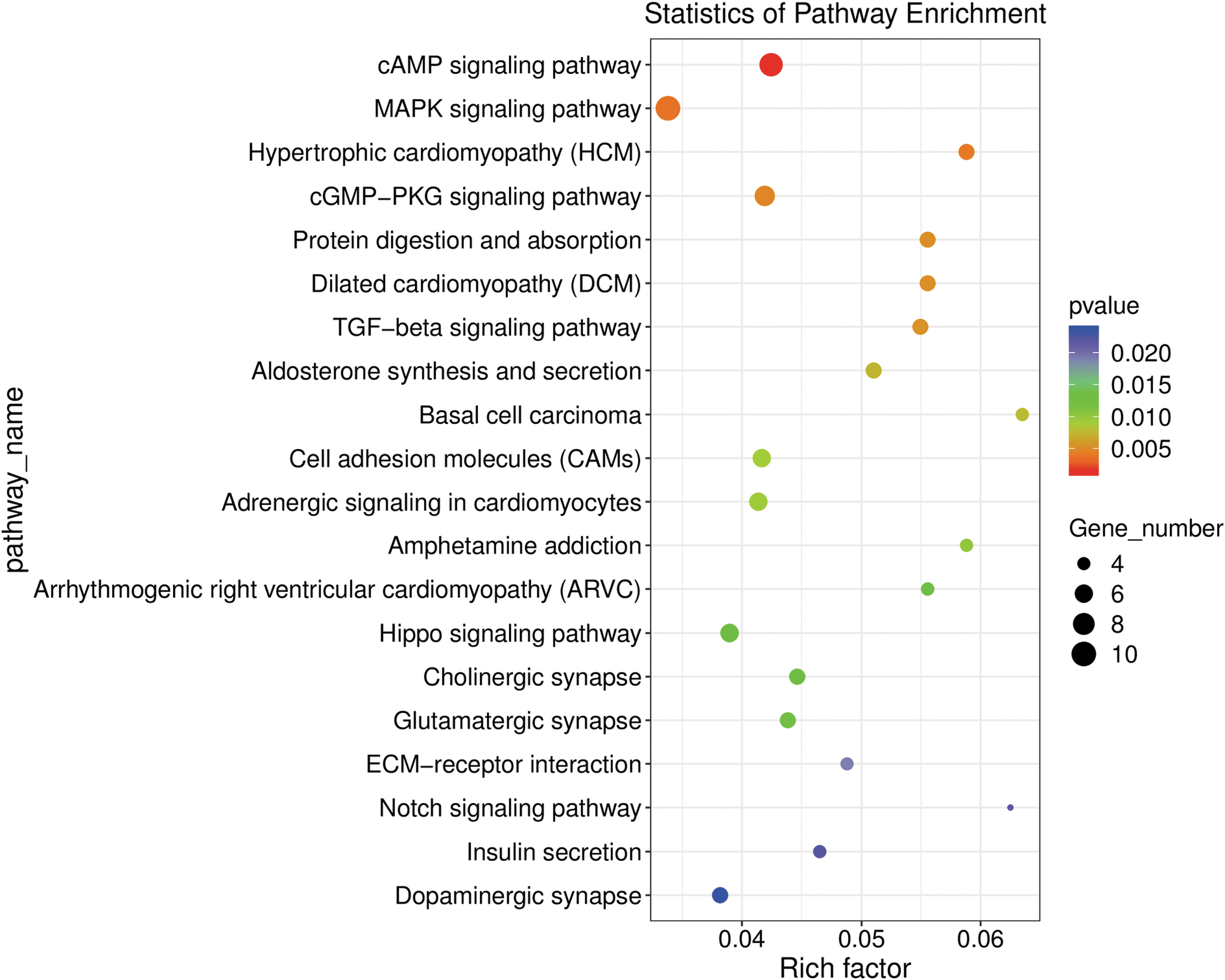
KEGG pathway enrichment analysis of DEGs regulated by dapagliflozin. Scatter plot showing the top 20 most significant KEGG pathways from the data obtained by RNA-seq after treatment of HK-2 cells with dapagliflozin for 0 or 24 h. The dot size indicates the number of DEGs enriched in the corresponding KEGG pathway. Different p-values are indicated by dot colors. The rich factor is the ratio of the number of DEGs to the total number of genes in the KEGG pathway. KEGG, Kyoto Encyclopedia of Genes and Genomes.
DAP treatment affects the expression of genes associated with cAMP, MAPK, and cGMP-PKG signaling pathway
Next, we analyzed the DEGs associated with cAMP, MAPK, and cGMP-PKG signaling pathways and found that a total of nine DEGs were clustered in the cAMP signaling pathway, CACNA1D, CACNA1C, ARAP3, HCN2, HTR6, CALML4, ATP1B2, CREB3L1, and GIPR, which were significantly downregulated after DAP treatment (Fig. 6A). We also confirmed the cAMP signaling pathway-related DEGs in HK-2 cells after DAP treatment by RT-qPCR (Fig. 6B). A total of seven DEGs were clustered in the MAPK signaling pathway, CACNA1D, CACNA1C, CREB3L1, ATP1B2, NFATC4, SLC8A2, and CALML4, which were downregulated after DAP treatment (Fig. 6C). RT-qPCR analysis also showed that the expression of CACNA1D, CACNA1C, CREB3L1, ATP1B2, NFATC4, SLC8A2, and CALML4 was downregulated in HK-2 cells after DAP treatment (Fig. 6D). As shown in Figure 6E, a total of 10 DEGs were clustered in the cGMP-PKG signaling pathway, of which 2 genes, MYC and LAMTOR3, were upregulated and 8 genes, CACNA1D, CACNA1C, DUSP8, AC011462, IGF2, DUSP4, DUSP5, and NGFR, were downregulated after DAP treatment. The cGMP-PKG signaling pathway-related DEGs affected by DAP in HK-2 cells were also confirmed by RT-qPCR (Fig. 6F). The FPKM value, fold change, and p-value of DEGs associated with cAMP, MAPK, and cGMP-PKG signaling pathway associated DEGs are shown in Supplementary Tables S6-S8. Our results demonstrated that DAP inhibited the expression of genes associated with the cAMP, MAPK, and cGMP-PKG signaling pathways.

Dapagliflozin treatment affects the expression of genes related to cAMP, MAPK, and cGMP-PKG signaling pathway.
Discussion
DN is one of the most serious complications of patients with diabetes and is often accompanied by microangiopathy of other organs or systems, such as diabetic retinopathy and peripheral neuropathy (Bellary et al., 2020). As the incidence of diabetes is increasing worldwide, DN has become the main cause of chronic kidney disease in patients with diabetes (Haraguchi et al., 2020). Therefore, early diagnosis and timely treatment of diabetes are of great significance for delaying the development of DN in these patients (Nakagawa et al., 2011). Several interventional strategies have involved multifactorial approaches, including intensive glucose and blood pressure control (Fineberg et al., 2013). SGLT-2 inhibitors can inhibit the reabsorption of glucose by the kidneys and excretion of excess glucose from the urine, thereby reducing blood glucose levels. SGLT-2 inhibitors are becoming a new class of antidiabetic drugs based on the results of large clinical trials (Chao and Henry, 2010). DAP, a specific SGLT-2 inhibitor, has been reported to decrease UACR and albuminuria and reduce the progression of kidney disease in patients with type 1 or type 2 diabetes (Mosenzon et al., 2019; Groop et al., 2020). In 2019, Pollock and his colleagues enrolled 461 patients with an increased UACR rate ranging from 30 to 3500 mg/g and found that a course of 24 weeks DAP treatment reduced UACR by 21.0% and the combination of DAP with saxagliptin decreased UACR by 38.0% (Pollock et al., 2019). In 2016, Fioretto et al. (2016) found that placebo-corrected UACR reductions (95% CI) of −57.2% (−77.1, −20.1) and −43.8% (−71.0, 9.0) occurred in the DAP 10 mg and 5 mg groups, respectively. Both Pollock's and Fioretto's groups did not discriminate between the different protective effects of DAP on lowering UACR in patients with <300 mg/g (indicating weak kidney damage) and in patients with UACR >300 mg/g (indicating severe kidney damage). In our study, however, we further separated the patients into two different groups according to the UACR level, and found that DAP was effective in inhibiting UACR levels in the 30-300 mg/g group, but had no effect on UACR level in the >300 mg/g group. Interestingly, this weak effect of DAP on patients with UACR levels >300 mg/g may not be attributed to the lower dosage of DAP we used (5 mg), as there was no significant difference between 5 and 10 mg of DAP on lowering the UACR level in Fioretto et al.'s study (Fioretto et al., 2016). However, due to the short marketing time of the SGLT-2 inhibitors in China and the few repeated follow-up cases in this study, further studies are needed to expand the sample size and increase follow-up time to verify the effect of DAP on the continuous reduction of UACR.
Inflammation plays a fundamental role in the pathophysiology of DN, and ameliorating the inflammatory response is a major therapeutic strategy for DN (Navarro-Gonzalez et al., 2011; Gnudi, 2015; Wada and Makino, 2016; Flyvbjerg, 2017; Tang and Yiu, 2020). Inflammatory mediators, including cytokines, chemokines, and matrix metalloproteinases, can recruit monocytes and macrophages to the kidneys to intensify the inflammatory response (Navarro-Gonzalez et al., 2011; Wada and Makino, 2016; Tang and Yiu, 2020). DAP has been reported to inhibit the levels of renal triglycerides, superoxide dismutase, and malondialdehyde, subsequently leading to a decrease in renal inflammation and fibrosis (El-Daly et al., 2018; Jaikumkao et al., 2018). DAP could also attenuate the progression of diabetic cardiomyopathy by inhibiting the activation of the NLRP3 inflammasome, which can repress the activation of the AMPK/mTOR complex 2 (mTORC2) in the hearts of BTBR mice (Chen et al., 2020). Other DAP-modulated signaling pathways, such as cAMP, STAT1/TGF-β1, and metabolic pathways, have also been studied. For example, Wang et al. (2017) discovered that DAP significantly prevented glucose-stimulated glucagon secretion in alpha cells by inhibiting the phosphorylation of hepatic cAMP-responsive element binding protein and reducing the expression of phosphoenolpyruvate carboxykinase 2. Furthermore, DAP not only improves hyperglycemia but also slows down the progression of diabetes-associated renal tubulointerstitial fibrosis by improving hyperglycemia-induced activation of the STAT1/TGF-β1 pathway in HK-2 cells (Huang et al., 2019). A metabolomics-based molecular pathway analysis indicated that DAP treatment significantly changed a total of 108 metabolites, among which 74 metabolites could be linked to 367 unique proteins/genes in patients with diabetic kidney disease (DKD) (Mulder et al., 2020). In this study, we found that DAP significantly downregulated the mRNA levels of PPKAR2A-AS1, FGFRL1, and GPRASP1, which are associated with inflammatory response and renal fibrosis.
Excessive glucose influx activates cellular signaling pathways, including the DAG-PKC, polyol, and hexosamine pathways, which are involved in the development of DN (Kanwar et al., 2008; Dragos et al., 2020; Haraguchi et al., 2020). Numerous studies have shown that the cAMP signaling pathway plays a key role in the pathogenesis of DN by activating inflammatory and profibrotic responses (Lagranha et al., 2007; Deb et al., 2017). It has been reported that the MAPK signaling pathway is activated during the course of DN, which means that the MAPK signaling pathway can promote the development of DN (Wang et al., 2006). It has been established that the pathogenesis of DN is associated with abnormal activity of nitric oxide-cGMP-PKG signaling, which plays a central role in the regulation of diverse processes, including smooth muscle relaxation, inflammation, and fibrosis (Vidyashankar et al., 2014). In our study, we found that DAP significantly downregulated genes related with cAMP, MAPK, and cGMP-PKG signaling pathways, such as CACNA1D, CACNA1C, ATP1B2, and IGF2. This implied that DAP could inhibit the inflammatory and profibrotic responses by inhibiting the cAMP, MAPK, and cGMP-PKG signaling pathways and attenuate the pathogenesis of DN.
Others have performed RNA-seq on HK-2 cells but not in the context of DAP exposure, which represents the specific effects of DAP on HK-2 cells. For example, in a recent study, Hickson et al. (2021) discovered that DKD-mesenchymal stem/stromal cells medium protected high glucose plus transforming growth factor-β-exposed HK-2 cells by reducing apoptotic, fibrotic, and inflammatory marker expression. Pyo et al. (2021) used transcriptome analysis to reveal that ochratoxin A significantly affected expression of genes associated with hypoxia, epithelial-mesenchymal transition, apoptosis, and xenobiotic metabolism in HK-2 kidney cells through the regulation of AhR, Smad2/3, and HIF-1α pathways. However, because SGLT-2 is not exclusively expressed by renal tubular epithelial cells, and glucose reabsorption is not necessarily aided by only renal tubular epithelial cells, it is a weakness of our study to interpret our data based on the behavior of only one cell type. Therefore, additional research is required to confirm and expand our results.
In conclusion, DAP could reduce UACR and inflammatory and profibrotic responses in patients with DN, which may be attributed to the ability of DAP to downregulate the expression of genes related to cAMP, cGMP-PKG, and MAPK signaling pathways. Our results provide a novel insight into the mechanism through which DAP reduces urinary albumin excretion in patients with DN and provide a theoretical basis for the clinical treatment of DN.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Statement of Licensing Committee and Institution
The human study was reviewed and approved by the ethics committee of Sir Run Run Shaw Hospital of Zhejiang University School of Medicine and conformed to institutional guidelines.
Footnotes
Authors' Contributions
J.Z. conceived the project and designed the experiments. G.C. and H.W. conducted the experiments and G.C. wrote the article. J.Z. revised the article. All authors have reviewed and approved the final article.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.