
Abstract
Aims:
Gout is a form of inflammatory arthritis characterized by the deposition of monosodium urate crystals. An important risk factor for gout is hyperuricemia. The relationship between SLC22A12 gene variants and the susceptibility to hyperuricemia has been reported, but these findings have been inconsistent. Thus, we aimed to assess the relationship between SLC22A12 gene variants and hyperuricemia susceptibility through a meta-analysis.
Methods:
The meta-analysis was performed by searching PubMed, Embase, Web of Science, and Chinese National Knowledge Infrastructure (CNKI) databases. The relationship between hyperuricemia risk and the SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, rs893006, and rs475688 variants was assessed by odds ratios and 95% confidence intervals.
Results:
In total, 20 eligible publications with 4817 cases and 6819 controls were included in the meta-analysis. Hyperuricemia risk was significantly associated with the SLC22A12 alleles rs3825018, rs7932775, and rs475688 under both the dominant and recessive models and with rs3825016 under the allelic and dominant models.
Conclusions:
Under the allelic model SLC22A12 rs3825018 and rs3825016 were risk factors for hyperuricemia and gout as was rs7932775 under dominant and recessive models, while the SLC22A12 rs475688 was protective against hyperuricemia under both dominant and recessive models.
Introduction
Gout is a heterogeneous disease comprising a purine metabolic disorder and hyperuricemia (Cao et al., 2021). Elevated blood uric acid levels are the material basis of gout. About 90% cases of primary hyperuricemia are associated with decreased excretion of uric acid (Narang and Dalbeth, 2020). The prevalence of hyperuricemia in the elderly is up to 20% (Luk and Simkin, 2005). Clinical and genetic epidemiology data show that environmental and genetic factors can combine to lead to hyperuricemia (Merriman, 2015). In addition to uric acid derived from dietary sources, the molecular mechanisms underlying the increased synthesis of uric acid are well understood (Min and Junwu, 2016).
Genetic factors play an important role in the development of primary hyperuricemia; researchers have estimated that the genetic influence of uric acid is 40-70% (Voruganti et al., 2009). To date, genome-wide association studies have targeted susceptibility loci that account for only about 7% of the variation in blood uric acid levels (Kolz et al., 2009; Cho et al., 2020). Genes involved in the reabsorption and secretion of uric acid by the kidney, also known as the urate transporters, are located on the lumen membrane and the basement membrane of the proximal renal tubules (Kon et al., 2019). The urate transporter genes that play a major regulatory role in blood uric acid homeostasis are SLC22A12, SLC2A9, and ABCG2 (Tu et al., 2018). SLC22A12 was the first urate transporter gene to be identified, and it encodes the hURAT1 protein, which has organic ion transporter homology and is expressed in the apical membrane of the renal tubules (Komatsuda et al., 2006). Graessler et al. (2006) have suggested that SLC22A12 gene variants are related to increased blood uric acid levels and reduced urate excretion.
The exon 2 rs11231825 may be related to the decrease in the uric acid excretion rate. The exon 1 rs3825016 can increase uric acid excretion and reduce uric acid reabsorption (Graessler et al., 2006). Shima et al. (2006) found that the SLC22A12 rs893006 variant was an independent susceptibility factor for hyperuricemia in a Japanese population; this locus was also verified in a male Chinese Han population (Guan et al., 2009). Meng et al. (2010) found that single nucleotide polymorphisms (SNPs) of the exon 8 rs7932775 and intron 8 rs7929627 of the SLC22A12 gene were closely related to primary hyperuricemia. Li et al. (2010) found that the variation in intron 3 rs161109885 and exon 8 rs7932775 of the SLC22A12 gene was significantly correlated with the uric acid level in a Chinese Han population.
However, this evidence is inconclusive with regard to the genetic associations between SLC22A12 gene variants and hyperuricemia susceptibility in populations of different ethnicities owing to the limited sample size of the studies. Meta-analysis is a powerful tool to obtain more conclusive evidence. Thus, we performed a meta-analysis by including eligible studies from electronic databases to further evaluate the genetic associations between SLC22A12 gene variants and hyperuricemia susceptibility.
Materials and Methods
Literature search
Studies were retrieved from online databases, such as the PubMed, Web of Science, Embase, and Chinese National Knowledge Infrastructure (CNKI) databases, with the following key words: “Human urate transporter 1” or “hURAT1” or “SLC22A12” and “polymorphism” or “single nucleotide polymorphisms” or “variation” or “SNPs” and “gout” or “hyperuricemia” (updated on April 26, 2021). At the same time, the references were included, and related reviews were assessed. There were no limitations on language.
Selection criteria
Inclusion criteria were as follows: (1) case-control or cohort study design; (2) documentation of the genetic association between SLC22A12 gene variants and hyperuricemia susceptibility; (3) available data on allelic or genotypic frequency.
Exclusion criteria were as follows: (1) duplicated study or review, letter, short communication, abstract; (2) not a case-control or cohort study design; (3) irrelevant to the genetic associations between SLC22A12 gene variants and hyperuricemia susceptibility; (4) lack of data on allelic or genotypic frequency.
Data extraction
W.K. and Q.H. independently carried out the data extraction. Disputes was resolved by discussion. The following information were extracted from the eligible studies: name of the first author, publication year, ethnicity, mean ages of study subjects, proportion of male subjects, genotyping methods, number of cases and controls, distribution of alleles, and genotypes of variants in cases and controls. The Newcastle-Ottawa Scale (NOS) was used to evaluate the included studies (Han et al., 2017). The quality scores ranged from 0 to 9. Studies were included if their NOS score was ≥6.
Statistical analysis
Combined odds ratios (ORs) and 95% confidence intervals (CIs) were employed to evaluate the genetic associations between SLC22A12 gene variants and hyperuricemia susceptibility. The Z test was used to assess the significance of the combined ORs. The interstudy heterogeneity was evaluated by Cochrane's Q-statistic and I2-statistic; I2% > 50 and p < 0.05 indicated the existence of interstudy heterogeneity. Furthermore, the random-effects model or a fixed model were used. Subgroup analysis was conducted on stratification by ethnicity in the present meta-analysis. Egger's test and Begg's test were used to evaluate and analyze the publication bias of each study. Meta-regression analysis was used to analyze potential sources of interstudy heterogeneity. p < 0.05 was considered significant. Data were processed with RevMan 5 (Oxford, United Kingdom) and STATA12.0 (StataCorp, College Station, TX).
Results
Characteristics of the eligible studies
On retrieving studies from the databases, 658 publications were identified. According to the selection criteria, the following studies were excluded: 293 studies that were duplicated study or review, letter, short communication, or abstract; 189 studies that were not case-control or cohort studies in design; and 160 studies that did not document the correlation between SLC22A12 gene variants and hyperuricemia susceptibility. Finally, 16 studies were included in the present study (Fig. 1) (Graessler et al., 2006; Wang et al., 2007, 2012; Han et al., 2010, 2012; Li et al., 2010, 2018, 2019; Meng et al., 2010; Tu et al., 2010; Flynn et al., 2013; Yakut et al., 2013; Lu et al., 2017; Zhao et al., 2017; Lin et al., 2018; Duong et al., 2019). Of these studies, one study contained four different populations and was treated as four individual studies (Flynn et al., 2013); another contained two different populations and was treated as two individual studies (Tu et al., 2010). Thus, 20 individual studies were included in the present meta-analysis.

Preferred Reporting Items for Systematic Reviews and Meta-Analyses flow chart of studies inclusion and exclusion.
Of the included studies, 13 studies were conducted in Asian populations (Graessler et al., 2006; Wang et al., 2007, 2012; Han et al., 2010, 2012; Li et al., 2010, 2018, 2019; Meng et al., 2010; Yakut et al., 2013; Lu et al., 2017; Zhao et al., 2017; Lin et al., 2018) and 3 studies were conducted in Caucasian populations (Tu et al., 2010; Flynn et al., 2013; Duong et al., 2019). Furthermore, 12 articles reported the relationship between SLC22A12 gene variants and hyperuricemia risk (Graessler et al., 2006; Han et al., 2010, 2012; Li et al., 2010; Meng et al., 2010; Wang et al., 2012; Lu et al., 2017; Zhao et al., 2017; Lin et al., 2018), while 4 articles reported the relationship between SLC22A12 gene variants and gout risk (Wang et al., 2007; Yakut et al., 2013; Li et al., 2018, 2019). The NOS scores of the included studies were higher than 6 (Table 1).
The Information of Eligible Studies Included in the Present Meta-Analysis
/, No data; NOS, Newcastle-Ottawa Scale; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism; i MLDR, improved Multiplex Ligase Detection Reaction.
Results of the meta-analysis
Relationship between hyperuricemia risk and SLC22A12 rs11602903, rs524023, and rs3825018
Table 2 presents a summary of the reported associations between SLC22A12 rs11602903, rs524023, and rs3825018 variants and the risk of hyperuricemia. No association was found between hyperuricemia susceptibility and all genetic models of SLC22A12 rs11602903 and rs524023 (p > 0.05) (Figs. 2A, 2C, 3A, 3C, 4A, 4C, and Table 2). A significant association was observed between hyperuricemia risk and the allelic model but not the dominant and recessive models of SLC22A12 rs3825018 (p = 0.007, OR [95% CI] = 1.34 [1.08 to 1.67]) (Figs. 2B, 3B, 3C, and Table 2). Subgroup analysis based on ethnicity showed that the allelic and dominant models of SLC22A12 rs11602903 were significantly associated with hyperuricemia risk in both Asian and Caucasian populations (p < 0.05) (Table 2). However, a significant relationship was found between hyperuricemia risk and the allelic model of SLC22A12 rs3825018 in Asian populations (p = 0.0001), but not in Caucasian populations (p = 0.26) (Table 2).

Forest plots of odds ratios for the association between the allelic model of SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, and rs893006 and rs475688 variants and hyperuricemia risk.
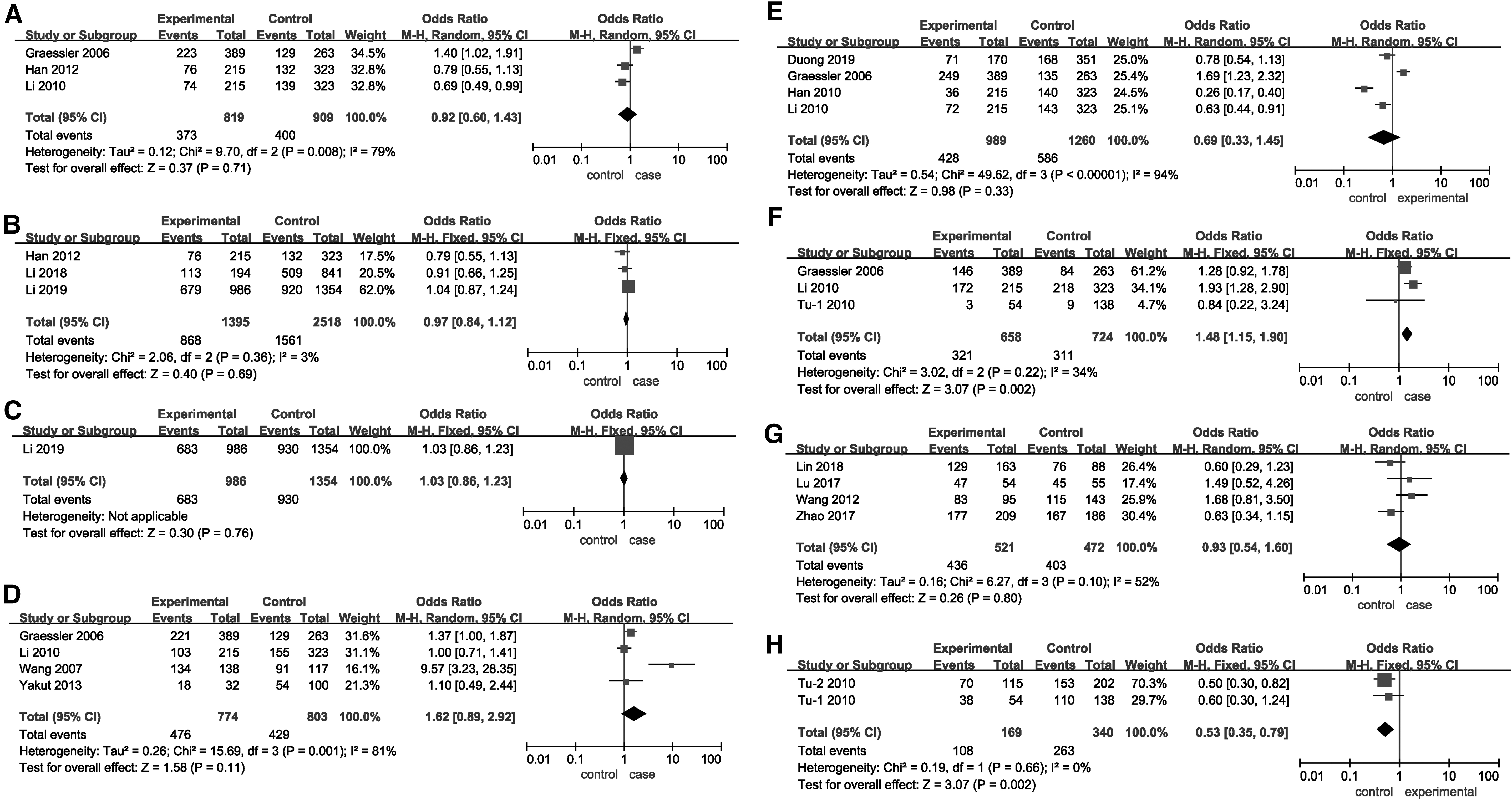
Forest plots of odds ratios for the association between the dominant model of SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, and rs893006 and rs475688 variants and hyperuricemia risk.

Forest plots of odds ratios for the association between the recessive of SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, and rs893006 and rs475688 variants and hyperuricemia risk.
The Genetic Associations Between the SLC22A12 rs11602903, rs524023, and rs3825018 and Risk of Hyperuricemia
—, Not available; CI, confidence interval; F, fixed model; OR, odds ratio; R, random model.
Relationship between hyperuricemia risk and SLC22A12 rs3825016, rs11231825, and rs7932775
No correlation was detected between hyperuricemia risk and all genetic models of SLC22A12 rs11231825 (p > 0.05) (Figs. 2E, 3E, 4E, and Table 3). Significant associations were found between hyperuricemia risk and the allelic and dominant models of SLC22A12 rs3825016 (Figs. 2D and 3D) as well as the dominant and recessive models of SLC22A12 rs7932775 (p < 0.05) (Fig. 3F, 4F, and Table 3). Subgroup analysis based on ethnicity showed that the allelic and dominant models of SLC22A12 rs3825016 were associated with hyperuricemia risk in Asian populations; data were insufficient in Caucasian populations (p < 0.05) (Table 3). In addition, the allelic model of SLC22A12 rs7932775 was associated with hyperuricemia risk in Caucasian populations, while the dominant model of SLC22A12 rs7932775 was associated with hyperuricemia risk in Asian populations. The recessive model of SLC22A12 rs7932775 was associated with hyperuricemia risk in both Caucasian and Asian populations.
The Genetic Associations Between the SLC22A12 rs3825016, rs11231825, and rs7932775 and Risk of Hyperuricemia
Relationship between hyperuricemia risk and SLC22A12 rs893006 and rs475688
There was no significant association between the risk of hyperuricemia and genetic models of SLC22A12 rs893006 (p > 0.05) (Figs. 2G, 3G, and 4G, and Table 4). Both dominant and recessive models of SLC22A12 rs475688 were related to hyperuricemia risk (p < 0.05) (Fig. 2H, 3H, 4H, and Table 4). Subgroup analysis based on ethnicity showed that the allelic and dominant models of SLC22A12 rs475688 were associated with hyperuricemia susceptibility in Asian populations (p < 0.05), while the recessive model of SLC22A12 rs475688 was associated with hyperuricemia susceptibility in Caucasian populations (p < 0.05).
The Genetic Associations Between the SLC22A12 rs893006 and rs475688 and Risk of Hyperuricemia
Heterogeneity
Heterogeneity was found for all eight variants in the overall analysis. Thus, subgroup analysis was conducted based on ethnicity. However, significant heterogeneity was still detected in the overall analysis. Significant heterogeneity in SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, rs893006, and rs475688 polymorphisms were primarily presented by Graessler et al. (2006), Meng et al. (2010), Tu et al. (2010), Han et al. (2012), Lin et al. (2018), Duong et al. (2019), and Li et al. (2019). No significant heterogeneity was detected after removal of these studies (I2 < 50%, p > 0.05).
Sensitivity analysis and publication bias
There was no deviation from the Hardy-Weinberg equilibrium or OR 3.0 in the 20 included studies on SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, rs893006, and rs475688 variants in the present meta-analysis. Thus, the associations between hyperuricemia risk and SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, rs893006, and rs475688 were stable and reliable. As shown in Table 5, Figure 5, and Figure 6, no evidence of publication bias was found on using Begg's test and Egger's test.
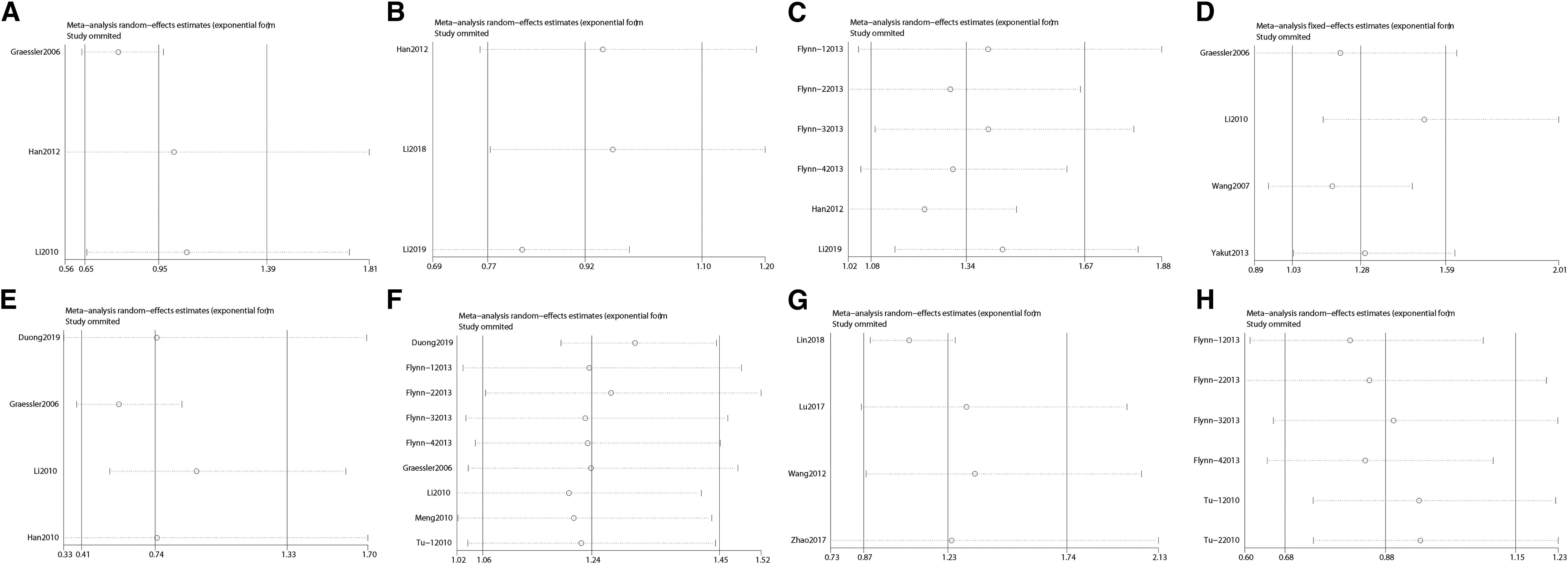
Sensitivity analyses between allelic models of SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, and rs893006 and rs475688 variants and hyperuricemia risk.
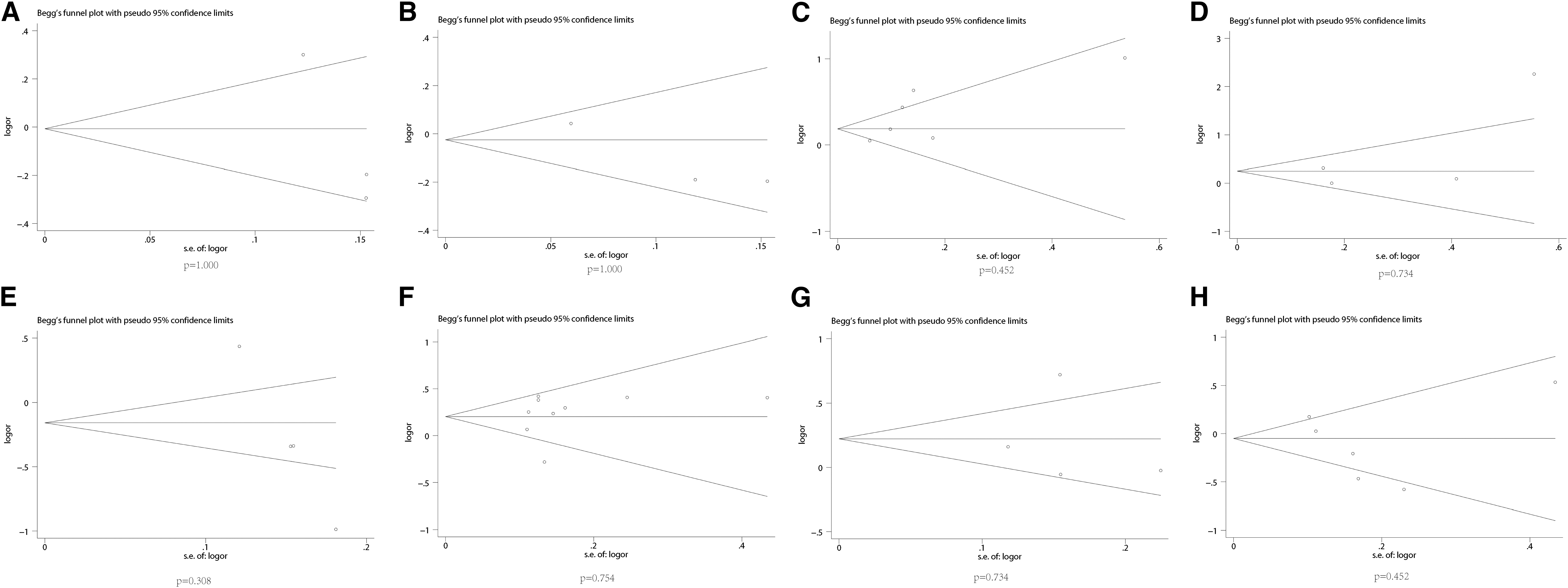
Publication bias of literature for SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, and rs893006 and rs475688 variants were tested by Begg’ s funnel plot.
The Egger's Test for SLC22A12 Polymorphisms
SE, standard error.
Discussion
An increasing number of case-control studies have begun to explore the role of genetic variants in the development of hyperuricemia. However, evidence of the relationship between hyperuricemia risk and SLC22A12 rs11602903, rs524023, rs3825018, rs3825016, rs11231825, rs7932775, rs893006, and rs475688 variants remains inconsistent. The differences in the results may be attributed to the small sample size of a single study. In the present study, we found that the allelic models of SLC22A12 rs3825018 and rs3825016 and the dominant and recessive models of SLC22A12 rs7932775 significantly increase the risk of hyperuricemia, while the dominant and recessive models of SLC22A12 rs475688 significantly decrease the risk of hyperuricemia.
SLC22A12 is a gene that is associated not only with hyperuricemia and gout but also with hypouricemia. Eraly et al. (2008) and Hosoyamada et al. (2010) have prepared SLC22A12 knockout mice, and found that renal uric acid resorption was impaired in the knockout mice, confirming the effect of URAT1 encoded by SLC22A12 on urate resorption. At the same time, a series of studies have found that SNPs and mutations in the SLC22A12 gene may also contribute to the development of type 1 hereditary hypouricemia (RUC1) (Iwai et al., 2004; Komoda et al., 2004). The SLC22A12 rs121907892 and rs121907896 were the common mutations in Japanese and Korean RHUC1 populations (Iwai et al., 2004; Komoda et al., 2004; Ichida et al., 2010). Moreover, the SLC22A12 rs200104135 and exon 7 delGGCAGGGCT with the highest frequency (5.6% and 1.9% separately) of predominant dysfunctional variants were identified in the Roma populations (Gabrikova et al., 2015; Stiburkova et al., 2016). While, there was null allele frequency in African American, Ashkenazi Jewish, South Asia, and European population (Slatkin, 2004; Miyauchi et al., 2021).
Furthermore, the deletion of A51fsX64 on exon 1 of SLC22A12 gene was reported to be associated with renal dysfunction in two independent Chinese families (Li et al., 2013). However, a recent study addressing a cohort of gout/hyperuricemia in the Czech population did not find URAT1 variants (Olha et al., 2014). The mutations of SLC22A12 gene are associated with hyperuricemia, hypouricemia, and gout-related diseases. The mutation region of SLC22A12 gene may be located in exon, intron, and promoter region, and the genetic predisposition of the gene varies from country to country and nation to nation. Therefore, it is necessary to further study the mutation site, SNP, and its adjacent genes in different ethnic groups. In the present study, we confirmed that the allelic models of rs3825018 significantly increased the risk of hyperuricemia in Asian populations but not in Caucasian populations. Differences in ethnicity may be responsible for the different genotypic distribution frequencies between Asian and Caucasian populations. The promoter region is where polymorphic loci of various genes are concentrated, and polymorphic loci in this region can enhance or weaken protein function by changing the binding activity of the promoter region.
Functional tests of the SLC22A12 gene promoter region showed that the minimum functional promoter region of the SLC22A12 gene was located from −253 to −39 bp. There are two activated protein 1 (AP-1) transcription factor-binding sites, one hepatic nuclear factor L (HNF-1) transcription factor-binding site, and one CCAAT/enhancer-binding protein B (CEBPP) transcription factor-binding site between −339 and −35 bp. It has been suggested that there are many cis-acting elements between −339 and −35 bp that affect the transcription activity of the SLC22A12 gene, and a change in a single base in this sequence may lead to changes in the gene transcription activity. The three polymorphic loci included in this study were located around the minimal promoter region of the SLC22A12 gene. It may be that the change caused by a variant led to changes in the transcriptional activity of SLC22A12 or the combined changes at several loci led to changes in the expression of hURAT1 (Tao et al., 2015).
The SLC22A12 rs3825016, rs11231825, and rs7932775 variants are located at the exon 1, 2, and 8, respectively. The rs3825016 variant can increase the uric acid excretion rate and reduce uric acid reabsorption. Iwai et al. (2004) found that rs3825016 was not associated with serum uric acid levels in a Japanese population and its genotypic distribution frequency in this population was significantly different from that in a German population (Graessler et al., 2006), which indicated that there were ethnicity-related differences in the SLC22A12 gene variant. In our meta-analysis, the allelic and recessive models of rs3825016 significantly increase hyperuricemia risk, which was similar to the results reported by Ma et al. (2014) and Zou et al. (2018). Furthermore, rs11231825 was shown to be related to the uric acid excretion rate. Among the four studies conducted on the association between rs11231825 and hyperuricemia risk, three showed that rs11231825 as significantly related to hyperuricemia and gout susceptibility. However, we found that genetic models of rs11231825 were not associated with hyperuricemia risk when results were combined, even in a subgroup analysis based on ethnicity.
Thus, rs11231825 might not be a risk factor for hyperuricemia and gout. The rs7932775 variant was first detected by sequencing in both Japanese (Iwai et al., 2004) and German populations (Graessler et al., 2006) and was revealed to encode a synonymous alteration in the protein sequence. In previous studies, five reported that rs7932775 was significantly associated with hyperuricemia risk, while others reported negative results. In our meta-analysis, we found that the dominant and recessive models of rs7932775 could significantly increase hyperuricemia risk, similar to the results reported by Ma et al. (2014). Moreover, we performed a subgroup analysis based on ethnicity and found that the allelic model of rs7932775 was a risk factor for hyperuricemia in Asian populations, the dominant model of rs7932775 was a risk factor for hyperuricemia in Caucasian populations, and the recessive model of rs7932775 was a risk factor for hyperuricemia both in Asian and Caucasian populations. These inconsistencies may be due to the different genetic background in Asian and Caucasian populations.
The rs893006 locus is at intron 4 in the SLC22A12 gene. Studies have shown that the rs893006GG genotype may be related to elevated blood uric acid levels (Ren and Hu, 2014). Guan et al. (2009) showed that the rs893006 variant may be a risk factor for Chinese male patients with hyperuricemia. Shima et al. (2006) found that the uric acid concentration of the GG genotype was higher compared with the GT and TT genotypes in Japanese men, suggesting that the GG genotype was an independent influencing factor for hyperuricemia. Zhao et al. (2017) showed that the G allele at rs893006 was a protective factor, and the risk of hyperuricemia with the GG+GT genotype was 0.405 times higher compared with the TT genotype. Lu et al. (2017) showed that there was no significant difference in the three genotypes between the two groups of a case-control study. These differences in the results may be related to regional, racial, and ethnicity-related differences.
Notable, genetic association was detected between the SLC22A12 intron variants and no nonsynonymous variants and hyperuricemia risk. Increasing number of studies have suggested that the effect of polymorphisms or variants in noncoding region might be responsible for aberrant splice processes or be linked to some other functional polymorphisms in gene (Baralle and Baralle, 2005; Buratti et al., 2006). In addition, major urate transporters, ABCG2, SLC22A12, and SLC2A9, act as a complex mechanism in the proximal kidney tubules, and it is very likely that the impaired function of one transporter could be compensated for by one or more of the other proteins (Nakanishi et al., 2013).
Heterogeneity among different studies is a key issue in meta-analysis. Therefore, this study additionally aimed to explore the potential heterogeneity among different studies. Although the results included in this meta-analysis were adjusted for common influencing factors such as ethnicity, some unstudied or unknown factors may be the source of heterogeneity. The meta-regression analysis showed that potential sources of heterogeneity (race and disease type) had no effect on the results. Thus, heterogeneity between studies may be caused by other factors such as lifestyle and environmental factors.
This study had some limitations. First, the number of included studies and the associated sample size were relatively small, especially for the rs11602903, rs524023, and rs3825016 variants. Thus, future studies should more rigorously confirm the associations between these three variants and hyperuricemia risk. Second, environmental factors, including age, gender, and dietary habits, have been shown to involve in the pathogenesis of hyperuricemia and gout. However, we could not perform further stratification analyses based on these factors owing to limited data. Third, most of the studies were conducted in Asian or Caucasian populations.
While genetic background was considered to be an important factor in the association between SLC22A12 gene variants and hyperuricemia risk, subgroup analysis was only conducted on classification into Caucasian and Asian populations. We failed to detect associations between SLC22A12 gene variants and hyperuricemia risk in other ethnicities, such as African and Latin American populations. In conclusion, the allelic models of SLC22A12 rs3825018 and rs3825016 and the dominant and recessive models of SLC22A12 rs7932775 were risk factors for hyperuricemia, whereas the dominant and recessive models of SLC22A12 rs475688 were protective factors for hyperuricemia.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.