
Abstract
Introduction:
Cytokines are cell signaling glycoproteins that are particularly important in immunity and inflammatory responses. Therefore, variations, such as single nucleotide polymorphisms (SNPs), in genes encoding for cytokines may have important consequences for their roles in health.
Materials and Methods:
A total of 222 unrelated, healthy, and un-admixed Malays (n = 97), Chinese (n = 77), and Indians (n = 48) with a median age of 30 years old (range 21-50) were typed for 22 cytokine gene SNPs: IL-1α −889 T/C, IL-1β (−511 T/C, +3962 T/C), IL-1R pst1 1970 T/C, IL-1RA mspa1 11100 T/C, IL-4Rα +1902 G/A, IL-12 − 1188 C/A, IFN-γ +874 A/T, TGF-β (cdn 10 C/T, cdn 25 G/C), TNF-α (−308 A/G, −238 A/G) IL-2 (+166 G/T, −330 T/G), IL-4 (−1098 T/G, −590 T/C, −33 T/C), IL-6 (−174 C/G, nt565 G/A), and IL-10 (−1082 G/A, −819 C/T, −592 A/C). This involved using well-established polymerase chain reaction procedures with sequence-specific primers and restriction fragment length polymorphism methods.
Results:
The majority of the screened cytokine gene SNPs are polymorphic in all three ethnicities. Exceptions include TGF-β cdn 25 (G/C), IL-1β +3962 (T/C), and TNF-α −238 (A/G), which were all observed to be monomorphic in Malays, Chinese and Indians. Many of the analyzed cytokine gene SNP genotypes deviated from Hardy-Weinberg equilibrium and the three ethnic study groups were all well-separated from reference Asian, African and European populations in a principal component analysis plot.
Conclusion:
We successfully typed 22 SNPs in 13 cytokine genes from genetic material collected from unrelated and un-admixed Malay, Chinese and Indian individuals in Peninsular Malaysia. These new cytokine gene population datasets reveal interesting contrasts with other populations.
Introduction
Malaysia is a multi-ethnic country located in Southeast Asia with Malays, Chinese, and Indians as the major population groups. Other residents include the three indigenous Orang Asli groups (Proto-Malays, Senoi, and Semang) who arrived in Peninsular Malaysia via several waves of ancient migrations (Aghakhanian et al, 2015). The earliest was by the Orang Asli Semang, a group of people (i.e., Bateq, Kintak, Lanoh, Jahai, Mendriq, and Kensiu) representing the first wave of “Out of Africa” migration and who first arrived in Peninsular Malaysia ∼50-70 kya (Aghakhanian et al, 2015).
These ancient settlements were then followed by the arrival of Orang Asli Senoi (i.e., Jahut, Mah Meri, Che Wong, Semai, Semoq Beri, and Semai) and Orang Asli Proto-Malays (i.e., Temuan, Semelai, Orang Kuala, Orang Kanaq, Seletar, and Jakun) ∼4000 and ∼3000 years ago, respectively (Hill et al, 2006). The Orang Asli Senoi speak Austroasiatic language and originated from mainland Asia, whereas Orang Asli Proto-Malays migrated south from Taiwan via the Philippines and Borneo to Peninsular Malaysia (Fix, 1995).
The Malays represent 60% of the Malaysian population and are genetically inferred to have arisen from Proto-Malays with admixture components from other ethnicities, including Chinese and Indians who settled in Peninsular Malaysia after large-scale migration during the British colonial era in the 19th century (Norhalifah et al, 2016a). It is important to note that a small number of Chinese and Indian immigrants have arrived and have significant contact with the earlier inhabitants in Peninsular Malaysia long before the colonial era.
This is evident by the influence of South Indian cultures in the ancient Kedah Empire, which was established from the 5th to the 13th century (Kumaran, 2008; O'Reilly, 2006). During this era, international traders from China, India, Sri Lanka, Europe, and Middle East came to Kedah, which replaced earlier entreports (e.g., Funan and Langkasuka Empires from the 2nd to the 6th century) in the Southeast Asia region (Hussien et al, 2007; Mukherjee, 2011; Wade, 2009). International traders including from China and India continue to trade in Peninsular Malaysia after the fall of Kedah Empire and this was largely due to the emergence of new maritime trade routes via Melaka, Sunda, and Java Straits (Hussien et al, 2007).
Over the past 15 years, polymorphic markers in genetic materials collected from individuals representing various population groups in Malaysia have been characterized using well-established serological and molecular approaches. These include blood group (e.g., ABO, Rhesus, MNS, and Kell), human neutrophil antigen, human platelet antigen, haemochromatosis, and chemokine genes where significant differences were observed between Malays, Chinese, Indians, and the various Orang Asli groups (Hajar et al, 2020a, 2020b, 2021; Jamaludin et al, 2020; Manaf et al, 2016; Syafawati et al, 2016).
These observations were attributed to their unique population histories described earlier and local selection forces in Peninsular Malaysia. Such earlier studies have not only helped to elucidate population history in the country, but also helped for health. For example, risk profiles for developing hereditary hemochromatosis are relatively higher among Indians because susceptibility genotype to this disease (compound heterozygote for C282Y and H63D in haemochromatosis gene) was only detected in this ethnic group (Hajar et al, 2021). In contrast, Malaysian Chinese individuals have a lower frequency of KIR2DL1 ligand (C2+HLA) than the Malays, which offer them some protection from preeclampsia (Tao et al, 2021).
In this study, we characterized 22 single nucleotide polymorphisms (SNPs) located in 13 cytokine genes using polymerase chain reaction with sequence-specific primers (PCR-SSP) or polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) techniques. These SNPs are uniquely distributed in worldwide populations (Gonzalez-Galarza et al, 2011) and have been shown to be clinically important in disease pathogenesis (Gulati et al, 2016).
These medical conditions include a wide range of autoimmune diseases and susceptibility to cancer and infection (Attar et al, 2017; Sayad, 2014; Scarel-Caminaga et al, 2003). Therefore, cytokine population data for Malays, Chinese, and Indians obtained from the present survey is anticipated to provide a valuable source of information for population and medical genetic research.
Materials and Methods
We recruited a total of 222 unrelated, healthy, and un-admixed volunteers, that is, individuals with three generations having no history of inter-marriage with any other ethnicity. These were from Malay (n = 97), Chinese (n = 77) and Indian (n = 48) ethnic groups. Each volunteer gave their written informed consent, and the present study was reviewed and approved by the Human Ethical Committee, Universiti Sains Malaysia (USM/JEPeM/16050191). Sampling locations were as described elsewhere (Hajar et al, 2020a).
In this study, we characterized 22 SNPs in 13 cytokine genes, namely IL-1α −889 (T/C), IL-1β −511 (T/C), IL-1β +3962 (T/C), IL-1R pst1 1970 (T/C), IL-1RA mspa1 11100 (T/C), IL-4Rα +1902 (G/A), IL-12 − 1188 (C/A), IFN-γ +874 (A/T), TGF-β cdn 10 (C/T), TGF-β cdn 25 (G/C), TNF-α −308 (A/G), TNF-α −238 (A/G), IL-2 + 166 (G/T), IL-2 − 330 (T/G), IL-4 − 1098 (T/G), IL-4 − 590 (T/C), IL-4 − 33 (T/C), IL-6 − 174 (C/G), IL-6 nt565 (G/A), IL-10 − 1082 (G/A), IL-10 − 819 (C/T), and IL-10 − 592 (A/C).
Genomic DNA templates were isolated from whole blood using the Invisorb® Spin Forensic Kit (STRATEC Molecular, Berlin, Germany), as described earlier (Hajar et al, 2020a). Genotyping of the 22 SNPs was performed using previously reported PCR-SSP or PCR-RFLP techniques; refer Table 1.
List of Methods Used for Amplification of 22 Single Nucleotide Polymorphisms in 13 Cytokine Genes (Pro-Inflammatory: IL-1α, IL-1β, IL-1R, IL-4Rα, IL-12, IFN-γ, TNFα, and IL-2; Anti-Inflammatory: IL-1RA, TGFβ, IL-4, IL-6, and IL-10)
IF, interferon; IL, interleukin; PCR-RFLP, polymerase chain reaction with restriction fragment length polymorphism; PCR-SSP, polymerase chain reaction with sequence-specific primers; TGF, tissue growth factor; TNF, tumor necrosis factor.
Allele and genotype frequencies were calculated by the direct counting method, whereas deviations from Hardy-Weinberg proportion (HWP) and haplotype frequencies were estimated using a χ2 test (with one degree of freedom) and the Python for Population Genomics (PyPop vs. 0.7.0) software package, respectively (Lancaster et al, 2007). A principal component analysis (PCA) was conducted by Multivariate Statistical Software Package version 3.1 (Kovach, 1999) using our novel 22 cytokine SNP data from the Malays, Chinese, and Indians plus other reported data for Malay subethnic groups, including Orang Asli in Peninsular Malaysia, together with those for Asian, African, and European populations (Gonzalez-Galarza et al, 2011; Javor et al, 2007; Kaur et al, 2007; Norhalifah et al, 2015, 2016b; Skorpil et al, 2007; Sodsai et al, 2011; Trajkov et al, 2009).
Results
The cytokine SNP genotype data generated from this study have been deposited in the Allele Frequency Net Database (AFND) under the population name “Malaysia Peninsular Malay Cytokine,” “Malaysia Peninsular Chinese Cytokine,” and “Malaysia Peninsular Indian Cytokine” (their AFND identifiers are 3757, 3758, and 3759 respectively). Results for allele, haplotype, and HWP analyses are given in Tables 2-4 respectively.
Cytokine Gene Single Nucleotide Polymorphism Allele Frequencies in Malays, Chinese, and Indians in Peninsular Malaysia
Cytokine Gene Single Nucleotide Polymorphism Haplotype Frequencies in Malays, Chinese, and Indians in Peninsular Malaysia
Hardy-Weinberg Proportion Parameters in Malays, Chinese, and Indians in Peninsular Malaysia
Any values with p < 0.05 (indicated by *) were interpreted as evidence that the genotype frequencies do not fit HWE expectations.
Italic values indicate the allelic names.
Chi-squared test not calculated, because expected n was <5.
E, expected; HWE, Hardy-Weinberg equilibrium; O, observed; p, probability.
The majority of the screened cytokine gene SNPs are polymorphic in all three ethnicities. These include IL-1α −889 (T/C), IL-1β −511 (T/C), IL-1R pst1 1970 (T/C), and IL-1RA mspa1 11100 (T/C). Exceptions comprise SNPs such as TGF-β cdn 25 (G/C), IL-1β +3962 (T/C), and TNF-α −238 (A/G), which were all observed to be monomorphic in Malays, Chinese, and Indians (Table 2). Significant departure from HWP was observed for many cytokine gene loci across the three ethnic groups (e.g., IL-1RA mspa1 11100, IL-2 − 330, IL-6 − 174, IL-4 − 1098, and IL-10 − 1082; Table 4). In our PCA data mapping analysis, the cytokine gene SNP population data for Malays, Chinese, and Indians were found to lie in the left quadrant of the plot.
Discussion
Cytokine gene SNPs have previously been characterized and analyzed for various population groups, including Orang Asli groups in Peninsular Malaysia (Aznag et al, 2019; Gonzalez-Galarza et al, 2020; Javor et al, 2007; Kaur et al, 2007; Norhalifah et al, 2015; Sodsai et al, 2011). However, no cytokine gene SNP population data have ever been reported for the Malays, Chinese, and Indians living in Peninsular Malaysia.
In this study, many of the tested gene SNPs are polymorphic and deviated from HWP (Tables 2 and 4), which reflects the relevance of cytokines in disease association studies (Javalera et al, 2020; Norhalifah et al, 2018; Plataki et al, 2018). Although low sample size may contribute to the deviation from HWP, natural selection cannot be excluded as a driving force. This is a common phenomenon for immune-related genes such as human leukocyte antigen, killer cell immunoglobulin-like receptor, and major histocompatibility complex class I chain-related protein A, which are quite likely to be affected by location specific factors (Askar et al, 2013; Edinur et al, 2013; Nemat-Gorgani et al, 2014; Riccio et al, 2013).
No obvious relationship was observed between cytokine gene SNP population datasets from Malaysia and their respective putative ancestors in Africa, China, and India on PCA data mapping (Fig. 1). In the PCA plot, most of the population groups in Peninsular Malaysia including Orang Asli fall in the left quadrant and are well-separated from reference Asian, African, and European populations. Nonetheless, distinctions between ancestrally unrelated population groups in Malaysia can still be seen in the PCA plot, that is, Malays versus Orang Asli Semang (Bateq, Lanoh, and Kensiu) versus Orang Asli Senoi (Semai and Che Wong) versus Orang Asli Proto Malays (Orang Kanaq) versus Chinese versus Indians.
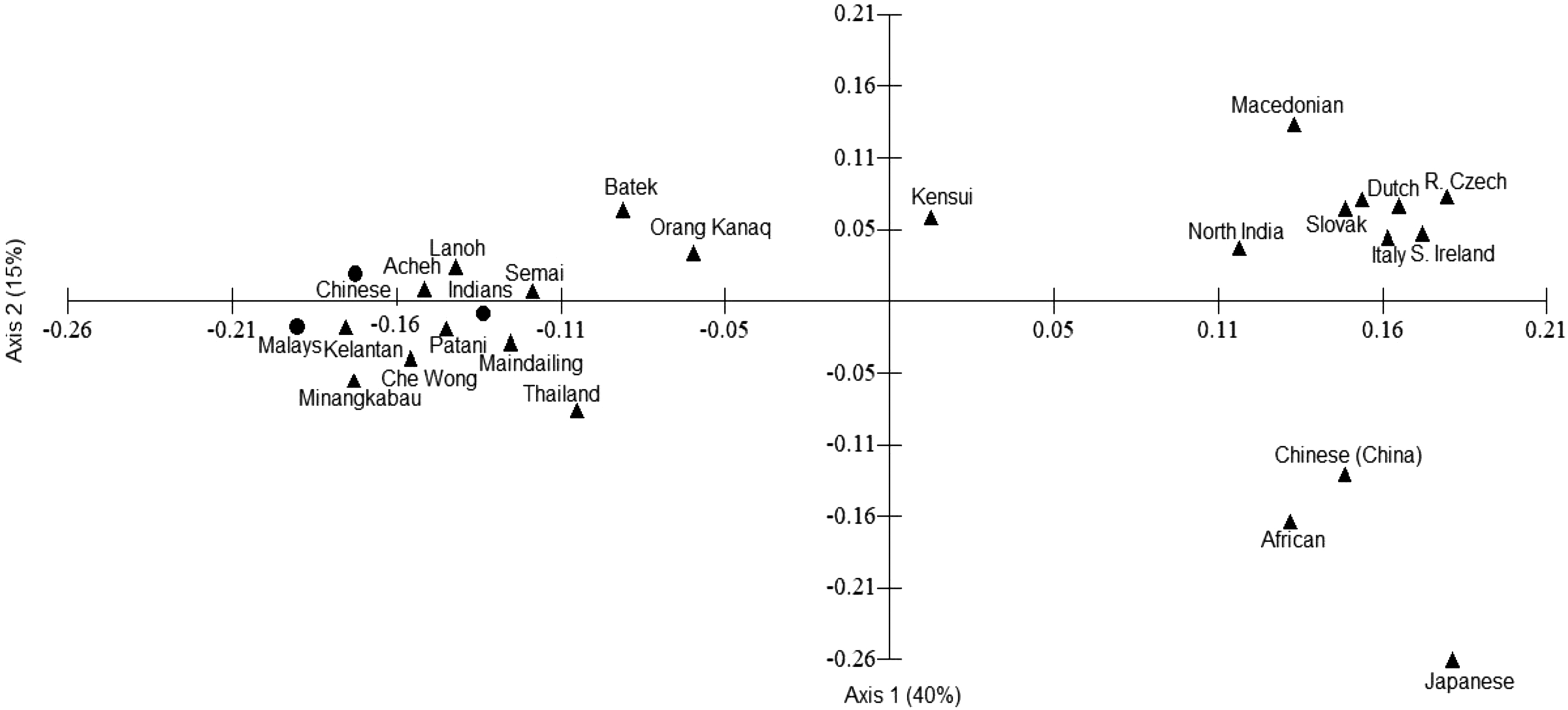
Principal coordinate analysis (PCA) constructed using cytokine data from Asian, African and European populations [Gonzalez-Galarza et al, 2011, Javor et al, 2007, Kaur et al, 2007, Norhalifah et al, 2015, Norhalifah et al, 2016b, Skorpil et al, 2007, Sodsai et al., 2011, Trajkov et al, 2008]. New population data from Malays, Chinese and Indians are indicated by • while reference populations are represented by a ▴ symbol.
In our opinion, local selective forces mentioned earlier plus admixture have all contributed to shaping the unique genepools of the various population groups in Peninsular Malaysia (Ahmad et al, 2021; Chambers and Edinur, 2021). Admixture, in particular, is reported to have occurred for more than hundred years and this may have resulted in significant gene flow between population groups in Peninsular Malaysia (Norhalifah et al, 2016a). Therefore, the newly generated cytokine gene SNP population datasets for Malays, Chinese, and Indians and our earlier cytokine data reported for Orang Asli and Malay ethnic groups (Norhalifah et al, 2015, 2016b) can serve as ethnically structured reference population datasets for future studies of ancestry and of diseases associated with cytokines.
Conclusions
Overall, we have successfully typed 22 SNPs in 13 cytokine genes in genetic material collected from unrelated and un-admixed Malay, Chinese, and Indian individuals in Peninsular Malaysia and have used the generated cytokine gene SNP datasets for an informative population-level study. Our analyses showed that various ancestral fractions in Peninsular Malaysia associated with multiple settlements in the region can still be observed, even though they may be partially obscured by local selection forces and admixture.
Footnotes
Acknowledgments
Geoff Chambers thanks Victoria University of Wellington, New Zealand for alumnus scholar support.
Authors' Contributions
N.H.C.G.: Conceptualization, writing—original draft, formal analysis, and writing—review and editing. S.M.A.: Conceptualization, methodology, writing—original draft, formal analysis, writing—review and editing, and funding acquisition. Z.Z., N.S.M.R., and T.H.T.M.: Resources-provision study materials. M.N.H., M.Y.A., and S.P.: Conceptualization, writing—review and editing. G.K.C.: Formal analysis, writing—review and editing. H.A.E.: Conceptualization, supervision, validation, writing—original draft, and writing—review and editing. N.F.C.M.: Conceptualization, methodology writing—original draft, formal analysis, validation, and writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Short Term Grant 304/PPSK/6315374 from the Universiti Sains Malaysia, Malaysia awarded to S.M.A.