
Abstract
Background:
There is increasing evidence that abnormal expression of microRNAs is involved in the occurrence and progression of tumors. In previous experiments, we found that the content of hsa-miR-1301-3p in tumor tissues of patients with nonsmall cell lung cancer (NSCLC) showed an obvious upward trend compared with that in normal tissues. We performed a detailed study on the impact and underlying mechanism of hsa-miR-1301-3p in NSCLC cells.
Methods:
The impact of hsa-miR-1301-3p on NSCLC cell proliferation, apoptosis, migration, and invasion was examined using colony formation, flow cytometry, modified Boyden chamber, and wound healing assays. Different doses of radiation were applied to NSCLC cells to investigate their sensitivity to radiotherapy. The potential target gene of hsa-miR-1301-3p was determined by dual-luciferase reporter assay and immunoblotting.
Result:
hsa-miR-1301-3p was upregulated in NSCLC tissues and cells. hsa-miR-1301-3p effectively promoted the rapid proliferation, migration, and invasion of NSCLC cells, while inhibiting apoptosis. It also induced radioresistance in NSCLC cells. hsa-miR-1301-3p targeted the homeodomain-only protein homeobox (HOPX) mRNA 3′ untranslated region and inhibited its transcription in NSCLC cells. Exogenous HOPX overexpression antagonized the mechanism by which hsa-miR-1301-3p regulates NSCLC cell proliferation, metastasis, and apoptosis.
Conclusions:
hsa-miR-1301-3p plays an oncogenic role in the occurrence and development of NSCLC. By targeting HOPX, hsa-miR-1301-3p can not only promote the proliferation and metastasis of NSCLC cells, but also alleviate apoptosis and reduce radiosensitivity.
Introduction
In recent decades, lung cancer has remained the top-ranked tumor in terms of mortality rate globally. Although surgery, chemotherapy, radiotherapy, targeted therapy, and immunotherapy have provided multiple means to treat lung cancer in recent years, its prognosis is still not optimistic. Clinically, >85% of lung cancer patients are diagnosed with nonsmall cell lung cancer (NSCLC) (Herbst et al., 2018; Siegel et al., 2020). Among these patients, >60% are already in the advanced stage at diagnosis, and 50% need radiation treatment.
Although there have been tremendous developments in radiotherapy technology in recent years, the effects of radiotherapy are still unsatisfactory. The overall prognosis of NSCLC is still poor, with a 5-year overall survival rate of 16% across all stages (Chen et al., 2016; Lazzari et al., 2017). The efficacy of radiotherapy depends on the radiosensitivity of tumor cells. It is very important to reveal novel mechanisms underlying radiosensitivity in NSCLC.
MicroRNAs (miRNAs) complement and pair with incomplete bases at the 3′ untranslated region (3′ UTR) of their target genes, resulting in mRNA degradation and restricted protein expression and production (Tutar et al., 2014). Dysregulation of miRNAs is associated with radiosensitivity in NSCLC (Gong et al., 2021; Xue et al., 2021). Previous studies have suggested that miRNAs can effectively regulate the transformation process of tumor cell differentiation and modulate the DNA damage response to radiation (Iorio and Croce, 2009; Oh et al., 2010).
According to the TargetScan (www.targetscan.org/vert_72/), miRDB (http://mirdb.org/), and OncomiR (www.oncomir.org/) prediction databases and online analysis, hsa-miR-1301-3p was identified to be associated with NSCLC development. Numerous studies have confirmed its close relationship with malignant biological behaviors of lung cancer (Xu et al., 2021), liver cancer (Liang et al., 2014; Yang et al., 2017), prostate cancer (Song et al., 2018; Wang et al., 2017), glioma (Bai et al., 2017; Zhi et al., 2017), breast cancer (Peng et al., 2018), osteosarcoma (Wang et al., 2018), and other tumors. However, there are no clear results on whether it has a significant response effect on radiosensitivity in NSCLC.
Homeodomain-only protein homeobox (HOPX) is a potential target gene of hsa-miR-1301-3p. Previous studies have indicated that HOPX plays important roles in embryonic development, tumor progression, immune regulation, and stem cell differentiation (Shin et al., 2002). HOPX was reported to be downregulated in malignant tissues, including head and neck (Yap et al., 2016), esophagus (Yamashita et al., 2008), lung (Cheung et al., 2013), breast (You et al., 2020), stomach (Ooki et al., 2010), pancreas (Waraya et al., 2012), colon/rectum (Katoh et al., 2012; Yamashita et al., 2013), placenta, and uterine malignant tumors (Yamaguchi et al., 2009).
HOPX overexpression inhibited tumor development (Chen et al., 2015; Mariotto et al., 2016). HOPX plays inhibitory roles in tumor progression and metastasis (Chen et al., 2015; Cheung et al., 2013). Through TCGA and cBioPortal (www.cbioportal.org/) databases, analyses revealed an inverse relationship between the protein content of HOPX in lung cancer patients and the expression of DNA repair genes in the body. We hypothesized that hsa-miR-1301-3p might affect the proliferation, migration, and other malignant biological behaviors of NSCLC through regulating HOPX expression, and that its expression levels could affect the radiosensitivity of NSCLC.
In this research, we identified the biological functions of hsa-miR-1301-3p in NSCLC cells, as well as its effects on the radiosensitivity of NSCLC cells. We also confirmed that hsa-miR-1301-3p targets HOPX in the radiosensitivity regulation process in NSCLC cells. Our studies provide a clue for enhancing the efficacy of radiotherapy in NSCLC treatment.
Materials and Methods
Cell culture
The NSCLC H1299 and H1975 cell lines and human normal lung epithelial cells (bronchial epithelium transformed with Ad12-SV40 2B [BEAS-2B]) were obtained from The Cell Bank of Type Culture Collection Center of Chinese Academy of Sciences. The cells were inoculated in RPMI-1640 medium (100 U/mL penicillin and streptomycin) containing 10% fetal bovine serum (HyClone, Logan, UT) and cultured in a 5% CO2, 37°C incubator.
Ethical approval
The trial was conducted in accordance with the Declaration of Helsinki. The study was approved by the scientific research ethics committee of Zhongnan Hospital of Wuhan University.
Modified Boyden chamber assays
Transwells (Becton-Dickinson Ltd.) were mainly used to test the effects of the hsa-miR-1301-3p complex on cell proliferation and metastasis. We diluted the cells in the logarithmic growth phase in a petri dish, and then placed them into each reaction chamber at a density of 5 × 104 cells per 200 μL. The chambers were coated with Matrigel for the invasion assay. After a reaction time of 48 h, 4% formaldehyde was added for 20 min to fix the cells. The cells were placed under a microscope for viewing and testing (200 × ).
Wound healing assays
Cells were seeded and grown in six-well plates until they reached 90% confluence. A scratch was made through each well using a sterile pipette tip. The plates were then washed twice with PBS. The process of wound healing was monitored for 48 h, and images were captured at the time point of 0, 24, and 48 h.
Quantitative real-time polymerase chain reaction
RNA was extracted with TRIzol reaction reagent, and then the RNA was reverse transcribed into cDNA; the results were quantitatively tested by polymerase chain reaction (qPCR). Primer sequences of hsa-miR-1301-3p and U6 (internal reference) are as follows: hsa-miR-1301-3p: 5′-ACACTCCAGCTGGGTTGCAGCTGCCTGGGAGTGA (forward), 5′-CTCAACTGGTGTCGTGGA (reverse), and 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGAAGTCAC (RT primer); U6 (internal reference): 5′-CTCGCTTCGGCAGCACA (forward) and 5′-AACGCTTCACGAATTTGCGT (reverse); HOPX: 5′-TCAACAAGGTCGACAAGCAC (forward) and 5′-AGGCGCTGCTTAAACCATT (reverse); and GAPDH (internal reference) are 5′-GACAAGCTTCCCGTTCTCAG (forward) and 5′-GAGTCAACGGATTTGGTGGT (reverse).
These primers were synthesized by Wuhan Tianyihuiyuan Biotechnology. The 2−ΔΔCT method was used to determine the transcription rate.
Immunoblotting
Total protein was isolated from cultured cells with RIPA lysis buffer supplemented with protease inhibitor (Beyotime, China), and the protein concentration was determined using the BCA quantification kit (Beyotime, China). Equal amounts of proteins were separated on sodium dodecyl sulfate polyacrylamide gel electrophoresis gels and transferred to polyvinylidene difluoride membranes. Then, the membranes were blocked with 5% nonfat milk. After 2 h, the membranes and the primary antibody were stored overnight at a constant temperature of 4°C. The membranes were then incubated with some horseradish peroxidase-conjugated antirabbit antibodies (SA00001-2; Proteintech) or antimouse antibodies (SA00001-1; Proteintech) for 1.5 h.
The Image J program was used to quantify the protein levels. The primary antibodies were as follows (all from Proteintech): anti-HOPX (11419-1-AP), anti-E-cadherin (20874-1-AP), anti-Vimentin (10366-1-AP), anti-Bcl-2 (60178-1-IG), anti-Bax (60267-1-IG), anti-Caspase-3 (cleaved, 66470-2-IG), and anti-GAPDH (10491-1-AP).
Dual-luciferase reporter gene assay
All samples were from Guangzhou Ruibo Biological Company, including hsa-miR-1301-3p mimic (miR10005797), mimic negative control (miR1N0000001-1-5), hsa-miR-1301-3p inhibitor (miR20005797), and inhibitor negative control (miR2N0000001-1-5). The wild-type (WT) and mutant (MUT) gene sequences in the HOPX 3′ UTR sequence were replicated into the pGL-3M composite recombinant plasmid. HEK-293 human embryonic kidney cells were cotransfected with hsa-miR-1301-3p mimics or negative control, and pGL-3M 3′ UTR WT or pGL-3M 3′ UTR MUT. Afterward, the degree of cellular response to luciferase was measured by a dual-luciferase detection system.
HOPX overexpression and knockdown
Human HOPX (NM_001145459.1)-overexpressing M02 plasmid was purchased from Wuhan Junengyitong Biology. HOPX siRNA was purchased from Suzhou GenePharma. To avoid off-target effects, two different HOPX siRNA pairs were selected for the experiment: HOPX-Homo-994: 5′-GAAGCUAUGUGUAUCUUCUTT (sense) and 5′-AGAAGAUACACAUAGCUUCTT (antisense); HOPX-Homo-1220: 5′-CCCUAAGUCACUUUCCUUATT (sense) and 5′-UAAGGAAAGUGACUUAGGGTT (antisense).
Statistical analysis
All experiments were carried out three times, and then the data results were put into SPSS 17.0 software for analysis. All data were tested using the mean value. In the analysis, the single factor effect was mainly used for the variance test (ANOVA). Data were considered statistically significant only when p < 0.05 was calculated. Statistical graphs were made with Graphpad Prism 7 software.
Results
hsa-miR-1301-3p promotes the proliferation and migration of NSCLC cells
We analyzed the RNA sequencing data of lung cancer samples in the TCGA database to explore the roles of hsa-miR-1301-3p in lung cancer. The content of hsa-miR-1301-3p in the tumor tissues was much higher than that in the normal tissues (Fig. 1A). The qPCR results showed that the expression levels of hsa-miR-1301-3p were higher in NSCLC (H1299 and H1975) cells than in BEAS-2B cells (Fig. 1B). The results suggested that hsa-miR-1301-3p upregulation was involved in the occurrence and development of lung cancer, implying that hsa-miR-1301-3p might play a procancer role in NSCLC.

hsa-miR-1301-3p was overexpressed in lung cancer tissues and cells.
To verify the biological activity of hsa-miR-1301-3p in NSCLC cells, H1299 and H1975 cells were treated with hsa-miR-1301-3p mimics or inhibitors, whose effects were verified by qPCR (Fig. 1C). Colony formation and CCK-8 assays indicated that hsa-miR-1301-3p mimics could effectively promote the proliferation of NSCLC cells, whereas its inhibitors showed opposite effects (Fig. 2A-C). The results of flow cytometry indicated that hsa-miR-1301-3p mimics significantly inhibited H1299 and H1975 cell apoptosis, whereas its inhibitors promoted NSCLC cell apoptosis (Fig. 2D, E).
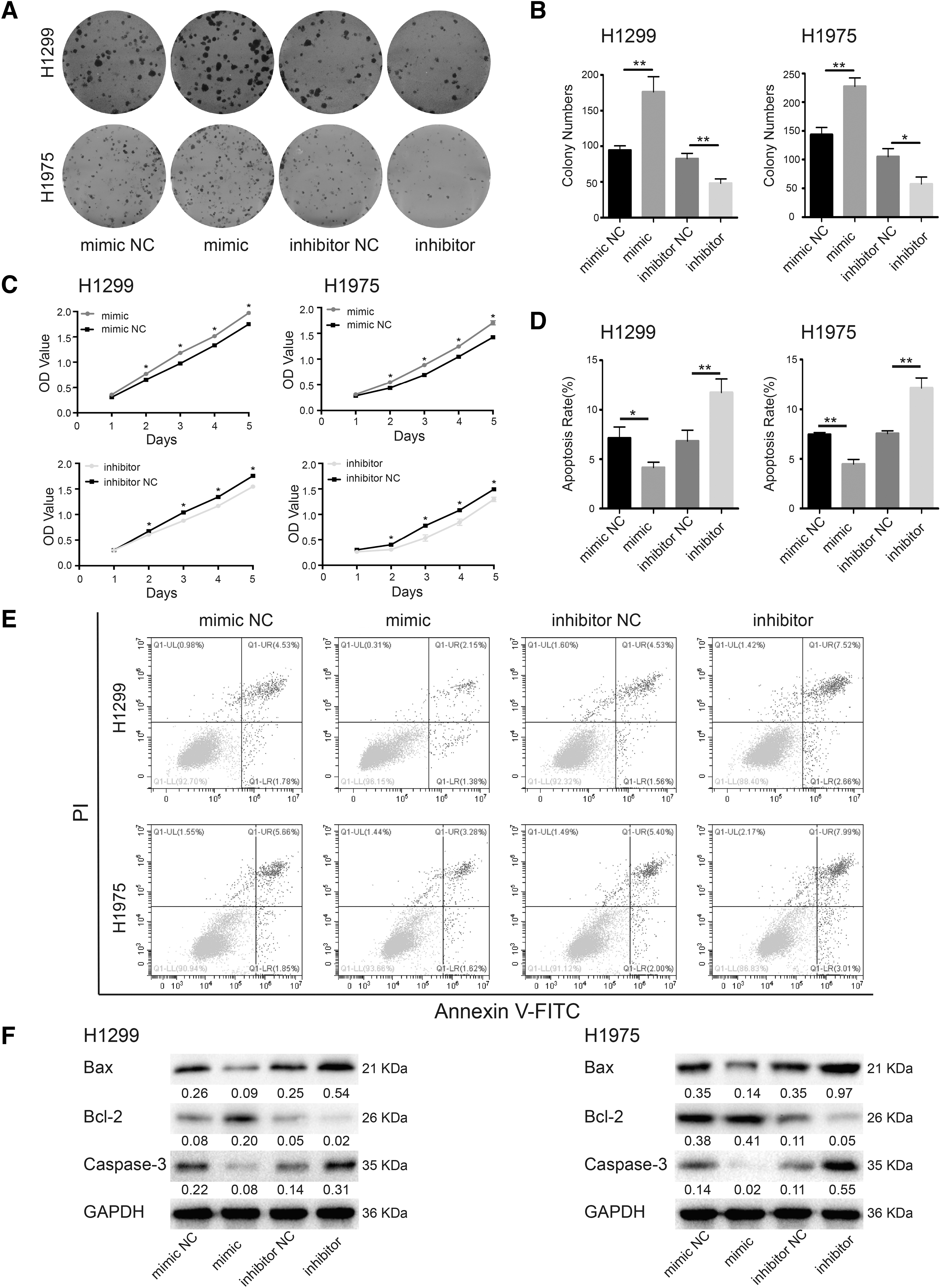
hsa-miR-1301-3p induced NSCLC cell proliferation and reduced apoptosis.
Immunoblotting results showed decreased Bax and Caspase-3 protein levels and increased Bcl-2 levels upon hsa-miR-1301-3p overexpression, whereas its inhibition induced the opposite effects (Fig. 2F). These findings indicated that hsa-miR-1301-3p could indeed induce NSCLC cells proliferation and suppress apoptosis. Modified Boyden chamber and wound healing assays showed that treatment of NSCLC cells with hsa-miR-1301-3p mimics significantly promoted metastasis and invasion compared with that in the negative control group, whereas hsa-miR-1301-3p inhibitors significantly reduced NSCLC cell migration and invasion (p < 0.05, Fig. 3A, B).

hsa-miR-1301-3p promoted invasion and migration of NSCLC cells.
Immunoblotting results showed decreased E-cadherin protein levels and increased Vimentin levels upon hsa-miR-1301-3p overexpression, whereas its inhibition displayed the opposite effects (Fig. 3C). These findings indicated that hsa-miR-1301-3p could induce the migration and invasion of NSCLC cells.
hsa-miR-1301-3p enhances the radiotherapy resistance of NSCLC cells
Radiation eliminates tumor cells mainly by inducing DNA damage. To improve the efficacy of radiotherapy in NSCLC, it is important to reveal the mechanism of radiation tolerance. To explore whether hsa-miR-1301-3p could affect the radiosensitivity of NSCLC cells, H1299 and H1975 cells treated with hsa-miR-1301-3p mimics were subjected to different doses of radiation. The radiosensitivity of H1299 and H1975 cells treated with mimics was significantly reduced compared with that in the control cells, whereas inhibitor treatment induced opposite effects (p < 0.05, Fig. 4). The above results indicate that hsa-miR-1301-3p mimics could increase the radiotherapy resistance of NSCLC cells.
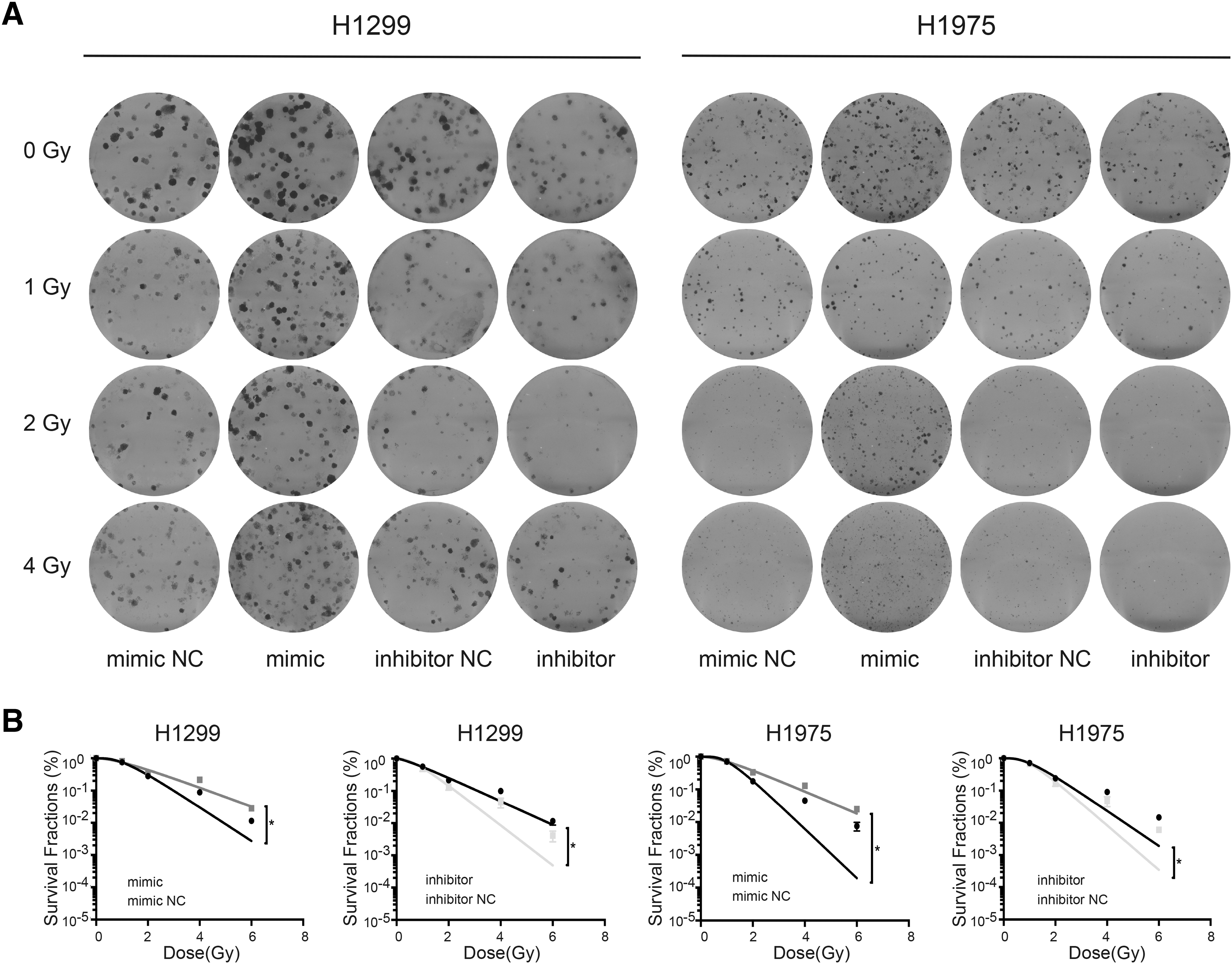
hsa-miR-1301-3p downregulated radiosensitivity of NSCLC cells.
hsa-miR-1301-3p targets the HOPX mRNA 3′ UTR and inhibits its expression
To explore the target genes of hsa-miR-1301-3p in NSCLC, we used the online analysis software Targetscan to conduct analysis of biological interactions in the samples. From the results, HOPX was indeed predicted to be a target gene of hsa-miR-1301-3p (Fig. 5A). To further confirm this targeted regulation, dual-luciferase reporter gene carriers containing HOPX 3′ UTR binding sites and corresponding mutation sites were established, and the genes were assessed by dual fluorescence assays. Luciferase activity was significantly reduced after cotransfection of hsa-miR-1301-3p mimics and dual luciferase reporter vectors containing WT binding sites of hsa-miR-1301-3p at the HOPX 3′ UTR.
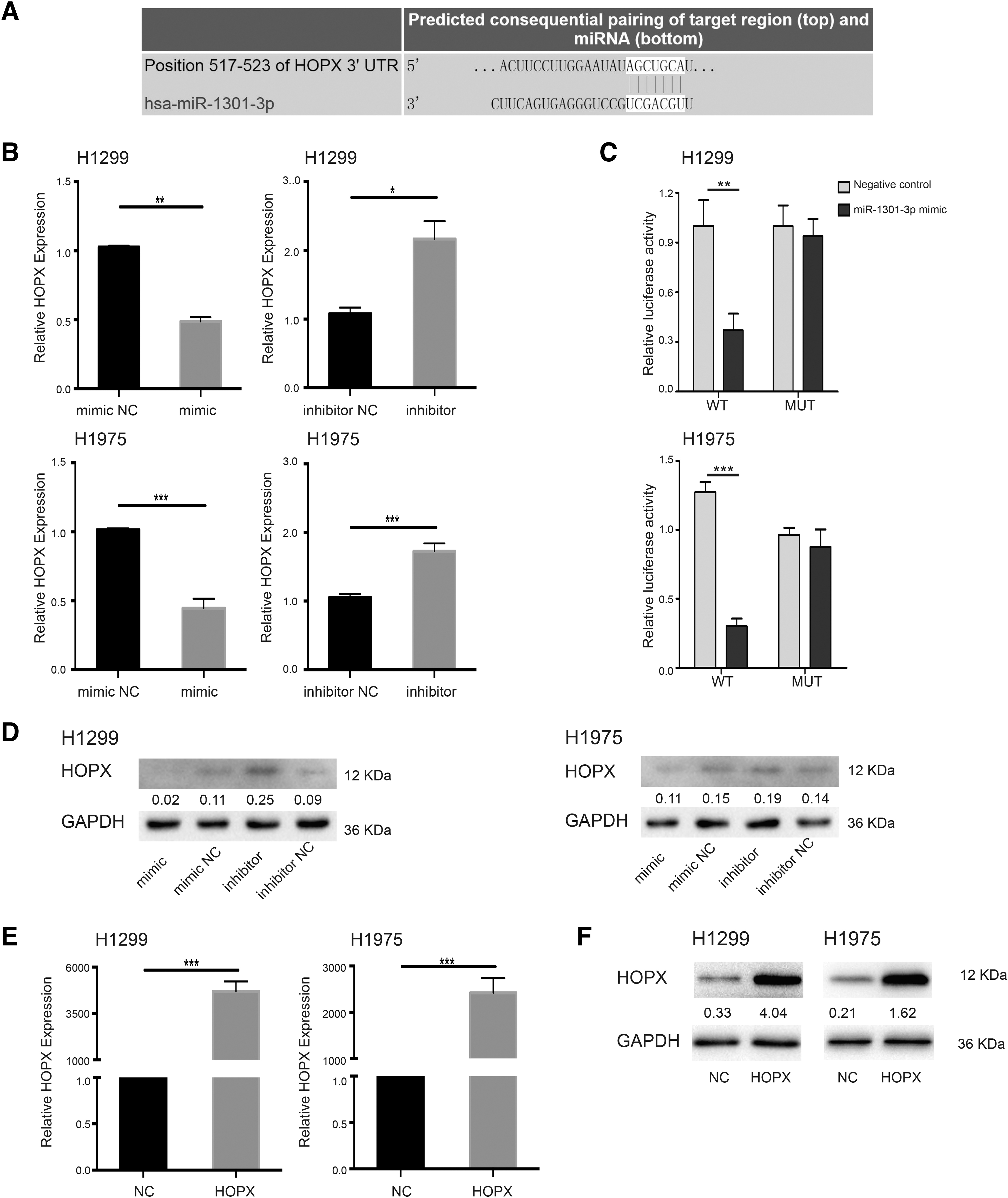
hsa-miR-1301-3p targeted HOPX.
However, luciferase activity was not affected by cotransfection of hsa-miR-1301-3p mimics and reporter vectors containing mutant binding sites of hsa-miR-1301-3p at the HOPX 3′ UTR (Fig. 5B). The results showed that compared with the negative control group, H1299 and H1975 cells were treated with the hsa-miR-1301-3p mimics, and it was found that the mRNA content and protein composition of HOPX in the cells were significantly decreased. At the same time, HOPX was significantly upregulated in H1299 and H1975 cells transfected with hsa-miR-1301-3p inhibitors (Fig. 5C, D). These results indicated that hsa-miR-1301-3p negatively regulates HOPX expression in NSCLC cells.
hsa-miR-1301-3p promotes the proliferation and migration of NSCLC cells by targeting HOPX
Radiation causes a variety of types of DNA damage, and whether tumor cells in vivo have significant radiation sensitivity depends on DNA repair capability (Marta et al., 2015). The genes involved in DNA damage repair and related to radiosensitivity include ATM, ATR, BRCA1, HIPK2, RAD51, RAD52, CHK1, γ-H2AX, XRCC1, and XRCC3 (Choudhury et al., 2006). However, bioinformatics analysis showed that hsa-miR-1301-3p did not target any of the abovementioned genes involved in DNA damage repair.
After analyzing the data of the TCGA and cBioPortal databases, the profiling results revealed that the expression of HOPX in the cells of lung cancer patients was negatively correlated with that of the above important genes (Supplementary Fig. S1). We speculated that hsa-miR-1301-3p might upregulate DNA damage repair genes by targeting HOPX, thus improving the tolerance of NSCLC cells to radiotherapy.
To verify this hypothesis, HOPX was overexpressed in H1299 and H1975 cells and verified by qPCR and immunoblotting (Fig. 5E, F). Colony formation and proliferation assays showed that overexpression of HOPX resulted in reversal of the induction of NSCLC cell growth by hsa-miR-1301-3p (Fig. 6A-C). The flow cytometry results indicated that HOPX overexpression antagonized the inhibitory effects of hsa-miR-1301-3p on NSCLC cell apoptosis (Fig. 6D, E). Immunoblotting also showed that HOPX overexpression inverted the mechanisms of has-miR-1301-3p on apoptosis-related Bax, Bcl-2, and Caspase-3 protein levels (Fig. 6F). These results suggest that hsa-miR-1301-3p regulates NSCLC behaviors by targeting HOPX.
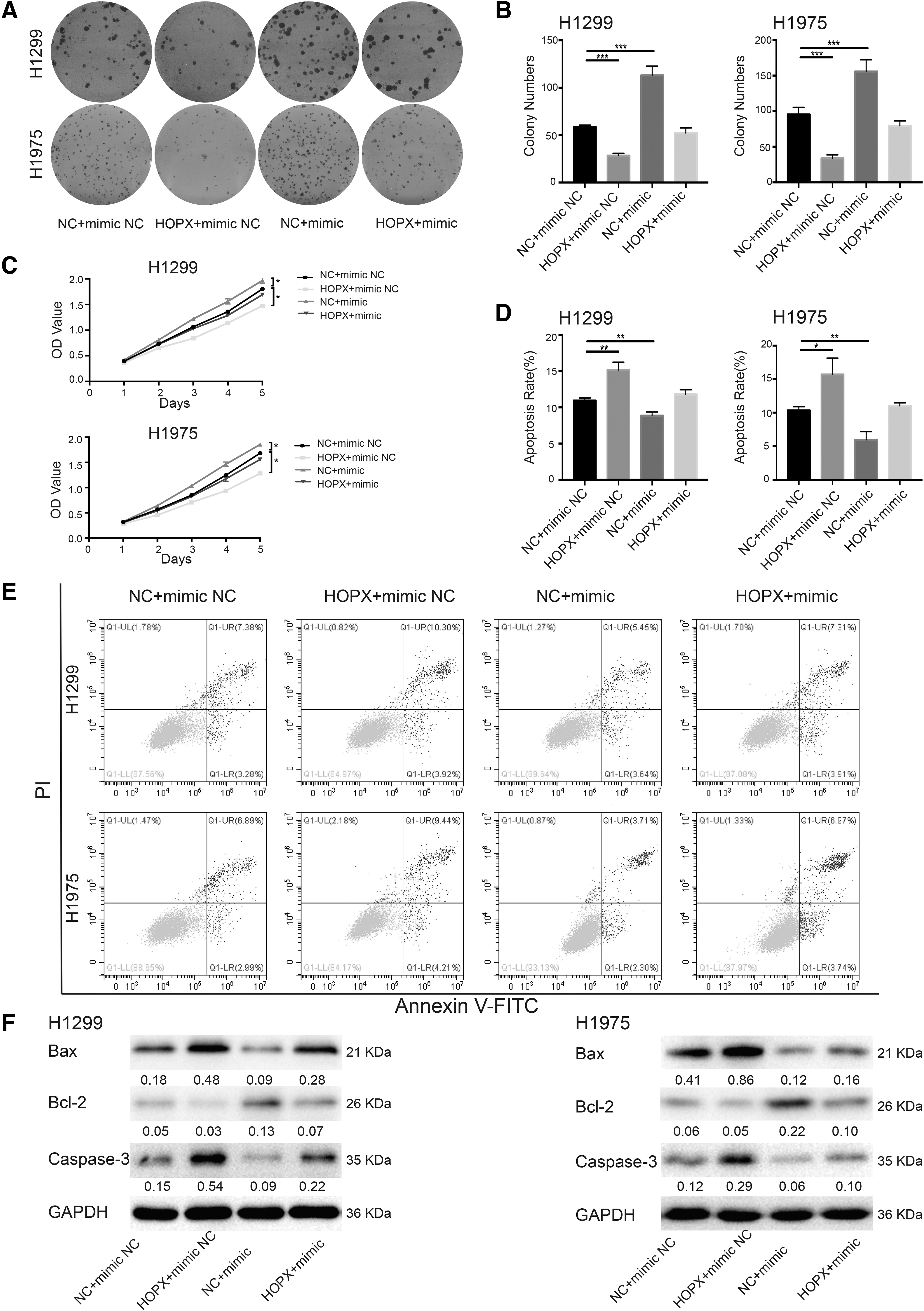
HOPX overexpression reversed the effects of hsa-miR-1301-3p on NSCLC cell proliferation and apoptosis.
Modified Boyden chamber and wound healing assays showed that HOPX overexpression antagonized the induction effect of hsa-miR-1301-3p mimics on the metastasis and proliferation of NSCLC cells (Fig. 7A, B). Immunoblotting also showed that HOPX overexpression reversed the effects of hsa-miR-1301-3p on E-cadherin and Vimentin protein levels (Fig. 7C). From these results, it can be inferred that hsa-miR-1301-3p promoted NSCLC cell migration and invasion by targeting HOPX.
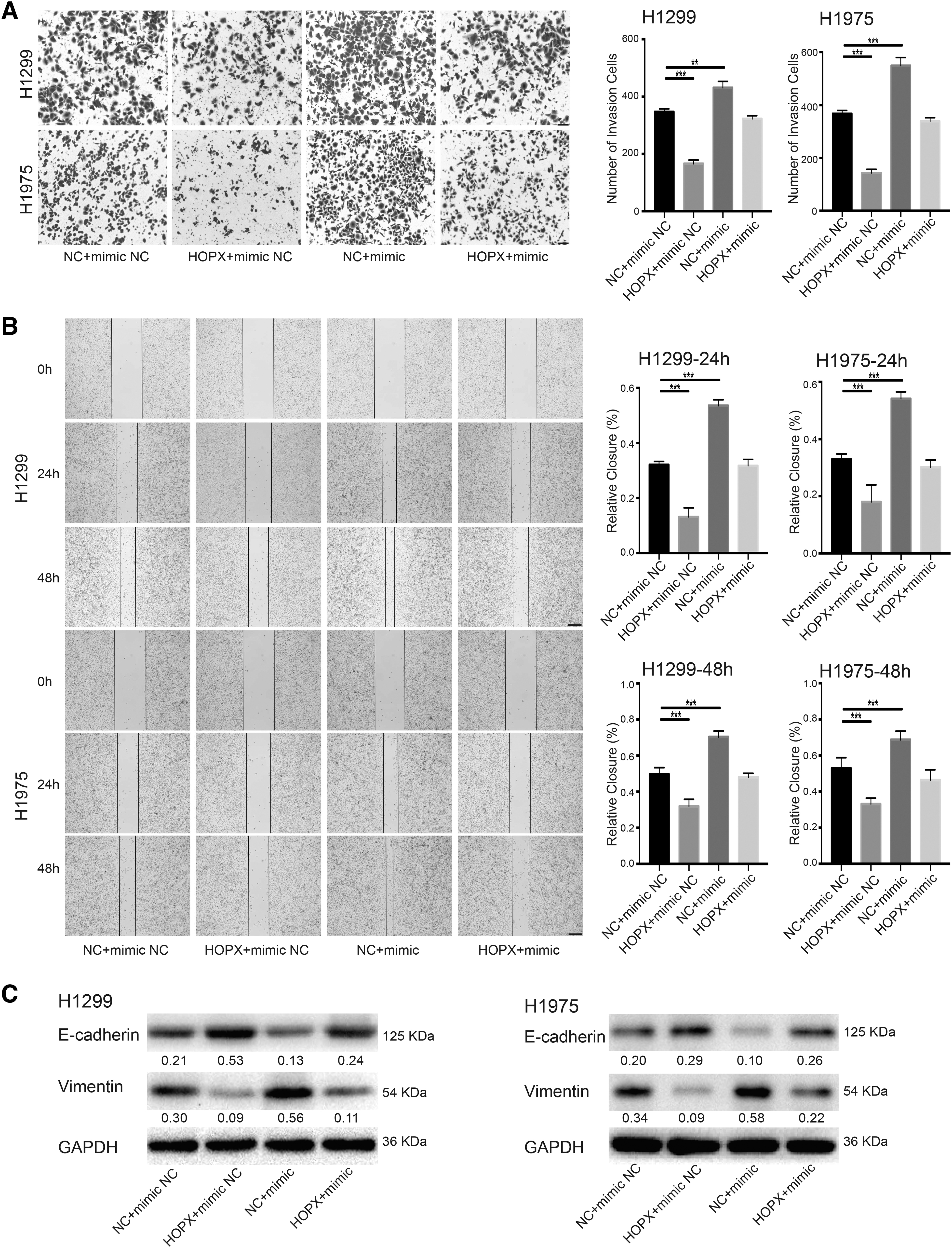
HOPX overexpression antagonized the effect of hsa-miR-1301-3p mimics on cell migration and invasion.
HOPX is downregulated in NSCLC as a tumor suppressor
Our previous results indicated that HOPX was the target gene of hsa-miR-1301-3p. The GEPIA database (http://kmplot.com/analysis/index.php?P=Service&Cancer=Lung) analysis showed that the median expression levels of HOPX in lung cancer tissues were significantly lower than those in normal tissues (Supplementary Fig. S2A). Survival analysis showed that high HOPX expression was associated with a better prognosis than low expression (p < 0.05, Supplementary Fig. S2B).
These results preliminarily predicted the antitumor effects of HOPX. In this study, qPCR results showed that HOPX expression in H1299 and H1975 cells was significantly lower than that in BEAS-2B cells (Fig. 8A). Immunohistochemical results of tissue samples from NSCLC patients also showed that HOPX expression in NSCLC tissues was lower than that in adjacent normal tissues (Fig. 8B). These results suggested that HOPX might be a tumor suppressor in NSCLC.

HOPX was downregulated in NSCLC cells and tissues.
Upon irradiation at a dose of 2 Gy, the protein levels of γ-H2AX were increased in the HOPX overexpression group, whereas HOPX knockdown suppressed γ-H2AX expression in H1299 cells (Fig. 8C). Moreover, the levels of the DNA damage repair proteins RAD-51 and BRCA-1 were measured, and the results showed that overexpression of HOPX led to decreased RAD-51 and BRCA-1 expression, whereas knockdown of HOPX yielded the opposite results (Fig. 8C). The downregulation of HOPX might promote NSCLC cell proliferation, invasion, and migration; inhibit cell apoptosis; and reduce the sensitivity of NSCLC cells to radiotherapy.
Discussion
Because of the high rate of metastasis and recurrence, the clinical outcome of NSCLC is still unsatisfactory (Goldstraw et al., 2016). Some dysregulated miRNAs have been found to be related to tumor growth, metastasis, or radiosensitivity of various tumors, including NSCLC (Gong et al., 2021; Xue et al., 2021; Zheng et al., 2017). The combined use of miR-492, miR-590-3p, and miR-631 was considered as a promising biomarker for NSCLC patients (Duan et al., 2021). Dysregulated expression of miR-133b, miR-124-3p, miR-145-5p, and miR-155-5p in NSCLC leads to adverse outcomes (Zhang et al., 2016).
MiR-9 (Wei et al., 2019), miR-519a (Gong et al., 2021), and miR-219a-5p (Wei et al., 2020) were reported to enhance the radiosensitivity of NSCLC, whereas miR-182 (Chen et al., 2019) and miR-210 (Grosso et al., 2013) had the opposite effects. Recent studies have shown that the abnormal expression of hsa-miR-1301-3p is closely related to tumor initiation. Previous studies have demonstrated that the content of hsa-miR-1301-3p is significantly increased in prostate cancer cells of patients. This results in a rapid expansion of stem cells and a rapid increase in the expression of genes associated with stem cell proliferation (Song et al., 2018).
In breast cancer, hsa-miR-1301-3p may induce cell stasis, thereby effectively inhibiting cell proliferation (Peng et al., 2018). In thyroid papillary carcinoma, the expression of hsa-miR-1301-3p was significantly decreased, which was related to T and N grade. Upregulation of hsa-miR-1301-3p inhibited cell proliferation and migration (Qiao et al., 2021). Moreover, hsa-miR-1301-3p expression can increase TNM stage, and lymph node metastasis is a factor that needs to be considered in the treatment of NSCLC (Xu et al., 2021). However, there is no report on the radiosensitizing roles and potential mechanisms of hsa-miR-1301-3p in NSCLC.
This study found that the expression of hsa-miR-1301-3p was significantly increased in the lung cancer tissues. hsa-miR-1301-3p promoted NSCLC cell proliferation, migration, and invasion, and inhibited cell apoptosis. More importantly, hsa-miR-1301-3p reduced the radiosensitivity of NSCLC cells at different doses.
In this study, we predicted the potential targets of hsa-miR-1301-3p, and conducted a detailed analysis of HOPX. HOPX was initially identified as a gene essential for cardiac growth and development. Recent findings suggested that HOPX acted as a tumor suppressor in various cancers. HOPX plays critical roles in epithelial cell homeostasis and serves as a tumor suppressor in head and neck cancer (Yap et al., 2016). In breast cancer, HOPX is epigenetically silenced, and its overexpression suppresses tumor progression (You et al., 2020).
HOPX was also reported as a tumor suppressor critical for tumor aggressiveness in pancreatic carcinogenesis (Waraya et al., 2012). HOPX and GATA6, as inhibitors of metastatic progression, cooperatively suppress the metastatic potential of adenocarcinoma cells (Cheung et al., 2013). Recent studies also found that HOPX overexpression inhibited the malignant phenotypes of NSCLC (Liang et al., 2019).
A dual-luciferase assay showed that hsa-miR-1301-3p could bind to the 3′ UTR of HOPX to regulate its expression. Through analysis of coexpression profiles of TCGA and cBioPortal databases, HOPX expression in lung cancer cells was found to be inversely related to the expression of DNA damage repair-related genes, including ATM, ATR, BRCA1, HIPK2, RAD51, and RAD52. HOPX downregulation by hsa-miR-1301-3p might upregulate these important genes, enhance DNA damage repair in lung cancer cells, and reduce the radiosensitivity of NSCLC cells.
HOPX overexpression antagonized the effects of hsa-miR-1301-3p on NSCLC cell proliferation, metastasis, and apoptosis. Therefore, our results suggest that hsa-miR-1301-3p might upregulate DNA damage repair genes by targeting HOPX, thus improving the tolerance of NSCLC cells to radiotherapy. These results provide novel targets and research ideas for the molecular diagnosis and radiosensitivity evaluation of NSCLC.
In conclusion, by targeting HOPX, hsa-miR-1301-3p promotes the proliferation, invasion, and migration of NSCLC cells; inhibits apoptosis; and reduces radiosensitivity. Our studies on the hsa-miR-1301-3p/HOPX axis may contribute to the research progress on overcoming the tolerance of NSCLC cells to radiotherapy and revealing the tolerance mechanism.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by National Natural Science Foundation of China (81972852 and 81800429), Key Research and Development Project of Hubei Province (2020BCA069), Health Commission of Hubei Province Medical Leading Talent Project, Young and Middle-Aged Medical Backbone Talents of Wuhan (WHQG201902), Application Foundation Frontier Project of Wuhan (2020020601012221), Translational Medicine and Interdisciplinary Research Joint Fund of Zhongnan Hospital of Wuhan University (ZNJC201922 and ZNJC202007), and Chinese Society of Clinical Oncology TopAlliance Tumor Immune Research Fund (Y-JS2019-036).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.