
Abstract
Aims:
Evaluating the association between a single nucleotide polymorphism in the 3′ untranslated region (3′UTR) of the miRNA binding site of the NLRP3 gene and the occurrence and development of chronic obstructive pulmonary disease (COPD) and providing information to aid in the early detection and treatment of COPD.
Materials and Methods:
The regulatory single nuclear polymorphisms (SNPs) located in NLRP3 3′UTR were searched by using the dbSNP database and miRNA binding site prediction database. Meanwhile, samples from COPD patients and healthy controls in the same period were used for verification. The clinical baseline information of all subjects was collected, and the transcription level and protein expression level of NLRP3 and the expression level of inflammatory factors downstream of NLRP3 were detected. The effects of SNPs’ single nucleotide changes on the transcription and expression of inflammatory factors were analyzed.
Results:
The study included 418 participants (249 in the COPD group and 169 in the control group). NLRP3 SNPs with miRNA binding sites include rs10754558 (G > C), rs1664774076 (ATAT > del), and rs1664775106 (C > G). Furthermore, two genotypes, GCG and GCA, were discovered to have a linkage mutation at 3′UTR 459-461. COPD susceptibility is tightly associated with the expression of the rs1664774076 del/del genotype, and the risk of COPD increased by 2.770 times (p = 0.003). Type 459-461 GCA was substantially related to the likelihood of developing COPD at various stages (p < 0.05). Except for rs10754558, all homozygous mutants increased NLRP3 mRNA and protein levels. NLRP3 had the greatest area under the receiver operating characteristic (ROC) curve for predicting the development and diagnosis of COPD when compared with its downstream inflammatory variables (AUC = 0.9291).
Conclusions:
The NLRP3 rs1664774076 del/del genotype is a COPD susceptibility gene, and the GCA genotype at 459-461 can be used as an early predictor of COPD exacerbation. The NLRP3 3′UTR polymorphism may alter the loss of miRNA binding sites, leading to an increase in NLRP3 expression. In the development of COPD, NLRP3 has a better diagnostic value than traditional inflammatory factors.
The Clinical Trials Registration number Z: protocol KY01-2020-11-06.
Introduction
Chronic obstructive pulmonary disease (COPD) is one of the diseases with the highest morbidity and mortality rates worldwide, and the global COPD prevalence rate is on the rise, reaching 9-10% among persons older than 40 (Vijayan, 2013). COPD is a lung disease characterized by incomplete reversibility and continuous progressive airflow limitation, with chronic inflammation and structural deterioration of the airway wall and lung parenchyma as its pathological basis. At present, it has been established that the cause of COPD is the inflammatory reaction of lung tissue, and that long-term stimulation of lung tissue by smoke and pollution is the inducer (Vogelmeier et al., 2020). Despite the fact that smoking is a major cause of COPD, between 25% and 45% of COPD patients have never smoked, and these patients have a tendency toward regional and familial clustering, suggesting that the onset of COPD is heritable (Ortega-Martinez et al., 2020). Genetic variation analysis may be useful for screening the COPD-vulnerable population. In terms of etiology, the inflammatory response plays a crucial indirect or direct role in the process of lung injury, which is a major cause of decreased lung tissue elasticity, retraction disorder, excessive alveolar expansion, and progressive fibrosis. Mutation of inflammatory factors may result in alterations to the inflammatory response. Therefore, we attempt to construct the inflammatory response based on the molecular genetic mechanism and investigate its relationship with the occurrence and progression of COPD.
Nucleotide-binding oligomerization domain-like receptor thermal protein domain associated protein 3 (NLRP 3) regulates the occurrence and development of chronic inflammation in COPD as a key factor of inflammatory corpuscles (Guo et al., 2022). Faner et al. described that NLRP3 inflammatory bodies lead to focal death and passive release of proinflammatory cytokines, including mature IL-1β and IL-18, which trigger inflammatory response in COPD patients, and that NLRP3 inflammatory bodies are overexpressed during the deterioration period, and IL-1β and IL-18 family cytokines are released into the surrounding tissues (Chen et al., 2019). Inhibiting the production of NLRP3 inflammatory corpuscles can therefore indirectly block the inflammatory effects of IL-1β and IL-18, making it an ideal therapeutic target for COPD.
Gene mutations in the core domain of the NLRP3 gene play a crucial role in the regulation of the immune inflammatory response, resulting in genetic susceptibility to a variety of acute and chronic inflammatory diseases (Lu et al., 2021). In contemporary molecular epidemiology, single nuclear polymorphism (SNP) is frequently used as a biomarker of disease susceptibility. One of the mechanisms by which SNPs affect disease susceptibility is by modulating the interaction between miRNA and mRNA to modulate gene expression. MiRNA (microRNAs) can precisely bind to the 3′ untranslated region (3′UTR) sequence of the target gene mRNA to inhibit translation or promote degradation of the target gene at the posttranscriptional level. Therefore, this study hypothesizes that the genetic variation of the NLRP3 3′UTR influences the binding of miRNA-mRNA, resulting in a change in the NLRP3 protein level and thereby influencing the occurrence or development of COPD.
In this study, we used the prediction website of miRNA target genes and their binding sites to identify meaningful 3'UTR NLRP3 SNP, and investigated the relationship between these SNPs and COPD susceptibility or progression to provide a new perspective for the exploration of its pathogenesis or potential therapeutic targets from the research of COPD genomics, and to identify genetic risk factors that can contribute to the early detection, early diagnosis, and prevention of COPD.
Materials and Methods
Study population
All of the subjects are from the Fifth Affiliated Hospital of GuangZhou Medical University and have received approval from the institution’s Ethics Review Committee. Inclusion and exclusion criteria were as follows:
COPD was chosen based on the diagnostic guidelines for COPD (Rabe et al., 2007). Exclude patients with additional chronic respiratory conditions, including lung cancer, thyroid cancer, and other malignant tumors, those having significant immune deficiency and immune system diseases, antibiotics being administered within 30 days before selection, and those with local or systemic infection. In addition, according to the Global Initiative for Chronic Obstructive Lung Disease (GOLD) standard, the COPD cohort patients were divided into the following categories: GOLD1, mild patients: FEV1%pred ≥70%. GOLD2, moderate patients: 50%≤FEV1%pred < 70%. GOLD3, severe patients: 30%≤FEV1%pred < 50%. GOLD4, extremely severe patients: FEV1%pred < 30%. Standard of healthy control group: healthy people who showed no abnormalities in the lungs by CT scan, from the physical examination center in the same period. Exclude healthy people with chronic respiratory disease, cough, and infection.
Screening of miRNA targeting 3′UTR of NLRP3 gene SNPs
The sequence of 3′UTR in NLRP3 was obtained by using the UCSC database (http://genome.ucsc.edu/), and the transcript ID was ENST_0000391828.3, and the matching NCBI blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was NG_007509.2, which was a gene fragment of NLRP3. All SNP sites of NG_007509.2 were screened from the NCBI SNP, that is, all SNPs of NLRP3 3′UTR. MiRNA binding sites of NLRP3 were obtained by using the miRNA prediction websites miRDB, miRTarBase, DIANA, miRcode, and TargetScan. Under bioinformatic simulation, the sequence of miRNA was aligned with the position of the 3'UTR SNP site, and the SNP site with the ability to regulate gene expression was identified.
Amplification and sequencing of 3′UTR of NLRP3
All participants in this study underwent NLRP3 3′UTR gene sequencing. After 12 h of fasting, 2 mL of ethylene diamine tetraacetie acid (EDTA) anticoagulant was extracted the following morning, followed by the extraction of whole blood DNA (TIANGEN blood DNA extraction Kit). The target fragment was amplified using PCR (TIANGEN 2 Taq PCR Master Mix II PCR Kit, primer shown in Table 1), and the sequence of the NLRP3 3′UTR was determined using the Sanger sequencing technique. ClustalX is used to compare the mutation situation of the population, and sites with a mutation rate greater than 30%, which are in agreement with the predicted SNP sites with regulatory function, are selected for inclusion in the study.
Primer Sequences for PCR and qRT-PCR
Detection of mRNA expression and downstream factor level
After fasting for 12 h, all subjects took 2 mL of EDTA anticoagulant and 5 mL of EDTA procoagulant in the morning of the next day. EDTA anticoagulation is used to extract total RNA from whole blood (Total RNA kit of whole blood, HangZhou Xinjing biochemical reagent Development Co., Ltd.). The mRNA was reverse transcribed into cDNA (TIANGEN gDNA Dispelling RT SuperMix cDNA kit), and then, the level of NLRP3 mRNA (TIANGEN Talent qPCR kit, & CobasZ480) was detected by real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR), and the mRNA level of NLRP3 gene was normalized to β-actin and calculated by the 2−ΔΔCt data analysis method (Table 1 for primers). The serum obtained by procoagulant centrifugation is used to detect inflammatory factors, including IL-1β (ELISA, Shanghai Jihe Biotechnology Co., Ltd. The sensitivity by this assay was 0.42 pg/mL. The antibody showed high specificity to IL-1β but no cross-reactivity with other soluble structural analogues. The detection range was from 0.7 pg/mL to 200 pg/mL, IL-18 (ELISA, Shanghai Jihe Biotechnology Co., Ltd. The sensitivity by this assay is 1.8 pg/mL. The antibody showed high specificity to IL-18 but no cross-reactivity with other soluble structural analogues. The detection range was from 3 pg/mL to 160 pg/mL.), NLRP3 (ELISA, Shanghai Jihe Biotechnology Co., Ltd.). The sensitivity by this assay is 0.3 ng/mL. The antibody showed high specificity to NLRP3 but no cross-reactivity with other soluble structural analogues. The detection range was from 0.625 ng/mL to 100 ng/mL, IL-6 (ECLIA, ROCHE).
Statistical analysis
Statistical examination SPSS27.0 was utilized for statistical analysis of the collected data, while GraphPad Prism 8.0 was utilized for diagramming. The χ2 test was used to examine the difference in genotype and allele frequency between groups. The χ2 test was used to determine whether the genotype distribution conforms to the Hardy-Weinberg equilibrium law, and sample representativeness was confirmed. The odds ratio (OR) and 95% confidence interval (CI) of logistic regression analysis, adjusted for gender and age, were used to estimate the relationship between NLRP3 gene polymorphism and the likelihood of COPD occurrence and development. The measured data conforming to a normal distribution were expressed as X¯±SD, and t-test was used between two groups, and one-way ANOVA test was used for comparison among multiple groups. While the measured data that did not conform to a normal distribution were expressed as median (P25-P75), the Mann-Whitney U test and the Welch ANOVA test were used to compare two groups. For correlation analysis, Pearson correlation analysis is used. p < 0.05 is statistically significant. Receiver operating characteristic curve analysis was used to predict the value, and the area under the curve (AUC) and CI were obtained.
Results
Population characteristics
From August 2021 to March 2023, a total of 168 healthy controls and 221 COPD patients participated. Compared with the control group, the proportion of men in the total COPD group is greater (p = 0.0039), as is the degree of aging (p < 0.0001), but the height and weight have no effect on the incidence of COPD (BMI, p = 0.33). Lung function-related diseases had been excluded from healthy controls and therefore lung function tests were not performed. There is no difference in age, BMI, or smoking as COPD progresses from mild to severe (p = 0.583, 0.141, and 0.272, respectively). Nevertheless, pulmonary function deteriorated significantly (p < 0.001), and FVC, FVCpred%, FEV1, and FEVpred% decreased progressively based on subgroup classification. As a result of pulmonary ventilation dysfunction, PaO2 fell and PaCO2 rose proportionally (Table 2).
Baseline Characteristics of Control Subjects and Each Subgroup of COPD Patients
—, No data were collected. GOLD, the Global Initiative for Chronic Lung Disease; BMI, body mass index; FVC, forced vital capacity; FVC predicted%, the ratio of FVC to its predicted value; FEV1, forced expiratory volume in the first second; FEV1predicted%, the ratio of FEV1 to its predicted value; FEV1/FVC, the ratio of FEV1 to FVC; PaO2, arterial partial pressure of oxygen; PaCO2, arterial partial pressure of carbon dioxide.
NLRP3 3'UTR SNPs verified by the subject samples
To cover as many SNPs as feasible in the 3′UTR, we determined the sequence information of transcripts in the 3′UTR and then determined the corresponding sequence position of transcripts from NCBI. A total of 86 SNPs out of 256 can bind to miRNA. As population verification, we used the NLRP3 3′UTR gene sequences of all the subjects included in this analysis, and ultimately, six SNPs met the inclusion criteria (Fig. 1 and Table 3).

Screening of NLRP3 3′UTR SNPs. SNPs, single nuclear polymorphisms; 3′UTR, 3′ untranslated region.
List of miRNA Target Sequences and SNPs in Binding Sites in the 3’ UTR of NLRP3
8mer site, The 2-8 bases at the 5' end of miRNA are complementary to the target gene, and the first base position of the target gene corresponding to the 5' end of miRNA is A; 7mer-m8 site, The 2-8 bases at the 5' end of miRNA are complementary to the target gene; 7mer-A1 site, The 2-7 bases at the 5' end of miRNA are complementary to the target gene, and the first base position of the target gene corresponding to the 5' end of miRNA is A.
One miRNA can modulate dozens of genes, and multiple miRNAs can recognize the same mRNA target. If there are multiple regulatory relationships, the effect on downstream expression will be greater. rs10754558(G > C) is recognized by miR-3529-3p and miR-549a, and rs1664774076 (ATAT > del) is recognized by miR-5011-5p and miR-1277-5p, according to the findings of this study. According to the results of five miRNA binding site prediction websites, miR-22-3p and miR-223-3p were highly predicted, and rs1664775106 (C > G) and rs200307362 (A > G) were recognized by miR-22-3p and miR-223-3p, respectively. The species with the strongest binding strength is 8mer, whose recognition sites are rs1664774076(ATAT > del) and rs1664775106(C > G) (Table 2). In this investigation, the population distribution of the research object at the locus rs1664775106(C > G) revealed a continuous mutation in the miR-22-3p binding seed zone, designated 459-461(CAG > GCG/GCA). The 3 bp-long mutation is located between the 457th and 464th sites in the NLRP3 3′UTR, and the SNP site of RS 1664775106 (C > G) is at the 457th site (Fig. 2).
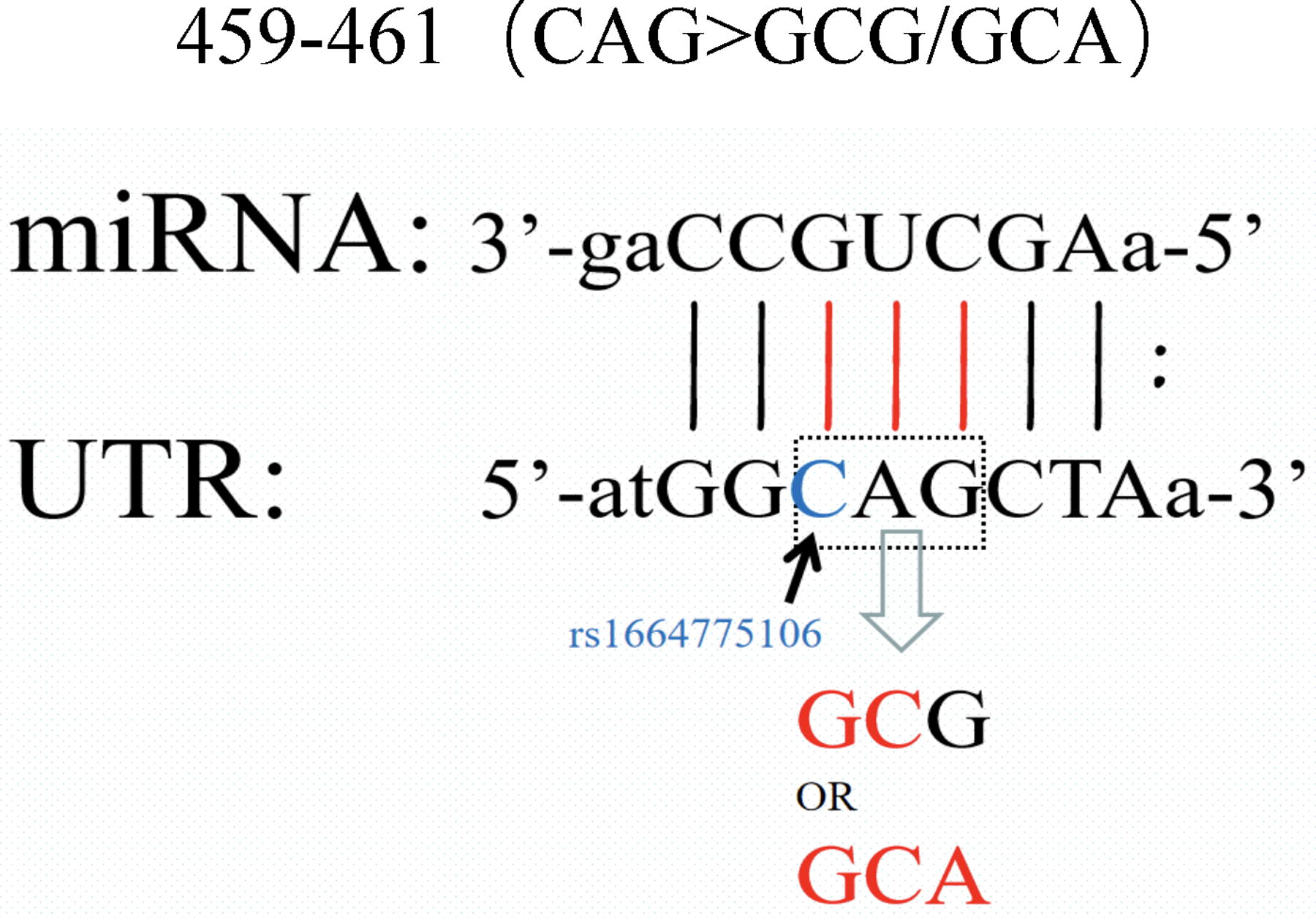
The continuous mutation is located between the 457th and 464th sites in the NLRP3 3'UTR. 3′UTR, 3′ untranslated region.
Association between NLRP3 3’UTR polymorphisms and COPD prevalence
In this study, the gene frequencies of rs10754558 (G > C), rs1664774076 (ATAT > del), and rs1664775106 (C > G) all conform to the Hardy-Weinberg genetic balance law and are representative of the population (p > 0.05, Table 4). However, in the study population, there were no homozygous mutation phenotypes in rs1445597018(G/T), rs1664772078(A > C), and rs200307362(A > G), and the Hardy-Weinberg statistics were p < 0.05. This Hardy-Weinberg rule depends on the allele frequency calculation, when mutation occurred. Therefore, the SNPs of rs1445597018(G > T), rs1664772078(A > C), and rs200307362 (A > G) were not involved in the follow-up study and analysis.
Genotype Frequencies of NLRP3 3′UTR Gene Polymorphisms in Control Subjects and COPD Patients
Adjusted by gender and age. CI, confidence interval; OR, odds ratio.
The frequencies of polymorphism of rs1664774076 (ATAT > del) and rs1664775106(C > G) were significantly different between COPD patients and control group. Polymorphisms of rs1664774076 (ATAT > del) and rs1664775106 (C > G) are associated with the occurrence of COPD. However, only C allele at rs10754558 and GCA genotype at 459-461 were associated with the risk (Table 4).
The GCG mutations of rs10754558 allele C, rs1664774076 allele del, rs1664775106 allele G, and 459-461 have a low prediction of moderate-to-severe COPD development risk, and the degree of risk aggravation is insignificant (GOLD2 vs. GOLD3-4p > 0.05). However, the risk of aggravation of mild COPD patients was significantly predicted (GOLD1 vs. GOLD2 p < 0.0001. GOLD1 vs. GOLD3-4p < 0.0001), except for rs10754558 allele C (GOLD1 vs. GOLD2 p = 0.142. GOLD1 vs. GOLD3-4p = 0.044. In particular, the exposure risk of 459-461 GCA gene type was statistically significant among COPD patients of varying severity (GOLD1 vs. GOLD2 p < 0.0001. GOLD2 vs. GOLD3-4 p = 0.017, GOLD1 vs. GOLD3-4p < 0.0001) (Table 5).
NLRP3 3’UTR SNPs Increase the Risk of COPD Exacerbation
Adjusted by gender and age. CI, confidence interval; OR, odds ratio.
Difference of plasma inflammatory marker level by genotypes of COPD patients
The direct downstream factors of NLRP3 are IL-1β and IL-18, while the main activating factor of IL-6 expression is IL-1β. Therefore, we choose to detect the protein levels of NLRP3 and its downstream IL-1β, IL-18, and IL-6 to evaluate the influence of gene changes on transcription and expression. Considering that inducers or exposure factors were required to stimulate the expression of NLRP3 and other proteins, and that these inflammatory factors were all in a low expression state in the control group (Fig. 3A), this study investigated whether SNP differences in COPD patients affected the expression of NLRP3 mRNA and downstream factors.
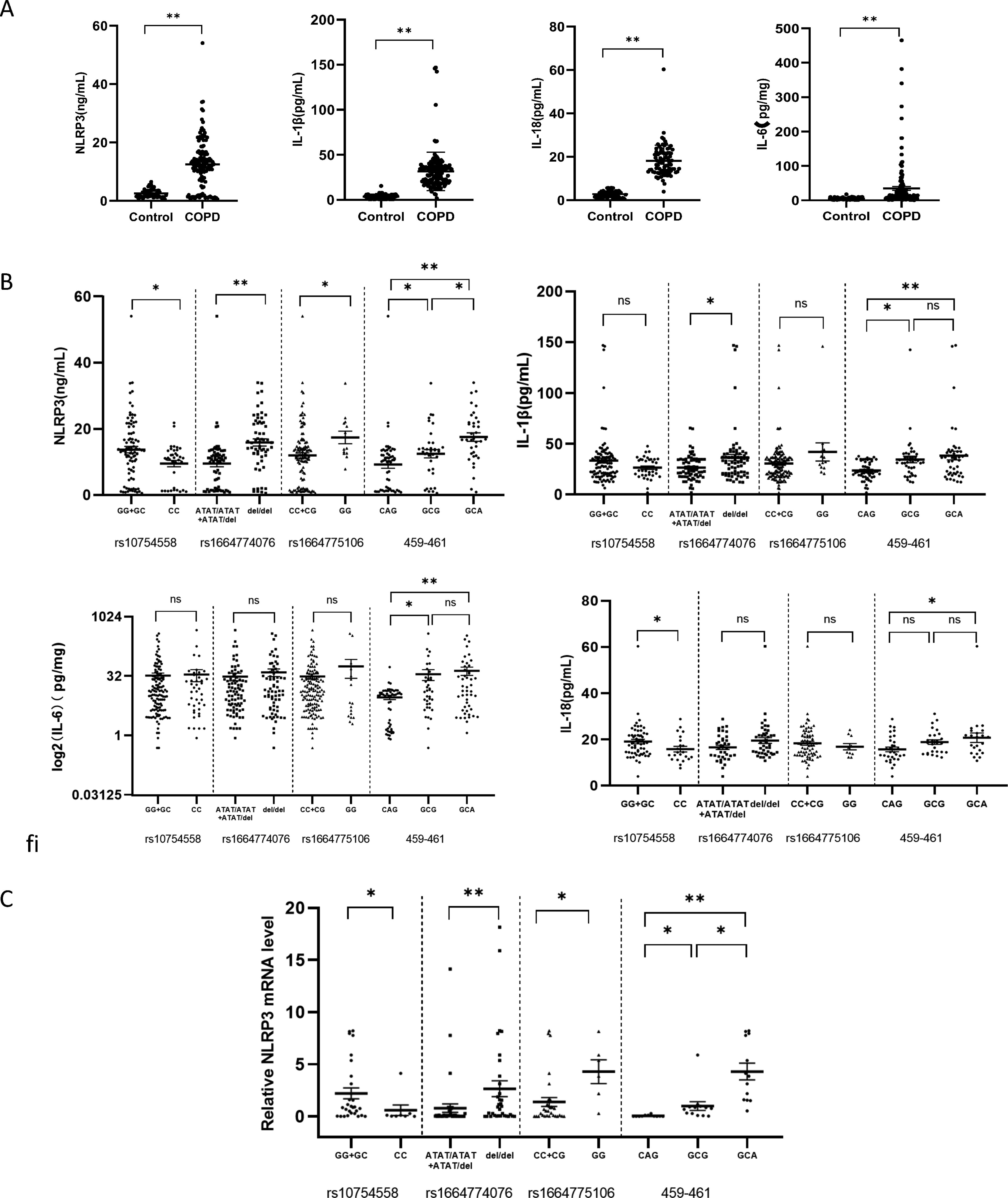
Potential regulatory effects of NLRP3 gene subtypes on downstream expressed proteins.
The plasma NLRP3 expression level was higher in the double-mutant allele groups rs1664774076 and rs1664775106. At sites 459-461, the plasma NLRP3 expression level increased as the number of mutant bases increased. NLPR3 protein expression was substantially different between the 459-461 GCA and rs1664774076 allele del groups (p < 0.001). However, the locus of rs10754558 is opposite, and the expression level of plasma NLRP3 in the double-mutant allele (CC) group is lower than that in the GG+GC group. The expression levels of inflammatory factors IL-1β, IL-18, and IL-6 downstream of NLRP3 were basically the same among the groups, but we found that the above inflammatory factors were all elevated in the GCA group at 459-461 (NLRP3: GCA vs. CAG, p < 0.001. IL-1β: GCA vs. CAG, p < 0.001.IL-18: GCA vs. CAG, p < 0.05. IL-6: GCA vs. CAG, p < 0.001), as shown in Figure 3B.
To further study whether there are differences in the expression of NLRP3 mRNA in different SNPs, 50 samples including different genotypes in this study were selected for analysis. The transcription level of NLRP3 in each SNP group is consistent with the expression of NLRP3 protein (Fig. 3C).
NLRP3 3′UTR mutant genotype affects transcription regulation and is related to lung function.
The relative expression of NLRP3 mRNA was negatively correlated with FEV1/FVC%, FEV1%pred, Pa02/PaCO2 (r = −0.7738, −0.6539, −0.4329, p < 0.05). As NLRP3 mRNA levels increased, lung function decreased. Figure 4 demonstrates that FEV1/FVC% has the strongest correlation with the relative expression of mRNA (r = −0.7738), whereas NLRP3 mRNA expression has a moderate correlation with Pa02/PaCO2 (r = −0.4329). From Figure 3C, we know that the relative expression of NLRP3 3′UTR mutant NLRP3 mRNA has increased significantly (rs1664774076 allele del vs. allele ATAT: p < 0.001. 459-461 GCA vs. CAG: p < 0.001), so we analyzed whether the mutant was related to the decline of lung function. rs1664774076 double-deletion genotype (del/del) and GCA genotype at 459-461 were negatively correlated with FEV1/FVC% (r = −0.6220,−0.6235, p < 0.001, Fig. 5).

Correlation analysis between lung function and relative expression of NLRP3 mRNA.
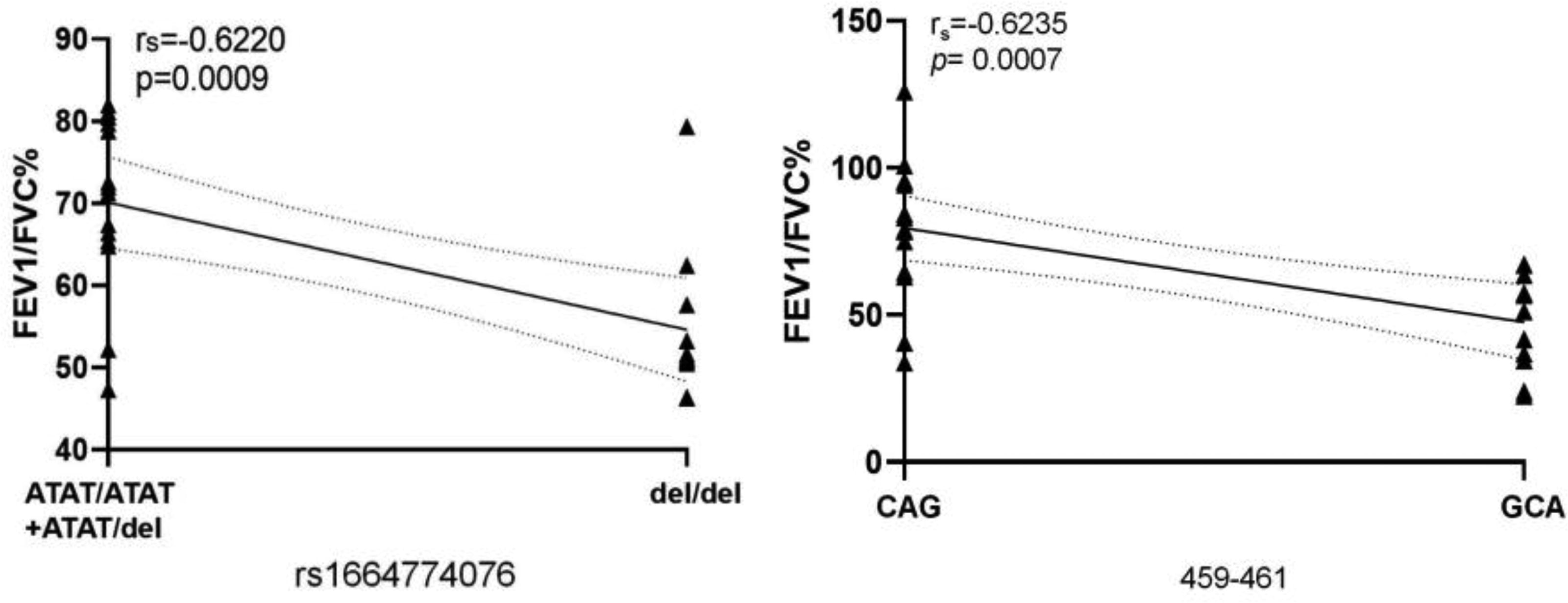
Correlation analysis between lung function and NLRP3 3′UTR mutant genotype. 3′UTR, 3′ untranslated region.
The predictive value NLRP3 3’UTR SNPs for the occurrence and development of COPD
The diagnostic value of rs1664774076 del/del genotype and GCA genotype at 459-461 locus in predicting the risk of COPD alone or in combination is low (AUC is 0.5852, 0.5708, and 0.5972, respectively). Rs1664774076 del/del genotype is more valuable in predicting the development of COPD than the 459-461 GCA genotype (rs1664774076 del/del AUC = 0.7390. 459-461 GCA type AUC = 0.5886), and the NLRP3 inflammatory factor protein has a higher predictive value than its downstream factors IL-6, IL-18, and IL-1β (NLRP3 inflammatory factor protein AUC = 0.9291). The AUCs of IL-6, IL-18, and IL-1β were 0.6797, 0.7458, and 0.7841, respectively. The diagnostic value of combining NLRP3 and rs1664774076 del/del was slightly improved (AUC = 0.9301) (Fig. 6).
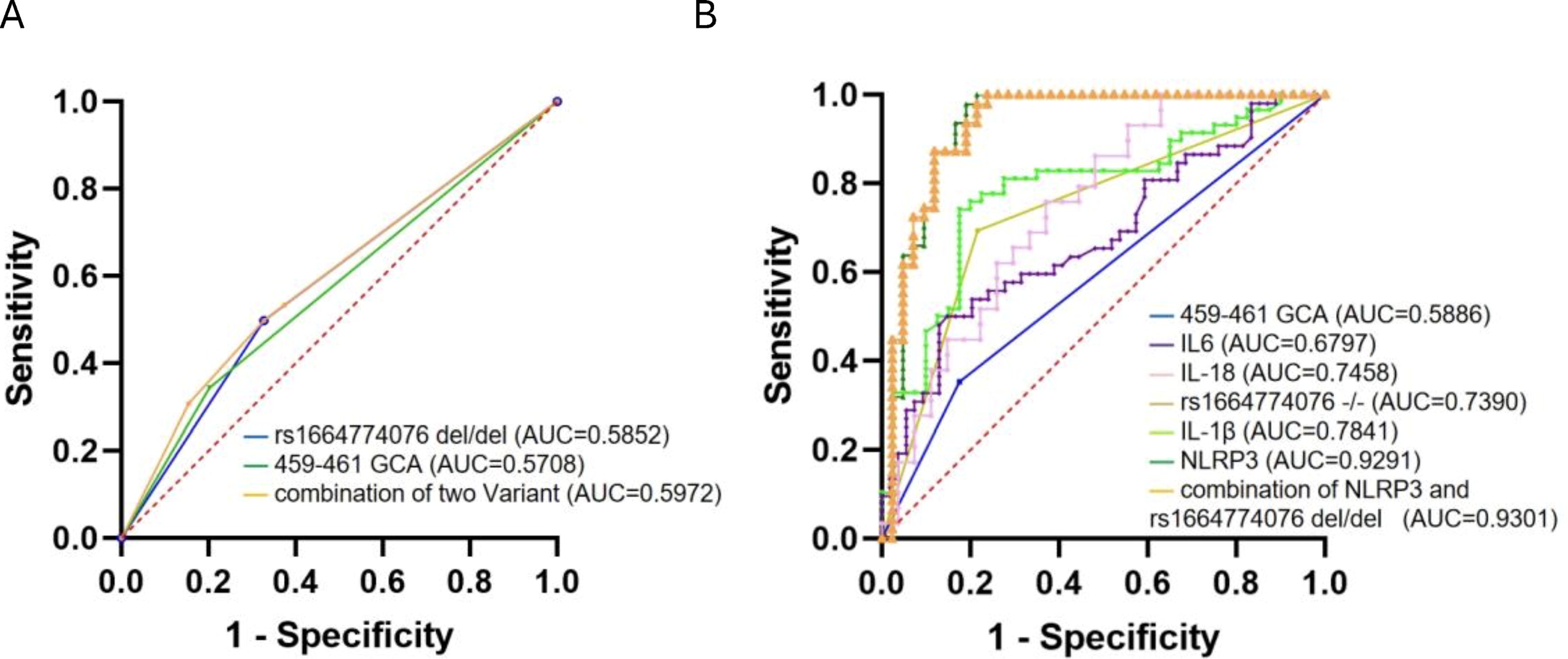
ROC curve of the combination of plasma inflammatory markers and SNPs.
Discussion
There are signaling molecules that activate inflammasomes in the patient’s body, which contribute to the complexity of COPD’s pathogenesis. Pathogen-related molecular pattern stimulation due to infection, such as viruses and bacteria (Kanneganti et al., 2006 ; Martinon et al., 2004), and damage-related molecular pattern stimulation due to oxidative stress, such as ATP and mitochondrial DNA (Riteau et al., 2010), may be the origins of these signaling molecules. This study confirmed the association between NLRP3 and COPD and found that the expression of NLRP3 and its downstream inflammatory factors was substantially higher in COPD patients than in healthy controls. This distinction is because of two factors: first, the stimulation of external risk factors. Second, the presence of SNPs in the NLRP3 population results in variations in inflammatory expression. This can explain why the mutation of the NLRP3 gene does not cause COPD in healthy individuals, and why the development of COPD patients stimulated by the same external factors is heterogeneous.
NLRP3 is located in chromosome region 1q43-q44, with 3 kb upstream of the transcription initiation site (exon and intron) and 2 kb downstream of the termination codon (a total of 37,953 kb) (Xu et al., 2021). Recent genetic research has focused on the association between NLRP3 genetic variation and susceptibility to autoimmune and inflammatory diseases (Hitomi et al., 2009; Queiroz et al., 2020; Wu et al., 2021; Zhao et al., 2017). However, they all investigate the polymorphism of the exon and intron genes in the NLRP3 coding region. This study concentrates on the 3′UTR and locates the functional site, which has been confirmed by the population, as opposed to remaining at the level of biological prediction.
There were continuous mutations at sites 459-461 in the 3′UTR, and the mutant genotypes were GCG or GCA. The rs10754558 CC, rs1664774076 allele del allele (ATAT deletion), and rs1664775106 are examples. COPD susceptibility is associated with the G allele and the 459-461 GCA genotype, which can enhance the risk of COPD. However, for the demonstration that GG is the susceptible genotype (Xu et al., 2021) and other articles (Cheng et al., 2021) have reported higher expression of the GG gene NLRP3, one of the possibilities for the difference in results is the regional variation of samples collected, and the effect of the rs10754558 SNP on COPD remains unknown. In addition, the distribution of SNPs differs between subtypes of COPD and is associated with COPD development. Notable is the fact that genotype 459-461 GCA, as a mutant containing three consecutive bases, has the highest degree of mutation, which can not only trigger the early aggravation of risk but can also foretell the development risk from GOLD2 to GOLD3-4. Consequently, site mutations are more likely to exacerbate the disease.
This study indicates that SNPs can increase the likelihood of disease occurrence or disease development. Polymorphisms in the 3′UTR influence mRNA stability and translation (Kim et al., 2021), which may substantially affect gene expression by removing, diminishing, or producing miRNA-binding sites. This section of the binding site is also known as the seed region. It consists of 6-7 bases that are complementary to the 3′UTR of the target mRNA. The combination of miRNAs can inhibit the expression of a target protein. The mutants of rs1664774076 (ATAT > del), rs1664775106 (C > G), and 459-461 all showed synchronous upregulation of NLRP3 mRNA expression and NLRP3 protein level, indicating that there may be a relationship of gene regulation.
Different SNP genotypes have no effect on the expression of NLRP3's downstream factors (IL-1β, IL-18, and IL-6). Nevertheless, the 459-461 GCA genotype demonstrated a high level of expression in NLRP3 and its downstream inflammatory factors (p < 0.05, Fig. 4B), which may indicate that the triple-base continuous mutation has a greater regulatory influence on the NLRP3 signaling pathway. The mechanism may be that the 459-461 site is located in the seed region of miR-22-3p, and the mutation of three bases results in the loss of the binding site for miR-22-3p, thereby reducing the inhibitory effect of miR-22-3p on the expression of NLRP3 protein and increasing NLRP3. Previous research has also demonstrated that miR-22-3p inhibits the expression of NLRP3 to reduce the inflammatory effects of gout (Wang et al., 2021; Zhang et al., 2021). Rs1664774076 (ATAT > del) is recognized by miR-1277-5p and miR-5011-5p, and the mutation at this site corresponds to the deletion of ATAT. In addition, there is a loss of four bases in the seed region of miRNA, indicating that the mutation at this site may play a larger role in regulating NLRP3. MiR-1277-5p can bind to the 3'UTR of IRS3(recombinant insulin receptor substrate 3) mRNA to inhibit its expression and reduce cell damage, according to studies (Sun et al., 2022). MiR-223-3p has been demonstrated to be a targeted anti-inflammatory therapeutic molecule, and its downregulation can reduce inflammation (Dong et al., 2019; Neudecker et al., 2017; Zhao et al., 2018;). Rs200307362(A > G) is located in the miR-223-3p-identified seed zone. In contrast, rs200307362 (A > G) did not contain any double-mutant GG in this study. It is inferred that the seed region containing rs200307362 can bind tightly to miR-223-3p, and that miR-223-3p inhibits NLRP3.
The level of NLRP3 mRNA increased as lung function declined, and the genotypes of rs1664774076 (ATAT > del) and 459-461 GCA also showed a correlation with lung function decline. The expression levels of NLRP3 mRNA were markedly higher in individuals with the rs1664774076 del/del genotype and the 459-461 GCA genotype. It can be inferred that these two SNPs affect the expression of NLRP3, which ultimately reduces COPD patients’ lung function and exacerbates the disease.
The rs1664774076 (ATAT > del) and 459-461 GCA genotypes can be used for risk prediction, whereas the detection of the NLRP3 gene has a low diagnostic value for COPD and cannot supplant conventional diagnostic methods. COPD acute exacerbation is measured using inflammation indices (such as CRP, IL-6, and IL-1β) (Guo et al., 2022). This study discovered that NLRP3 has a greater predictive value than IL-1β and IL-6. In light of the fact that the expression of inflammatory factors has been stimulated by inducing factors, that is, the expression of inflammatory factors trails behind that of gene detection, it is suggested that the detection of SNPs plus NLRP3 can aid in the clinical evaluation of the progression of a patient’s disease.
To conclude, this study has some limitations. This analysis is limited to the Chinese population and may not be generalizable to other populations. In addition, the scope of this study is limited to the prediction of the miRNA binding site, and it is still necessary to complete the double luciferase reporter gene verification experiment to determine the nature of their relationship. In addition, it is necessary to accomplish the miRNA population survey necessary for this study. If the expression level of the miRNA population is minimal, it will also reduce the regulation of SNPs on inflammatory factors.
Footnotes
Authors’ Contributions
H.W. performed miRNA binding site prediction and SNP screening, data analysis, and article writing. C.H. completed the whole experiment and wrote the original draft. L.Z. participated in the screening and enrollment of subjects, blood sample collecting, and clinical data collection. Both X.Y. and L.P. contributed to project administration. W.L. accomplished the funding acquisition and the writing and review of the article. All the authors read and approved the final article.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of the Fifth Affiliated Hospital of Guangzhou Medical University (protocol KY01-2020-11-06).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The research was funded by the Medical Scientific Research Foundation of Guangdong Province of China (