
Abstract
Background:
Castration-resistant prostate cancer (CRPC), which has developed resistance to next-generation antiandrogens, such as enzalutamide (Enz), is a lethal disease. Furthermore, transcriptional regulation by super enhancers (SEs) is crucial for the growth and spread of prostate cancer, as well as drug resistance. The functions of SEs, a significant class of noncoding DNA cis-regulatory elements, have been the subject of numerous recent studies in the field of cancer research.
Materials and Methods:
The goal of this research was to identify SEs associated with Enz resistance in C4-2B cells using chromatin immunoprecipitation sequencing and cleavage under targets and tagmentation (CUT&Tag). Using HOMER analysis to predict protein/gene-binding motifs, we identified master transcription factors (TFs) that may bind to SE sites. Using small interfering RNA, WST-1 assays, and qRT-PCR, we then confirmed the associations between TFs of SEs and Enz resistance.
Results:
A total of 999 SEs were screened from C4-2B EnzR cells in total. Incorporating analysis with RNA-seq data revealed 41 SEs to be strongly associated with the promotion of Enz resistance. In addition, we finally predicted that master TFs bind to SE-binding regions. Subsequently, we selected zinc finger protein 467 (ZFP467) and SMAD family member 3 to confirm the functional connections of master TFs with Enz resistance through SEs (ZNF467).
Conclusions:
In this study, SMAD3 and ZNF467 were found to be closely related to Enz-resistant CRPC. Our research uncovered a sizable group of SEs linked to Enz resistance in prostate cancer, dissected the mechanisms underlying SE Enz resistance, and shed light on potential clinical uses for SEs.
Introduction
The main treatment for advanced prostate cancer is androgen deprivation therapy, as androgens are crucial to the development of prostate cancer (Armstrong et al., 2010; Donkena et al., 2010). Abnormal androgen receptor (AR) signaling still plays a major role in final-stage castration-resistant prostate cancer (CRPC), resistant to the second generation of the AR antagonist enzalutamide (Enz) (Scher et al., 2012). Although patients with CRPC are initially sensitive to Enz, drug resistance occurs within 6-12 months (Buttigliero et al., 2015), and poor outcomes are experienced by patients with CRPC due to a lack of available therapies following relapse.
Enhancers are a cluster of noncoding cis-acting elements combined with transcription factors (TFs), coactivators, and mediator complexes that interact with RNA polymerase II (RNA pol II) to activate or enhance gene expression (Tower, 2004).
Super enhancers (SEs) are a large cluster of transcriptionally active enhancers enriched with a high density of master TFs, cofactors, and histone modification markers (Hnisz et al., 2013). Recent studies have shown that SEs play an important role in cell differentiation, metabolism, the immune response, and the acquisition of chemoresistance (Hnisz et al., 2013). Therefore, SEs can be used as biomarkers to understand the mechanism of progression in cancer or as potential therapeutic targets. SEs are associated with chemoresistance in various cancers, including ovarian cancer, osteosarcoma, and small cell lung cancer (Bao et al., 2019; Lu et al., 2021; Shang et al., 2019). Some disorders of SEs can lead to the downregulation of chemotherapy-related TFs. For example, in the small cell lung cancer chemoresistant cell line H69AR, TFs such as Foxp1, IRF1, and SP1 are regulated by SEs. When this regulation is dysfunctional, the chemotherapeutic-resistant small cell lung carcinoma drug-resistant strain increases its sensitivity to chemotherapeutic drugs (Bao et al., 2019). SEs may promote enhancer activation and drug resistance, but how and for what reason is unknown. Therefore, it is critical to comprehend how SEs control drug resistance in patients with CRPC to find new therapeutic targets.
In this study, we aimed to explore the relationship between SEs and drug resistance and identify their target genes. We simultaneously predicted TFs that would specifically target these SEs in C4-2B EnzR. Our findings elucidate the potential mechanism of Enz resistance. This study provides new insights and directions for the treatment of patients with Enz-resistant prostate cancer.
Materials and Methods
Cell culture and reagents
C4-2B cells were obtained from the American Type Culture Collection (Rockville, MD, USA). C4-2B EnzR cells were generated by cultivating C4-2B cells for at least 6 months in a medium supplemented with 50 uM Enz. Cells were cultured in RPMI-1640 10% fetal bovine serum in a 37°C and 5% carbon dioxide environment. Enz (MD3100) was obtained from Selleckchem.
Cleavage under targets and tagmentation (CUT&Tag)
The CUT&Tag assay was carried out as previously described, but with modifications. In a nutshell, a total of 100,000 cells were gently washed twice in a wash buffer (20 mM HEPES pH 7.5, 150 mM NaCl, 0.5 mM Spermidine, and 1× Protease Inhibitor Cocktail). Bangs Laboratories’ BP531 Concanavalin A-coated magnetic beads were added to each sample and incubated at room temperature for 10 min. Dig wash buffer (20 mM HEPES pH 7.5; 150 mM NaCl; 0.5 mM Spermidine; 1x Protease inhibitor cocktail; 0.05% Digitonin; 2 mM EDTA) and a 1:50 dilution of primary antibody (H3K4me1: Abcam, ab8895; H3K27ac: Abcam, ab4729) were used to remove unbound supernatant and resuspended bead. Using a magnet stand, the primary antibody was eliminated. Cells were incubated with a secondary antibody (Rabbit Anti-Mouse IgG H&L: Abcam, ab611709) diluted 1:100 in dig wash buffer for 60 min at room temperature. Cells were incubated at room temperature for 60 min with a secondary antibody (Rabbit Anti-Mouse IgG H&L: Abcam, ab611709) that had been diluted 1:100. In dig wash buffer, cells were washed twice or three times using the magnet stand. Dig-med buffer (0.01% digitonin, 20 mM HEPES pH 7.5, 300 mM NaCl, 0.5 mM Spermidine, 1× Protease Inhibitor Cocktail), in which the pA-Tn5 adapter complex was prepared at a 1:100 dilution, was used to incubate cells for 1 h at room temperature. One mL of Dig-med buffer was used to wash the cells 2-3 times for 5 min. After that, cells were reconstituted in tagmentation solution (10 mM MgCl2 in Dig-med buffer), and they were incubated at 37°C for 1 h. Using phenol-chloroform-isoamyl alcohol extraction and ethanol precipitation, DNA was purified.
Two μL each of a universal i5 and specially barcoded i7 primer and 21 μL of DNA were combined to amplify libraries. NEBNext HiFi 2 PCR Master Mix in a volume of 25 μl was then added and blended. The sample was placed in a thermocycler with a heated lid, and the cycling conditions were 72°C for 5 min (gap filling), 98°C for 30 s, 14 cycles of 98°C for 10 s and 63°C for 30 s, 72°C for 1 min of final extension, and hold at 8°C. Cleanup of the libraries was done using XP beads (Beckman Counter, a63881). Agilent 4200 TapeStation analysis was used to determine the size distribution of the libraries, and libraries were mixed to achieve the desired equal representation with the goal of achieving the final concentration advised by the manufacturer. Following the manufacturer’s recommendations, sequencing was done in the Illumina NovaSeq 6000 sequencer using 150 bp paired end.
Data Analysis
FastQ format raw data (raw reads) were first processed using internal Perl scripts. In this step, adapter-, ploy-, and low-quality reads were removed from the raw data to produce clean data (clean reads). The clean data for Q20, Q30, and GC content were calculated concurrently. The clean, high-quality data served as the foundation for all subsequent analyses. The clean reads were then aligned to hg38 reference genome using BWA program. After reads mapping, MACS2 were used to perform peak calling process with peak cutoff q-value < 0.05. After peak calling, ROSE software was used to perform enhancer calling process. The H3K4me1 peak file identified by MACS2 and the H3K27ac BAM file were used as input for the algorithm. By connecting intergenic and intronic H3K4me1 peaks within 12.5 Kb, enhancers were identified as a single entity spanning a genomic region. The level of H3K27ac signal in the genomic region was used to rank stitched enhancers and individual enhancers without neighboring peaks within 12.5 Kb. The remaining enhancers were referred to as typical enhancers (TEs), and SEs were categorized as stitched or individual enhancers with an H3K27ac intensity above a cutoff, where the slope of the distribution plot of H3K27ac intensity exceeded a predetermined threshold. The Integrative Genomics Viewer was used to visualize the signal of H3K27ac and H3K4me1 intensity. The HOMER’s findMotifsGenome.pl tool was used to perform the SE and TE motif enrichment analysis. SEs and TEs were annotated by using homer’s annotatePeaks.pl with default parameters. After SE and TE annotation, the SE-associated genes were subjected to Gene Ontology (GO), HALLMARK, and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses using the R package cluster Profiler (version 4.4) (Yu et al., 2012).
Data preparation
The discovery cohort contained 37 Enz-treated patients with CRPC with histology and clinical outcome patients retrieved from the Abida et al. (2019). To examine the associations between each gene and the Enz resistance-free time of patients with CRPC, a univariate Cox proportional hazard regression analysis was conducted.
Construction of prognostic training model and validation
CoxRidge algorithm was applied to develop prognostic training model based on the 41 SE-associated Enz-R supporters. The ridge regression coefficients of these genes were used to develop a prognostic training model (Li et al., 2022). To assess the prognostic difference between high- and low-risk subgroups, a Kaplan-Meier survival analysis was used. To identify whether the gene signature could be a more accurate predictive model, nine independent prostate cancer cohorts (DKZF, CIT, CancerMap, Belfast, CPC-Gene, Stockholm, Taylor, Cambridge, and GSE54460) were used for external validations.
siRNA transfection
SE transcriptional regulation is important in CRPC for acquired drug resistance. We transfected the related TF with siRNA. siRNAs of control, ZNF467, and SMAD3 (Santa Cruz Biotechnology, sc-37007, sc-89557, and sc-38376) were transiently transfected into C4-2B Enz-R cells by Lipofectamine 2000 (Thermo Fisher Scientific) for 6 h.
RNA extraction and quantitative real-time polymerase chain reaction
Using the NucleoSpin RNA Isolation Kit (TaKaRa), total RNA from cells was extracted in accordance with the instructions in the user’s manual. A spectrophotometer was used to measure the concentration and quality of RNA. RNA samples were reverse transcribed into cDNA using the ReverTra Ace qPCR RT Master Mix Kit (DiagnoCine) according to the product guide. Quantitative real-time polymerase chain reaction (qRT-PCR) was run with ChamQ Universal SYBR qPCR Master Mix (Vazyme, #Q711). The 2-(ddCt) relative expression of mRNA was determined using GAPDH as the loading control. Data of qRT-PCR were obtained in triplicated experiments. Supplemental File S1 lists the PCR primers used for qRT-PCR.
In vitro proliferation assay and statistics
The WST-1 Cell Proliferation Assay Kit (WST-1) method was used for the in vitro proliferation assay using the WST-1 Kit, in accordance with the manufacturer’s instructions (Dojindo). Data were calculated as mean standard deviation (SD). To determine statistical significance, the Student’s t-test was used.
Results
Epigenetic landscape of C4-2B Enz-resistant cells
The flow chart of the whole study is shown in Figure 1. Enz-resistant C4-2B cells (C4-2B EnzR) were created by treating C4-2B cells with Enz for more than 6 months to define the mechanisms that drive prostate cancer cell resistance to the next-generation antiandrogen Enz. Proliferation assays were used to calculate the IC50 of Enz in C4-2B EnzR cells (see Supplemental File S2). The most informative histone marks are H3K4me1 and H3K27ac, which are a crucial part of the reference epigenome (http://ihec-epigenomes.org/research/reference-epigenome-standards/). Using H3K27ac and H3K4me1 ChIP-seq data, SEs were identified using the ROSE (Rank Ordering of Super-Enhancers) algorithm (Lovén et al., 2013; Whyte et al., 2013). To evaluate the epigenetic landscape and the role of SEs in Enz resistance in prostate cancer, CUT&Tag analysis was performed to profile the active histone modifications, H3K27ac and H3K4me1, in C4-2B EnzR cells. H3K4me1 is associated with poised/active enhancers, whereas H3K27ac is associated with active regulatory elements (both enhancers and promoters). An analysis of CUT&Tag profiles across 3-kb regions between the transcription start site (TSS) and transcription end site (TES) revealed that the H3K4me1 and H3K27ac marks on the entire genome of C4-2B EnzR cells displayed the same profile around the TSS, with a major peak located downstream of the TSS (Fig. 2A, B). Next, when we evaluated the read distribution across all peaks of H3K4me1 and H3K27ac CUT&Tag signals in C4-2B EnzR cells, the results showed that most of the reads were mainly concentrated near the center of the peaks, indicating that the signals in the peak area were relatively concentrated (Fig. 2C, D). Furthermore, the annotation of the peaks showed that promoter (30.44%) and distal intergenic (20.44%) regions tended to have enhancer-related H3K4me1 modification, whereas promoter (37.87%) and intron (36.59%) regions tended to have enhancer-related H3K27ac modification (Fig. 2E). Finally, the majority of H3K27ac peaks on enhancer regions overlapped with H3K4me1 peaks, indicating active enhancers, while H3K4me1 peaks that did not overlap were linked to poised enhancers (Fig. 2F) (Kimura, 2013).
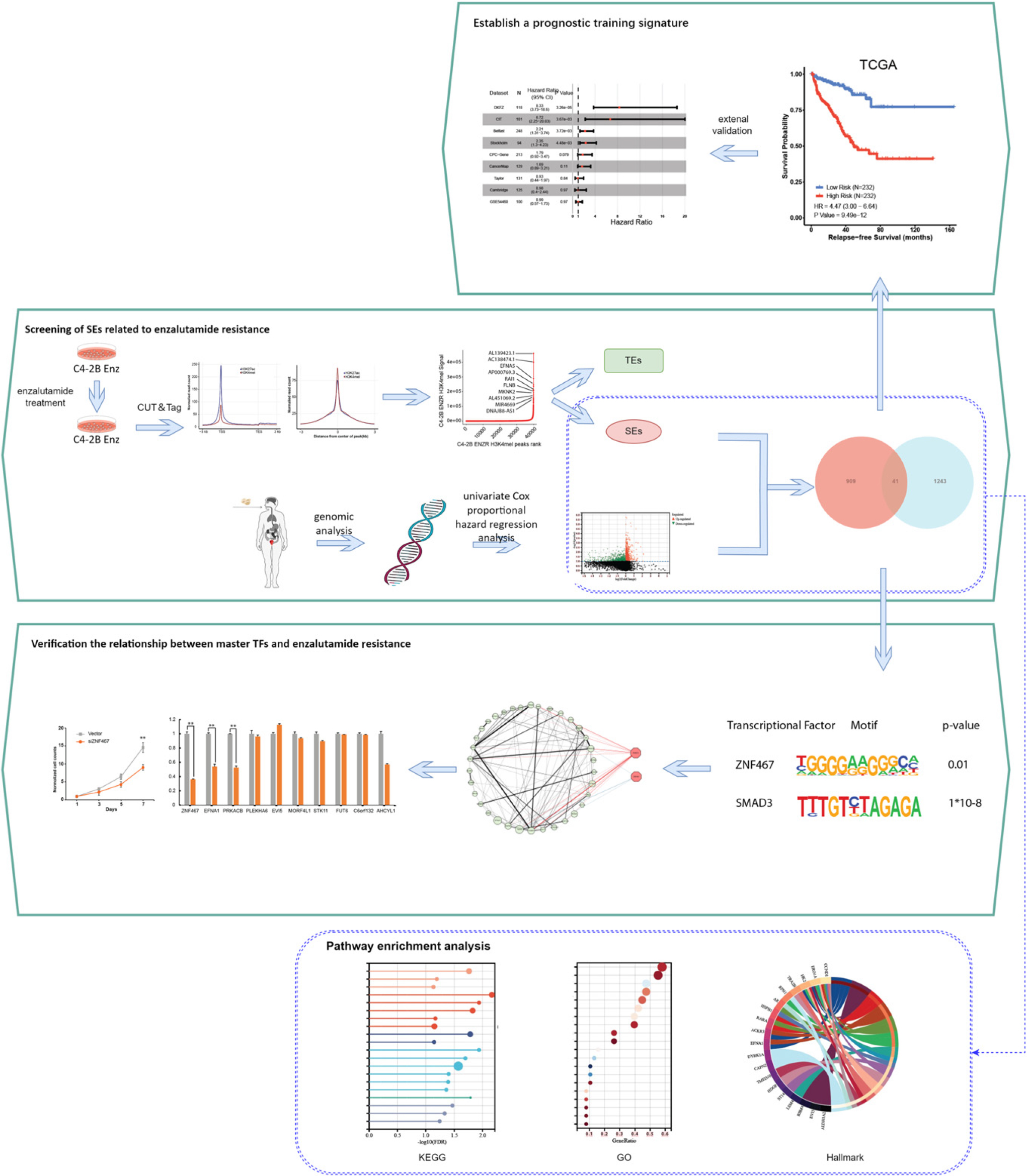
The data preprocessing and analysis process.
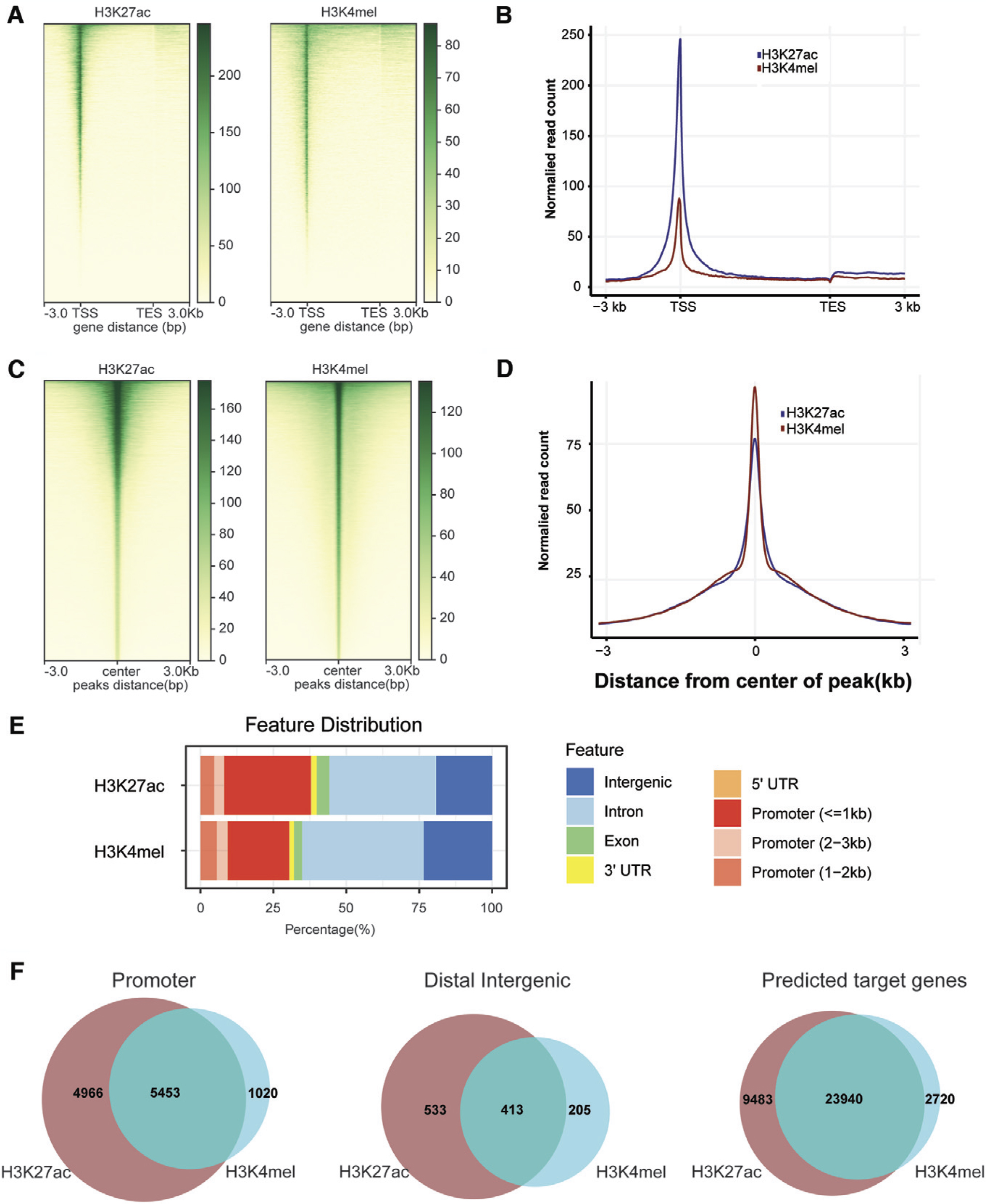
Histone mark enrichment distribution in C4-2B EnzR cells.
Screening of SEs in C4-2B EnzR cells
To further investigate the role of SEs in C4-2B EnzR cells, we predicted the SEs and TEs by analyzing H3K4me1 and H3K27ac CUT&Tag data of C4-2B EnzR cells with the ROSE algorithm and found that 999 SEs and 38905 TEs were identified from C4-2B EnzR cells with a cutoff value of 14328.171 (Fig. 3A). Detailed information on the SEs is shown in Supplemental File S3. We further displayed the average CUT&Tag signal for both TEs and SEs. The results showed that the length of the spanned DNA domains of these SEs (median length was 23,804 bp) was dramatically higher compared with the TEs (median length: 816 bp), indicating a stronger enrichment density in SEs than in TEs (Fig. 3B).
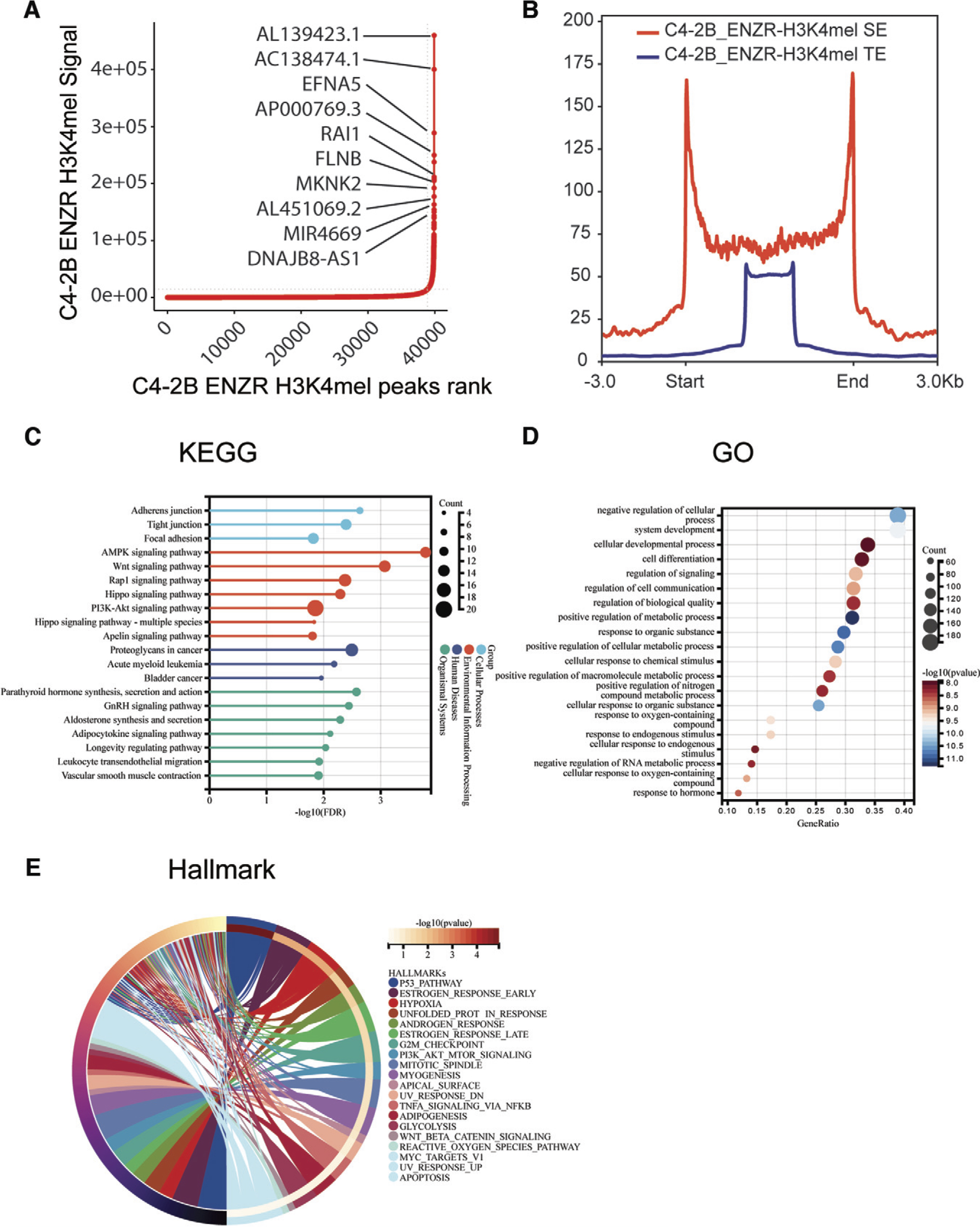
SEs identified in C4-2B EnzR cells.
We attempted to understand the underlying mechanisms by which Enz resistance is driven in prostate cancer. KEGG pathway analysis of SEs revealed that environmental information processing may be greatly influenced by pathways, including the AMPK, Wnt, Rap1, Hippo, PI3K-Akt, and apelin signaling pathways (Fig. 3C). Among these pathways, the AMP, Wnt, Hippo, and PI3K-Akt signaling pathways have been reported to mediate the development of Enz resistance in prostate cancer (Adelaiye-Ogala et al., 2020; Bainbridge et al., 2020; Imamura and Sadar, 2016; Wang et al., 2021b; Zhang et al., 2018). This finding demonstrated that the Rap1 and Apelin signaling pathways may be important pathways of Enz resistance. The associated genes were then subjected to GO analysis, and we discovered that the associated genes were enriched in the negative regulation of cellular processes, system development, cellular developmental processes, cell differentiation, and signaling regulation (Fig. 3D). These findings showed that alterations in cellular biological functions occurred concurrently with the development of Enz resistance. Furthermore, HALLMARK analysis showed that the p53, early estrogen response, hypoxia, unfolded protein response, and androgen response pathways were enriched (Fig. 3E). The results indicated that the p53, hypoxia, and androgen response pathways play important roles in the progression of Enz resistance, consistent with previous reports (Farah et al., 2022; Geng et al., 2018; Li et al., 2019). The estrogen and unfolded protein responses may be potential mechanisms involved in drug resistance.
Thus, after screening 999 SEs associated with Enz resistance, we proposed possible mechanisms through which Enz resistance was driven in prostate cancer. Moreover, our results suggested that the Rap1 signaling pathway, Apelin signaling pathway, estrogen response and unfolded protein response could have significant effects on Enz resistance in prostate cancer.
Differential expression pattern analysis based on the clinical outcome of Enz-resistant CRPC
To validate the prognostic efficacy of a gene set that can predict prognosis in Enz-resistant CRPC, genomic analysis of a patient cohort containing 37 Enz-treated patients with CRPC with histology and clinical outcomes was performed (Abida et al., 2019). Using univariate Cox proportional hazard regression analysis, we identified 1283 genes as predictors of negative Enz resistance-free outcome (EnzR supporters), whereas 669 genes were predictors of a positive outcome (EnzR suppressors) with log-rank p value <0.1 (Fig. 4A).

Bioinformatics analysis of EnzR supporters and suppressors in patients with prostate cancer.
To investigate the potential functions of EnzR supporters in prostate cancer, GO, KEGG, and HALLMARK enrichments were used for analysis. From pathway enrichment in GO analysis, nuclear part, nuclear lumen, macromolecule biosynthetic process, cytosol, and protein-containing complex were identified as the top five pathways (Fig. 4B). Intriguingly, the KEGG analysis revealed significant enrichment for spliceosome, basal TFs, cell cycle, endocytosis, and protein processing in endoplasmic reticulum pathways (Fig. 4C). In addition, HALLMARK enrichment identified additional crucial pathways: E2F targets, G2/M checkpoint, Myc targets, MTORC1 signaling, and apical junction (Fig. 4D).
Taken together, the findings of the bioinformatics analysis revealed that the majority of EnzR supporters fell under the categories of cellular components and transcriptional regulation (Yuan et al., 2019), cell cycle (Toren et al., 2015), endocytosis (Su et al., 2022), protein processing in endoplasmic reticulum (Lin et al., 2021), E2F targets (Xu et al., 2020), Myc targets (Furlan et al., 2021), and MTORC1 signaling pathways (Fruman and Rommel, 2014), which were involved in Enz resistance in previous studies. Our findings also suggested that spliceosome, apical junction, and macromolecule biosynthetic process pathways may contribute to the onset of Enz resistance in prostate cancer.
Identification of SEs related to Enz resistance
We investigated the roles of SEs in Enz resistance by combining 999 specific SEs with 1283 EnzR supporters from the Enz-treated CRPC patient cohort (see Supplemental File S4). Forty-one genes were obtained by Venny analysis (Fig. 5A).

Screening and analyzing of differential expression SE-associated genes.
We used these SE-associated EnzR supporters to evaluate the biochemical recurrence (BCR) survival of patients after radical prostatectomy in different datasets. First, 41 of the SE-associated EnzR supporters shown above were included to establish a prognostic training model in The Cancer Genome Atlas (TCGA) dataset by using the CoxRidge algorithm. Kaplan-Meier survival analysis of BCR-free survival showed that patients with high gene expression exhibited a significantly worse outcome in the training cohort (TCGA) (Fig. 5B). Consistently, the signature predicted BCR in multiple authoritative validation cohorts, such as DKZF, CIT, CancerMap, Belfast, CPC-Gene, and Stockholm (Fig. 5C).
Next, these genes were also classified by GO, KEGG, and HALLMARK analyses. GO analysis showed that metabolism and response to chemical stimulus pathways may be important in developing Enz resistance (Fig. 5D). According to KEGG analysis, the functions of the screened SE-associated EnzR supporters were mostly related to different signaling pathways (Fig. 5E). Among these pathways, the AMPK (Nguyen et al., 2014), Wnt (Zhang et al., 2018), FOXO (Monga et al., 2022), and MAPK (Li et al., 2019) signaling pathways that are involved in the control of Enz resistance were discovered. The Hedgehog signaling pathway is thought to be involved in the occurrence and development of CRPC (Guo et al., 2017), which implies its potential effects on Enz resistance, which has not been well studied. Protein processing in the endoplasmic reticulum is also enriched in SE-associated EnzR supporters, which is consistent with our analysis of EnzR supporters and a previous study (Lin et al., 2021). The results of the HALLMARK analysis indicated that hypoxia, K-Ras, apoptosis, TNFA signaling through KFKB, estrogen response, MTORC1 signaling, and unfolded protein response were the top enriched pathways (Fig. 5F). Hypoxia, apoptosis, and tumor necrosis factor (TNF)-α signaling through the NF-κB and MTORC1 signaling pathways are known to regulate the progression of Enz resistance. K-Ras is one of the Ras superfamily isoforms (N-RAS, H-RAS, and K-RAS) that is frequently mutated in the cancer pathway. K-Ras is an important factor in mediating cancer drug resistance due to its promotive effects on cellular proliferation and survival pathways (Kim et al., 2021), which indicates its potential key role in Enz resistance in prostate cancer. Our results again indicated that the estrogen response and unfolded protein response could have significant effects on Enz resistance in prostate cancer.
SEs predicted master TFs using HOMER
SEs aid in cell identity maintenance, and SE-driven transcription alters not only protein-coding genes but also noncoding regulatory components, resulting in the state of cancer cells (Kimura, 2013; Su et al., 2022). Interactions between TFs and the transcriptional machinery that are extremely well organized, especially master TFs, are required for SE-driven transcription regulation (Hay et al., 2016; Huang et al., 2018). As a result, we applied HOMER analysis to these SEs to forecast their master TFs (Heinz et al., 2010). Two master TFs were predicted (Fig. 6A). Subsequently, we performed the expression correlation analysis of 41 SE-associated EnzR supporters and the master TFs by constructing a protein-protein interaction network. We found that SMAD3 and ZNF467 were predominantly positive with 41 SE-associated EnzR supporters (Fig. 6B). In addition, 41 SE-associated EnzR supporters exhibited mainly positive correlations with each other (Fig. 6B).
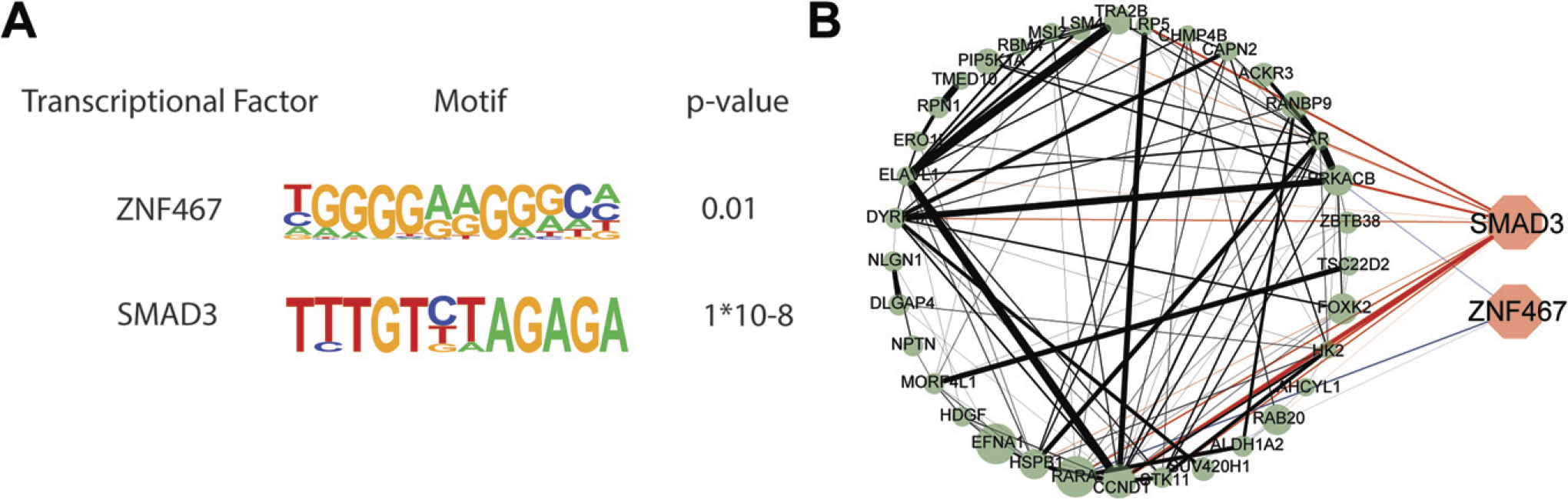
Predicted TFs and their interaction with SE-associated Enz-resistance-induced genes.
Confirmation of the connection between master TFs and Enz resistance
We chose ZNF467 and SMAD3 from the master TFs to study the connections between TFs and Enz resistance to ascertain the roles of the projected master TFs. First, we discovered that ZNF467 and SMAD3 have a close relationship to other SE genes. Second, there is a strong correlation between ZNF467 and the prognosis of prostate cancer (see Supplemental File S5).
SMAD3 and Enz resistance
Breast, bladder, and gastric cancers have been associated with the transcriptional repressor SMAD3, which has been classified as an oncogene (Hu et al., 2021; Wang et al., 2021a; Zhao et al., 2018). We used small interfering RNA to knock down SMAD3 expression in C4-2B EnzR cells to see how it affected Enz resistance. Then, we analyzed SMAD3-knockdown cells by WST-1 assays. The results demonstrated that SMAD3 significantly inhibited the proliferation of C4-2B EnzR cells compared with the vector control (p > 0.05; Fig. 7A).
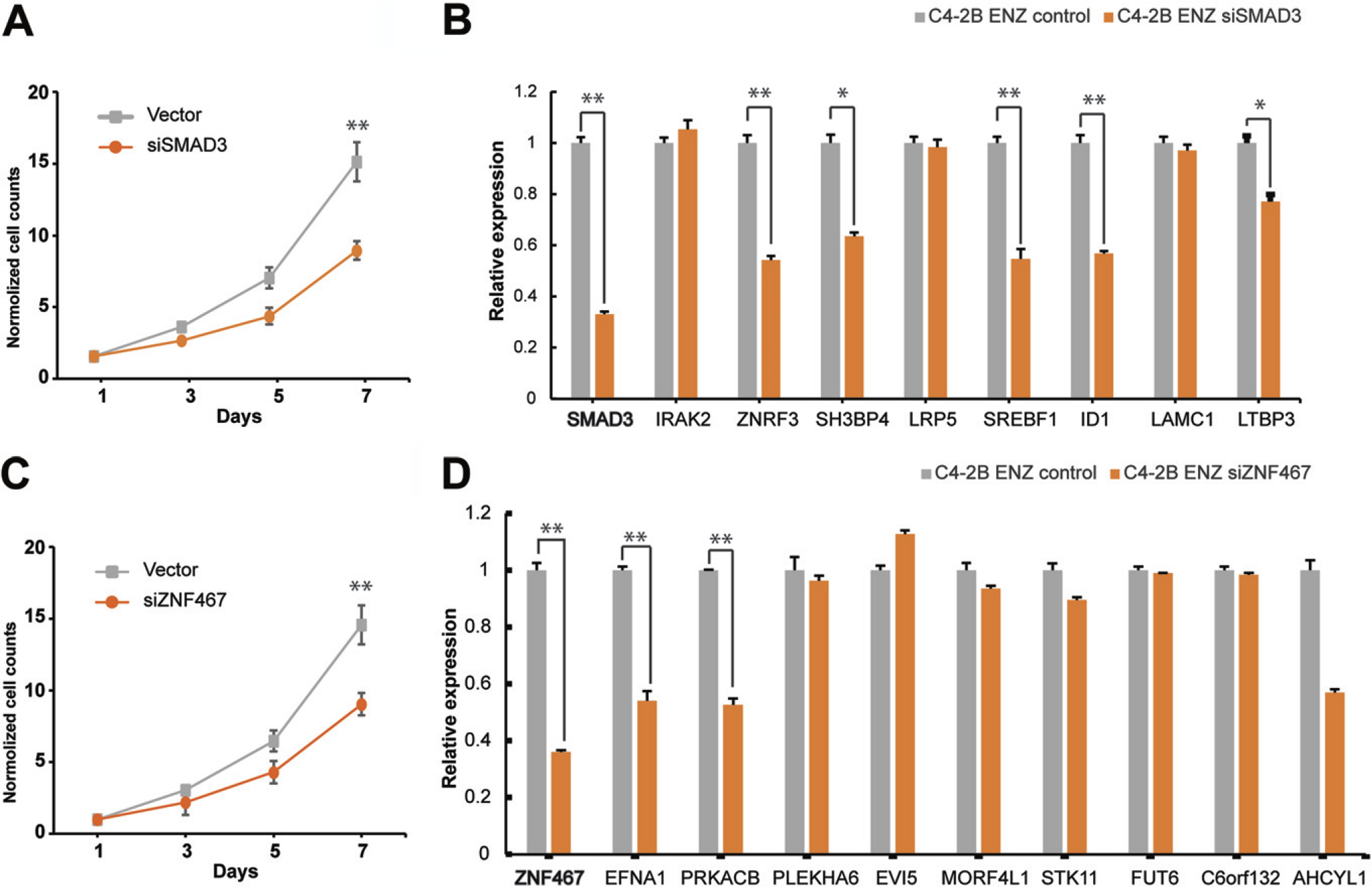
Proliferation assays and qRT-PCR analyses in C4-2B EnzR cells transfected with siRNAs targeting SMAD3 and ZNF467.
To confirm whether SMAD3 is associated with Enz resistance, the expression of SMAD3 was detected using qRT-PCR (highlighted) in Figure 7B, and it was observed that knockdown of SMAD3 significantly reduced the expression levels of ZNRF3, SH3BP4, SREBF1, ID1, and LTBP3 mRNAs in C4-2B EnzR cells. The genes, ZNRF3, SREBF1, and ID1, are strongly linked to drug resistance (Liu et al., 2021; Yin et al., 2017; Zhou et al., 2019). ID1, in particular, is related to taxol resistance in prostate cancer (Zhang et al., 2006). Thus, our results showed that SMAD3 induces Enz resistance in prostate cancer.
ZNF467 and Enz resistance
ZNF467’s function in human cells is still unknown. In mice, it was isolated as an Oncostatin M-inducible mRNA from hematogenic endothelial LO cells (Nakayama et al., 2002, p. 3). WST-1 assays were used to examine ZNF467-knockdown cells. ZNF467 significantly reduced C4-2B EnzR cell proliferation when compared with the vector control (p > 0.05; Fig. 7C).
To ascertain whether ZNF467 is connected to Enz resistance, qRT-PCR was modified to measure the expression levels of related genes. The result is that after knocking out ZNF467, as highlighted in Figure 7D, there was a conspicuous decrease in the expression levels of EFNA1 and PRKACB mRNAs in C4-2B EnzR cells. It has been previously reported that these genes are closely related to drug resistance (Gao et al., 2021; Makondi et al., 2017). Therefore, our results showed that ZNF467 induces Enz resistance in prostate cancer. The precise mechanisms by which ZNF467 targets PRKACB and EFNA1 are still unknown.
Discussion
Although the majority of patients with CRPC are sensitive to Enz treatment, drug resistance will inevitably occur after a period of medication, and there is no effective alternative treatment after drug resistance. As a result, it is essential to look into the causes of Enz resistance to increase patients’ survival time and quality of life. Current Enz resistance mechanisms include androgen receptor, DNA repair, cell cycle, PI3K-AKT-mTOR, and Wnt-catenin pathways (Schmidt et al., 2021). However, this is insufficient to fully explain the mechanism. SE is an area of active research that has been linked to the development of drug resistance. As a result, we investigated how SEs contributed to the emergence of Enz resistance in CRPC.
Enz-resistant C4-2B cells were constructed over 3 months and CUT&Tag analysis was used to identify 999 SEs. Cut&Tag analysis can effectively reveal the regulatory information in the genome (Janssens et al., 2018). We chose CUT&Tag instead of chromatin immunoprecipitation-sequencing (ChIP-seq) because it is easier to operate and has fewer steps. Generally, CHIP-seq takes 3-4 days (Landt et al., 2012), whereas CUT only takes 1 day to perform all the steps. The most important step is to minimize batch and handling effects, thereby maximizing repeatability (Kaya-Okur et al., 2019). It is critical to identify and filter SEs. A total of 41 SE-associated EnzR supporters were identified using these SEs combined with clinical and histological analysis data combined from Abida et al. (2019). To determine the clinical value of these SEs, we used these genes to construct a signature through Cox-Ridge regression to predict BCR in patients with prostate cancer. To verify the predictive performance of the signature, we tested the model in 9 external multicenter cohorts and found that it had a significant predictive effect in multiple external validations (DKFZ, CIT, Belfast, and Stockholm), indicating that these genes constructed in this signature are closely related to the biochemical recurrence of prostate cancer.
GO, KEGG, and HALLMARK function analyses revealed that many pathways may be involved in drug resistance, including AMPK (Tan et al., 2019; Wu et al., 2020), Hedgehog (Wang et al., 2020), and TNF (Borst, 2004). For example, overexpression of PrLZ inhibits autophagy by regulating the AMPK pathway, leading to docetaxel resistance (Zeng et al., 2018). In addition, the estrogen response and unfolded protein response (UPR) appeared in our analysis many times, which may be associated with Enz resistance. The estrogen response has been shown to be associated with chemoresistance (Sisinni et al., 2019), but there is no evidence that it is associated with Enz resistance in prostate cancer. UPR is a potential mechanism through the activation of the PI3K/AKT axis, and the overexpression of downstream oncogenes overrides the activity of trastuzumab (Kumandan et al., 2013).
SE is critical in the development of drug resistance. This SE-driven drug resistance mechanism suggests possible solutions to drug resistance. The screening of numerous SEs linked to Enz resistance in CRPC has been made easier by the use of next-generation sequencing technologies. Therefore, our findings suggest a novel therapeutic strategy for elucidating drug resistance mechanisms and clinical applications of SEs. They work in tandem with regulatory factors and major regulatory factors, which are also known as master TFs (Saint-André et al., 2016). The binding of specific TFs to SEs triggers the interaction between promoters and enhancers, loading SEs to homologous promoters to initiate and activate the transcription of downstream target genes (Saint-André et al., 2016). The development of drug resistance and TFs are both closely related to the development of cancer (Bradner et al., 2017).
In our research, our predictions for master TFs that bind to SEs and activate target genes were confirmed. To examine their relationship with Enz resistance, we chose two master TFs (ZNF467 and SMAD3). Prostate cancer progression is accelerated by SMAD3 by controlling the expression of angiogenic molecules, which is overexpressed in advanced prostate cancer (Lu et al., 2007). It has been reported that SMAD3 is associated with drug resistance. For example, SMAD3 exerts targeted therapy resistance in melanoma by promoting EMT-like processes (Gautron et al., 2021). We speculate that SMAD3 also plays an important role in Enz resistance in prostate cancer.
The Krüppel-like family of TFs, which includes 12 repeats of C2H2-type zinc finger motifs, includes the mouse ortholog of ZNF467 (Nakayama et al., 2002). It is reported that ZNF467 contributes to cellular function in general because it is widely expressed (Nakayama et al., 2002). In addition, it has been demonstrated to control adipocyte and osteoblast commitment. Osteoblast differentiation was delayed by overexpression of ZNF467, whereas adipocyte differentiation was accelerated (Quach et al., 2011), and the suppression of ZNF467 promoted osteoblast differentiation (You et al., 2012). According to a more recent study, ZNF467 may control Wnt signaling by encouraging Sost to bind to LRP5/6 (You et al., 2012). Our results showed that inhibiting ZNF467 and SMAD3 could reduce the resistance to Enz, indicating that these master TFs may be potential targets for the treatment of Enz resistance.
In conclusion, our findings highlight the role of SEs in patients with CRPC and Enz resistance and offer insights into how SEs might be used as biomarkers to create new therapeutic approaches to treat patients with Enz resistance. To demonstrate their distinct molecular functions and mechanisms for controlling gene expression, there is not enough genetic evidence. Thus, several follow-up studies are required. It is necessary to first show that SEs and master TFs do in fact combine. The relationship between these TFs and SEs will be examined using ChIP-seq and antibodies against SMAD3 and ZNF467. Second, we will also simultaneously construct LNCap Enz-resistant cell line. After that, we plan to perform CUT&Tag sequencing to identify the common SEs of Enz resistance to determine the common targets of Enz resistance. Finally, we hope to verify whether these TFs can be used as therapeutic targets for Enz resistance in vivo and clinically.
Footnotes
Acknowledgments
The authors thank all the staff who participated in this study from the First Affiliated Hospital of Guangzhou Medical University, the Second Affiliated Hospital of Guangzhou Medical University, and the Guangzhou First People’s Hospital.
Availability of Data and Materials
The datasets generated and analyzed during the current study are available in the GEO repository (record GSE228308). The following secure token has been created to allow review of record GSE228308 while it remains in private status: mdghwuwylrkhncp. The clinical and sequencing data of 37 patients were obtained from the public dataset from GitHub (![]() ).
).
Authors’ Contributions
Chao Cai: Conceptualization, Supervision, Funding acquisition, and Software. Qinwei Liu: Data Curation, Writing—original draft, and Visualization. Haoran Shan: Validation. Chuanfan Zhong: Methodology and Formal analysis. Guidong Chen: Methodology and Formal analysis. Zhouda Cai: Software and Validation. Yu Zheng: Resources. Jianming Lu: Funding Acquisition and Investigation. Jiaojiao Tang: Project Administration and Validation. Zhuoyuan Lin: Funding Acquisition, Writing—Review & Editing, and Supervision.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.