
Abstract
Background:
The global pandemic of Coronavirus Disease 2019 (COVID-19) has resulted in significant fatality rates. Clinical outcomes for affected individuals range from being asymptomatic to severe illnesses requiring intensive care unit (ICU) admission. Among the various factors contributing to the variation in clinical outcomes, host genetics play a prominent role. Interleukin-6 (IL6), a key player in immune responses, has been identified as having a crucial impact on viral infections, including the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Specifically, certain variations known as single nucleotide polymorphisms (SNPs) in the IL6 promoter region have been found to significantly influence IL6 expression and the severity of viral infections.
Materials and Methods:
To explore the relationship between these genetic variations and COVID-19 in asymptomatic and ICU-admitted Kurdish patients, genetic sequencing was performed to determine the genotypes of nine IL6 SNPs.
Results:
The study findings revealed that although the proportion of the GG genotype of rs1800795 was slightly higher in asymptomatic COVID-19 cases, the difference was not statistically significant (chi2 = 2.666, p = 0.236). Notably, Kurdish patients displayed a uniform genetic makeup (monomorphic) for the dominant alleles of rs2069830 (C), rs142759801 (C), rs2069857 (C), rs2069829 (G), rs2234683 (G), rs13447446 (T), rs527770772 (C), and rs13447445 (C). Furthermore, patients carrying the haplotype GCGGCTCCC were found to have a 0.481-fold higher likelihood of being asymptomatic with COVID-19 (p = 0.016, OR = 0.481).
Conclusions:
This study demonstrates that the rs1800795 SNP is not statistically associated with COVID-19 at the genotype level. However, the presence of the dominant G allele of rs1800795 in the haplotype was found to be statistically associated with asymptomatic COVID-19 patients.
Introduction
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the causative agent of Coronavirus Disease 2019 (COVID-19), a global pandemic with significant fatality rates. Patients infected with SARS-CoV-2 exhibit a wide range of clinical symptoms and illness courses, which are not only explained by age and the presence of comorbidities (Ali et al., 2022a; Ali et al., 2024a; Zhu et al., 2020). Due to the broad spectrum of symptoms reported by patients, it is likely that host genetics significantly influence the susceptibility and severity of COVID-19 (Abdullah et al., 2020; Ali et al., 2023;Ovsyannikova et al., 2020, Rashid and Salih, 2022; Rashid and Salih, 2023c).
Interleukin-6 (IL6) serves as a critical element in the cytokine network of immune defense. It is increasingly evident that the IL6 pathway is pivotal not only in normal immune regulation but also in immunological dysregulation (Choy et al., 2020). One of the most reliable indicators of the stress response is IL6, which is frequently elevated in response to viral infection and inflammation (Ali et al., 2022b; Tanaka et al., 2014). The immunological responses that are induced by a SARS-CoV-2 infection help in the clearance of the virus but are likely to cause cytokine release syndrome (CRS) in individuals with COVID-19 (Lim et al., 2021). Patients with COVID-19 who required intensive care exhibited higher levels of IL6. It is possible that a substantial number of pathogenic T cells and inflammatory monocytes secreting elevated levels of IL6 will enter the pulmonary circulation, triggering an inflammatory storm. This can lead to immunological dysfunction, extensive damage, the potential failure of multiple organ systems, and high mortality (Ali et al., 2022b). Therefore, IL6 functions as a critical inflammatory mediator in COVID-19 patients, contributing to cytokine release syndrome (CRS), and may be used to accurately assess disease severity and predict prognosis (Fujino et al., 2021). In addition, IL6 is likely to contribute to COVID-19-associated coagulopathy by producing tissue thrombin, stimulating platelet activation, and inducing endothelial dysfunction (Zhang et al., 2020). Serum IL6 levels have been shown to be elevated in patients with severe COVID-19, and this has been shown to be substantially connected with severe clinical outcomes such as ICU admission, ARDS, and mortality (Chen et al., 2020).
The IL6 gene is found on 7p21 of human chromosome 7. The transcription of the IL6 gene is primarily controlled by components in the 5-prime region. Several polymorphisms are present in the first 1.2 kb of the IL6 promoter. The most extensive SNP investigated was rs1800795 (-147G/C), which influences levels of IL6 gene expression. It is the candidate SNP that has the potential to affect the severity of inflammatory, autoimmune, and infectious diseases (Fujino et al., 2021; Sghaier et al., 2017). IL6 gene expression is altered in a cell type-specific way by various promoter haplotypes. Many researchers have attempted to determine the impact of these polymorphisms on gene expression, but the results have been inconsistent (Fife et al., 2005). This inconsistency is due to differences in ethnicity and genetic variation (Chen et al., 2018). We conducted an IL6 SNP analysis on the Kurdish population in response to recommendations from other researchers to investigate the impact of IL6 SNPs in different ethnic groups, aiming to ascertain the association of IL6 polymorphisms with asymptomatic and ICU COVID-19 patients.
Material and Methods
Study participants and sampling
In this research, nasopharyngeal swab samples were taken from 48 critically infected COVID-19 Kurdish patients who were on ventilators in the ICU of Shahid Hadi Hospital in Sulaimani province, KRG, Iraq. Another 48 samples were taken from asymptomatic Kurdish passengers who had tested positive for SARS-CoV-2 during travel screening tests for COVID-19. They were followed up to ensure they remained asymptomatic cases. All patients were unvaccinated and aged 20 to 60. The nasopharyngeal swabs were stored in viral transport medium (VTM) at −70°C in the COVID-19 molecular diagnostic laboratory of Shahid Tahir Hospital.
RNA extraction and SARS-CoV-2 detection
A Viral RNA Extraction Kit (T-200-96; Zybio Inc., China) and an automated extractor, the Zybio EXM6000 system, were used to extract viral RNA from 200 μL of VTM. SARS-CoV-2 detections were performed using the one-step reverse transcription Real-Time PCR Kit for Novel Coronavirus 2019-NcoV (Aehealth, Germany). The TaqMan probe and primer were set as follows: the FAM-labeled probe for detection of the ORF1ab gene of the virus. The probe was labeled with VIC to detect the N gene. A CY5-labeled probe was used for detection of the RNase gene in humans as an internal control to validate the extraction procedure (Rashid and Salih, 2023b). The Bio-Rad thermocycler was programmed at 50°C for 10 min, 95°C for 3 min, then 45 cycles of 95°C for 10 sec, and 60°C for 40 sec. A result was deemed positive when the cycle threshold [Ct] fell below 40.
DNA extraction
The positive SARS-CoV-2 samples from the screening step were subjected to DNA extraction using an AddPrep Genomic DNA Extraction Kit (lot No. G201906A) according to the manufacturer’s instructions (Addbio, South Korea).
Amplification of IL6 gene
The PCR amplification reaction was conducted according to the manufacturer’s instructions using the Add Star Taq Master Mix PCR Kit (Addbio, Korea). In brief, the PCR involved mixing 10 μL of master mix, 5.0 μL of DNA sample, 3 μL of DEPC-H2O, and 1 μL of 10 pmol of each forward primer (IL6-F TTGTCAAGACATGCCAAAGTG) and reverse primer (IL6-R GATGCCGTCGAGGATGTACC). The primers were designed using the Primer3Plus online database and verified with the NCBI Primer Blast web-based primer design tools to amplify a 722 bp segment of the IL6 gene. The thermocycler (Bio-Rad, USA) was programmed for an initial denaturation phase of 5 min at 95°C, followed by 40 cycles of denaturation at 94°C for 30 s, annealing at 59°C for 30 s, extension at 72°C for 45 s, and a final extension phase of 3 min at 72°C. The PCR products were analyzed by loading 4.0 μL of the PCR product on a 1% agarose gel in 1× TBE buffer. The gel was stained with 7 μL of safe gel dye (Addbio, Korea), and electrophoresis was run at 120 volts for an hour on the electrophoresis system. A 100 bp DNA ladder was used to demonstrate the migration pattern of the PCR amplicons.
Sequencing and genotyping
The PCR products were purified using the AddPrep PCR Purification Kit following the manufacturer’s instructions (Addbio, South Korea). Subsequently, they were sequenced using the Sanger method at a Macrogen sequencing facility in South Korea. The identity of each nucleotide sequence was confirmed using the same forward and reverse primers used in the amplification techniques. The generated DNA sequences were assessed for quality and analyzed using FinchTV. Homozygous SNPs were identified based on high single-base peaks with high Phred quality scores, whereas heterozygous SNPs were identified based on overlapping peaks of two nucleotide bases with lower Phred quality scores compared to adjacent bases. All SNPs in this study were specified by the DNA plus strand (cDNA strands). For example, guanine (G) represented the major frequency allele and cytosine (C) represented the minor frequency allele for the rs1800795 SNP, and so on for other SNPs.
Statistical analysis
The wpcalc database (https://wpcalc.com/en/equilibrium-hardy-weinberg/) was utilized to calculate Hardy-Weinberg equilibrium. The SHEsis online database (http://analysis.bio-x.cn/myAnalysis.php) was used for allele frequency, genotype distribution, and haplotype analyses (Shen et al., 2016). Statistical analysis was performed using the Statistical Package for Social Sciences, version 25.0 (SPSS, Chicago, IL) (Ali et al., 2024a; Ali et al., 2024b). The chi-square test was utilized to assess the association of each SNP, genotype, and allele with ICU and asymptomatic groups. The statistically significant level used in this study was set at p ≤ 0.05. Logistic regression analysis was conducted to analyze the odds ratios (OR) and 95% confidence intervals (CI) to determine the role, strength of association, and risk of the variants in both groups.
Results
This study aimed to analyze IL6 polymorphisms between two groups: asymptomatic COVID-19 patient groups and ICU-admitted patient groups. Nine SNPs were genotyped, including two missense SNPs in IL6, rs2069830, c.94C > T (P32S), and rs142759801, c.91C > A (P30T). These two SNPs also overlapped with the intron of interleukin 6 antisense RNA 1 (IL6-AS1), an antisense long noncoding RNA (lncRNA) (Fig. 1A). The rs2069857 c.-48 variant is also found in the intron of the IL6-AS1 overlap sequence and the prime untranslated region of IL6. The remaining six SNPs were in the IL6 promoter region near the transcription start point and overlapped with the intron of IL6-AS1. These include the following variants: −190C > T (rs2069829), −176G > C (rs2234683), −174G > C (rs1800795), −111G > T (rs13447446), and c.−85T > C (rs527770772), −63C > G (rs13447445). All SNPs were found to be in Hardy-Weinberg equilibrium. Table 1 provides a summary of the SNP genotyping results.
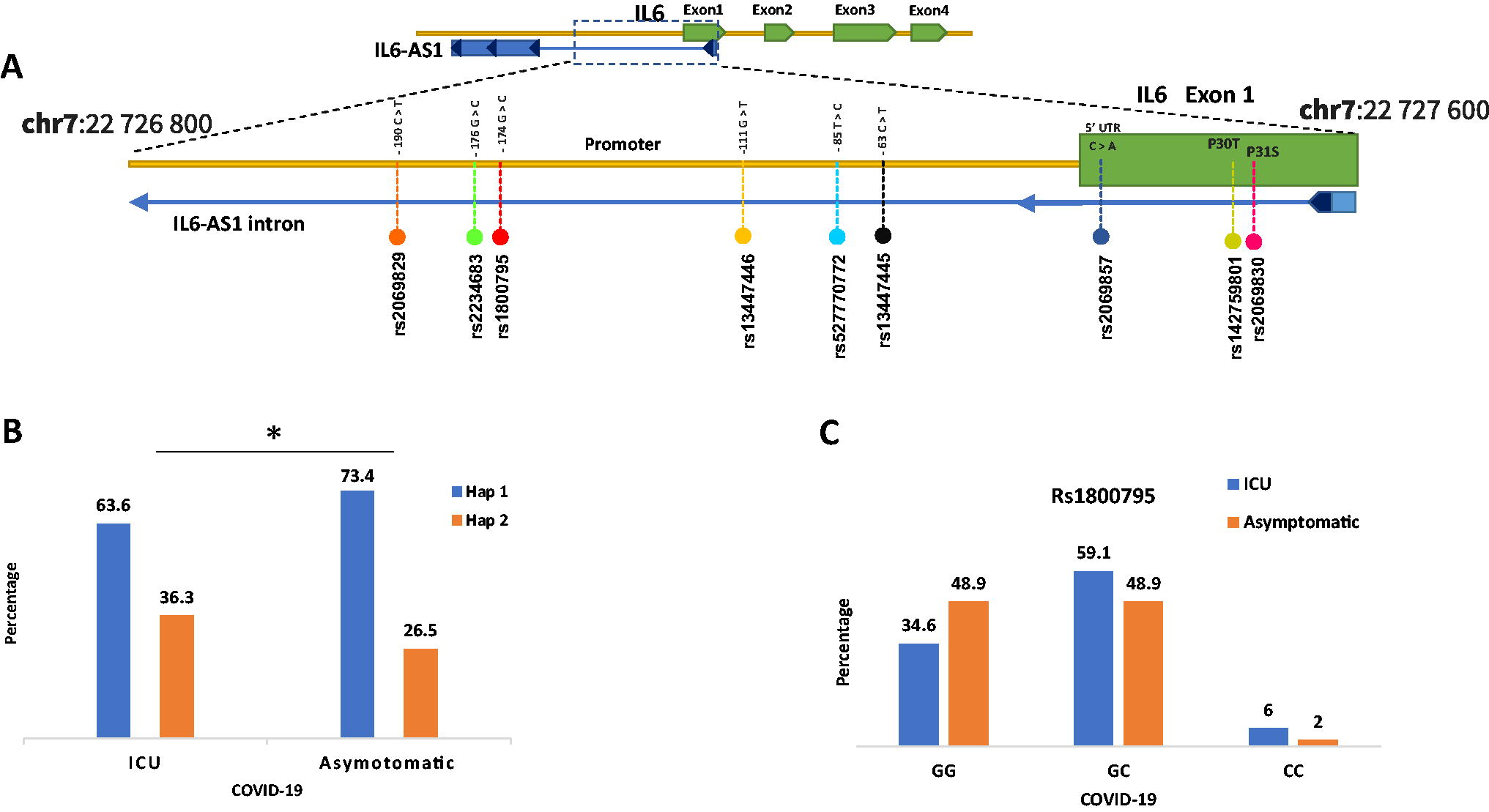
Representation of the IL6 SNPs
The Genotyping Results of IL6 SNPs
ICU, intensive care unit; IL6, Interleukin-6; SNP, single nucleotide polymorphisms.
The asymptomatic group exhibited a larger proportion of GG genotypes for rs1800795, while the ICU group had a higher percentage of CC genotypes (Fig. 1C). However, chi-square analysis revealed that rs1800795 had no statistically significant association with either group (p = 0.263) (Table 1). Logistic regression analysis was performed to determine the role and degree of association between the variants. The odds ratios were not significant, according to the findings. Genotype combinations of the dominant model (CG+CC vs. GG) were also examined using MedCalc biostatistical analysis; the results were p = 0.177, OR = 1.75. This indicates that compared with the GC and CC genotypes, the dominant model of the GG genotype was 1.75 times more likely to develop asymptomatic COVID-19. However, this risk is statistically insignificant.
All patients in both groups were homozygous for the following main alleles: rs2069830 (C), rs142759801 (C), rs2069857 (C), rs2069829 (G), rs2234683 (G), rs13447446 (T), rs527770772 (C), and rs13447445 (C). Because there was no polymorphism in these nucleotides in Kurdish patients, statistical analysis was not applicable (Table 1).
This study performed haplotype analysis for all the SNPs to assess possible significant haplotypes. As a result, the SHEsis database demonstrated two haplotypes (Table 2). Only the first haplotype (Hap 1) showed a statistically significant higher frequency in the asymptomatic groups with p = 0.01 and OR = 0.481 (Table 2). The G allele of rs1800795 in Hap-1 was shown to be associated with an asymptomatic result.
Haplotypes Were Carried out Using Nine SNPs in the Following Order: rs1800795, rs2069829, rs2234683, rs13447446, rs13447445, rs527770772, rs142759801, and rs2069830 by SHEsis
The significant haplotype designated in bold.
ICU, intensive care unit.
Discussion
The IL6 gene encodes interleukin-6, which has dual roles as an anti- and pro-inflammatory cytokine. Epidemiological and gene polymorphism studies have revealed variations in the severity of lung diseases such as chronic obstructive pulmonary disease and asthma among individuals (Jin and Wang, 2003). IL6 polymorphisms have been associated with certain viruses. These include the hepatitis C virus and influenza viruses, as well as the hepatitis B virus (Linnik and Egli, 2016; Rashid and Salih, 2023a; Riazalhosseini et al., 2018).
IL6 may lead to cytokine storms, which are the major immunopathologic symptoms observed in individuals with severe SARS-CoV, MERS-CoV, and SARS-CoV-2 infections, impairing immune response control and potentially resulting in fatal outcomes (Barrett, 2019; Channappanavar and Perlman, 2017; Gupta et al., 2020). Therefore, IL6 levels are associated with the progression of an infection from moderate to severe (Liu et al., 2020). Some studies have linked elevated IL6 levels with certain inflammatory conditions to polymorphisms in the IL6 gene promoter region (Foster et al., 2000; Sghaier et al., 2017). Since IL6 is a key part of controlling CD4-T cells, it seems likely that learning more about the genetic differences in the IL6 gene could shed light on the cause of the disease and the outcome of COVID-19 (Kirtipal and Bharadwaj, 2021).
This study investigated nine SNPs in the promoter of the IL6 gene to determine the possible association of COVID-19 outcomes between ICU-admitted patients and asymptomatic COVID-19 individuals in the Kurdish population. The rs1800795 SNP has been extensively studied for various diseases across different ethnic groups and populations; however, its association remains controversial depending on the pathological condition and genetic polymorphism of the studied population. Our results revealed a higher percentage of the GG genotype of the rs1800795 SNP in the asymptomatic patient group (Fig. 1C). However, this difference was statistically insignificant. This finding is consistent with Iranian research that demonstrated a nonsignificant association between the GG genotype and mild COVID-19 (Falahi et al., 2022).
Additional research in children with pneumonia discovered that the GG genotype in Egyptians is protective against acute respiratory failure and ICU admission (Zidan et al., 2014). In contrast to our findings, research in the Turkish population found a significant association between the IL6 polymorphism at rs1800795 and developing severe COVID-19. The researchers found that those with macrophage activation syndrome (MAS) had a greater frequency of the GG genotype and G allele than those without MAS and that the G allele was associated with elevated IL6 levels and an increased likelihood of developing MAS in COVID-19 patients (Kerget and Kerget, 2021). The ICU group had a higher frequency of the C allele in our study, which is consistent with the findings of a meta-analysis study that found the C allele to be associated with severe pneumonia (Table 1) (Mao et al., 2017). The C-minor allele of rs1800795 was correlated with mortality in the COVID-19 study, but this correlation did not reach statistical significance (Smieszek et al., 2021). However, additional research has linked the CC genotype of rs1800795 to a reduced immune response to chronic Hepatitis C Virus (Bogdanović et al., 2016).
In this study, all the other eight SNPs were monomorphic in Kurdish patients; all had the homozygote for the dominant allele at rs2069830 (C), rs142759801 (C), rs2069857 (C), rs2069829 (G), rs2234683 (G), rs13447446 (T), rs527770772 (C), and rs13447445 (C). Similar results were obtained in cohort research examining the impact of IL6 polymorphism on HCV (Yee et al., 2009), where alleles for rs13447445, rs2069829, and rs13447446 were monomorphic in Caucasian Americans. To our best knowledge, there is no study on the effect of rs527770772 on IL6 or disease conditions. Furthermore, the analysis of the missense mutation suggested that rs142759801 may affect IL6 expression and function by interfering with normal splicing pathways (Strafella et al., 2020). In contrast to this study, other research discovered polymorphism in rs2234683, but found no statistical association with neurodevelopmental delay (Clark et al., 2010).
Haplotype analysis for SNPs revealed a statistically significant association of Hap-1 with the asymptomatic group (Fig. 1B); this haplotype included the G allele of rs1800795, as well as the dominant alleles of the other eight SNPs (Table 2). Similar to the present study, previous research on chronic renal failure also discovered a statistically significant haplotype linked with the condition, even though none of the haplotype’s SNPs, including rs1800795, was statistically correlated with the disease at the genotype level (Ng et al., 2008). The results of this research conflict with those of a previous study that found that haplotypes carrying the dominant G allele of rs1800795 experience more severe HCV progression in Italians (Cussigh et al., 2011). Most of the previous studies focused on the correlation of the SNPs directly with the promoter of IL6. However, these SNPs also overlap with IL6-AS1, which has a regulatory effect on the expression of IL6. In order to stabilize IL6 mRNA, IL6-AS1 works as an endogenous sponge by competitively binding to microRNA (miR-149-5p) (Ahmed et al., 2022). IL6-AS1 regulates IL6 promoter activity by recruiting transcription factors in the nucleus and increasing histone methylation levels. Expression of IL6 is regulated by IL6-AS1 through epigenetic, transcriptional, and post-transcriptional pathways. Overall, IL6 levels are associated with IL6-AS1 expression (Yi et al., 2021). One of the limitations of this study is that both groups were small in size; thus, further research is needed on the whole spectrum of SNPs in the IL6 gene in a more representative sample of the population.
Conclusion
The current research was conducted in response to previous work, aiming to perform host genetic analysis on IL6 promoter polymorphisms across various countries and ethnicities to investigate their influence on COVID-19 outcomes. This study demonstrated that rs1800795 is not statistically associated with COVID-19 at the genotype level. However, the haplotype harboring the dominant G allele of rs1800795 may be associated with asymptomatic COVID-19 in Kurdish patients.
Footnotes
Acknowledgments
The authors would like to thank Dr. Dana I. Omer for consultation of statistical methods. As well as many thanks to Dr. Nahla M. Saeed for providing access to some laboratory facilities.
Authors’ Contributions
P.M.A.R. contributed in conceptualization, data curation, formal analysis, and writing—original draft. G.F.S contributed in supervision, visualization, and writing—review and editing.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Preprint
An earlier draft of this article was posted as preprint at Research Square (doi: 10.21203/rs.3.rs-3246475/v1).
Ethics Approval Statement
In this study, all methods were carried out in accordance with the relevant institutional and national guidelines and regulations. In addition, we vouch for the Ethics Licensing Committee of the Health Directorate’s approval for all experimental protocols (No. 8375 on May 20, 2021). We received verbal consent from an illiterate individual to utilize their samples that had previously been taken from them for diagnostic purposes.
Author Disclosure Statement
The authors declare no conflict of interest. The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
No funding was received for this article.