
Abstract
Background:
Vitamin D receptor (VDR) is a nuclear hormone receptor widely expressed in the substantia nigra. Its association with an increased risk of Parkinson's disease (PD) is based on vitamin D deficiency and/or different polymorphisms in its gene receptor. This fact has been demonstrated by several case-control studies.
Materials and Methods:
Consequently, we investigated the association between VDR ApaI, BsmI, FokI, and TaqI gene polymorphisms and PD in a Spanish cohort that included 54 cases and 17 healthy controls. The detection of single nucleotide polymorphisms (SNPs) was performed using a polymerase chain reaction-restriction fragment length polymorphism.
Results:
Our data indicate that the SNPs were not associated with the age of onset of PD, nor with the occurrence of motor symptoms. However, only BsmI polymorphism was significantly associated with PD in this Spanish cohort. In fact, BsmI genotype was five times higher among PD patients than among controls, and the A allele was considered as a genetic risk for PD. Additionally, the combination of FokI and BsmI polymorphisms was significantly associated with PD and could represent a risk factor.
Conclusion:
We conclude that ApaI, TaqI, and FokI polymorphisms were not associated with PD, but BsmI could be a risk factor for PD in this randomized population.
Introduction
Vitamin D receptor (VDR) is a nuclear hormone receptor that modulates gene transcriptions in response to 1,25-dihydroxyvitamin D3 (1,25[OH]2D3), the active form of Vitamin D (Butler et al., 2011). In fact, 25-hydroxyvitamin D3 (25[OH]D3) is metabolized to 1,25(OH)2D3 by 1-α-hydroxylase (Yeshokumar et al., 2015). Both 1-α-hydroxylase and VDR are abundantly expressed in the human brain, particularly in the substantia nigra (Cui et al., 2013, Eyles et al., 2005). The activation of VDR by 1,25[OH]2D3 is critical for the transcription of vitamin D responsive genes. This ligand-binding process enables VDR to promote histone modifications via the recruitment of co-activator proteins and the activity of histone acetyltransferases (HATs), thus activating gene expression. Interestingly, the unbound VDR, generally located in the cytoplasm, can also bind DNA and interact with histone deacetylases (HDACs) through co-repressor proteins, leading to gene repression (Fetahu et al., 2014). This interaction between VDR and HDACs (specifically HDAC4 and HDAC6) is known to decrease cyclically in response to 1,25[OH]2D3 in MCF-10A cells (Malinen et al., 2011).
Importantly, an imbalance in HAT and HDAC activity has been observed in Parkinson's disease (PD). This imbalance is evidenced by the epigenetic alterations in fibroblasts from PD patients (Yakhine-Diop et al., 2019b), including increased levels of certain class I (HDAC2 and HDAC3) and II (HDAC4) HDAC proteins (Yakhine-Diop et al., 2019a), which correlate with decreased levels of acetylated proteins (Yakhine-Diop et al., 2019b) and peptides (Yakhine-Diop et al., 2018) in idiopathic PD.
The link between VDR and PD pathogenesis is supported by the observations of low levels of 25[OH]D3 in PD patients' plasma (Ding et al., 2013) and polymorphisms in the VDR gene affecting receptor activity (Smolders et al., 2009). Reduced vitamin D levels correlate with postural instability (Peterson et al., 2013) and symptom severity (Suzuki et al., 2012) in PD patients. Interestingly, the administration of 1,25[OH]2D3 has a neuroprotective effect in animal models of PD, attenuating the inflammation-mediated neurodegeneration (Kim et al., 2006) and promoting neurogenesis (Cataldi et al., 2018). Furthermore, Vitamin D has been found to protect 6-Hydroxydopamine (6-OHDA)-injured rats by enhancing dopamine levels via increased endogenous glial cell line-derived neurotrophic factor expression in the substantia nigra (Smith et al., 2006, Wang et al., 2001). However, VDR gene variants can impair its function and the enzymatic regulation of vitamin D metabolism (Smolders et al., 2009).
Since then, evidences suggest that VDR polymorphisms play a role in PD pathogenesis (Butler et al., 2011), despite some inconsistencies in the data. For example, the FokI C allele is significantly represented in Hungarian PD patients (Torok et al., 2013) and is also associated with the milder form of PD in Japanese population (Suzuki et al., 2012). The VDR BsmI GG genotype in Korean PD patients (Kim et al., 2005) further reinforces the role of these polymorphisms. Contrarily, studies in Faroese (Petersen et al., 2014) and Taiwanese populations (Lin et al., 2014) did not find an association between VDR polymorphisms and PD risk. These differences observed across distinct PD populations could be attributed to the strength of linkage disequilibrium between adjacent genetic variants in the VDR gene (Fang et al., 2005).
The VDR gene, situated on the long arm of the chromosome 12, contains eight coding exons (2-9) and six isoforms (1f-1c) in its 5′ noncoding exon1 (Fig. 1) (Deuster et al., 2017). Among ∼57 single nucleotide polymorphisms (SNPs) identified in VDR gene (Fang et al., 2005), our research concentrated on the four well-known restriction fragment length polymorphisms (RFLPs): TaqI (rs731236), ApaI (rs7974232), BsmI (rs1544410), and FokI (rs10735810). In this study, we investigated these VDR polymorphisms in Spanish sporadic PD patients. Notably, the prevalence of the BsmI genotype was fivefold higher in PD patients compared to controls, a finding that was statistically significant. This suggests that the BsmI polymorphism could be a potential risk factor for PD within the Spanish population. Furthermore, our analysis revealed a significant association between the combination of BsmI and FokI polymorphisms and sporadic PD, indicating their combined relevance as a risk factor in this population.
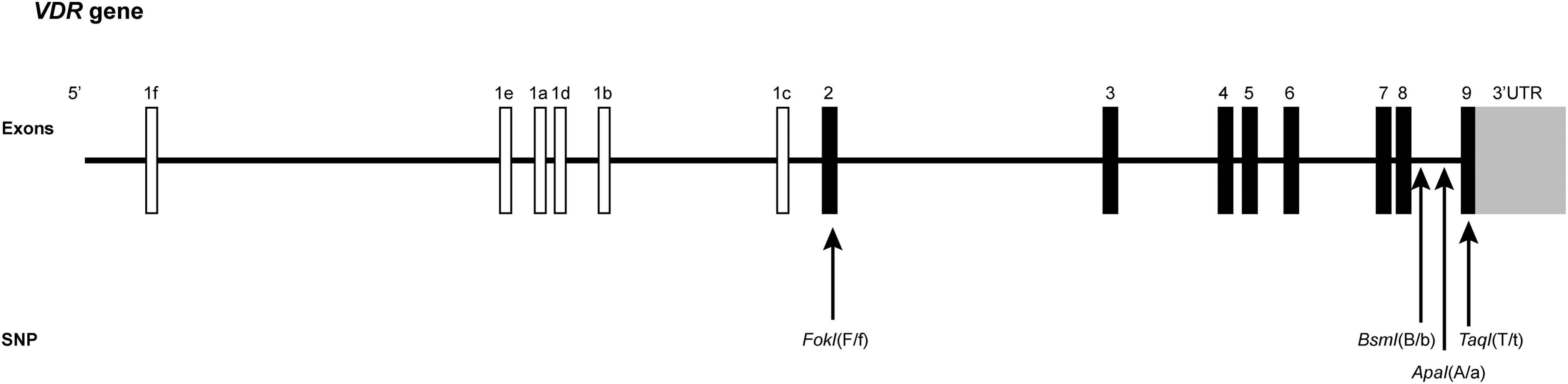
Gene structure of VDR and its four SNPs. White and black charts represent, respectively, the noncoding exon and coding exon. FokI polymorphism is located in exon 2 and provokes a cleavage in T (f) when C (F) is substituted. Both BsmI and ApaI are located in intron 8 and induce restriction site in G (b,a) when A(B) and T(A) are, respectively, substituted. TaqI is in exon 9 and cleaves in C(t) when T is substituted. In the case of absence of cleavage site in restriction enzyme assays, the respective allele is denoted with a capital letter (A for Apa-I, T for Taq-I, F for FokI, and B for BsmI), while the presence of a restriction site is indicated by a lowercase letter (a, t, f, b). SNPs, single nucleotide polymorphisms; VDR, vitamin D receptor.
Materials and Methods
Characteristics of PD patients and healthy controls
A total of 70 subjects participated in this study, conducted at the “Servicio de Neurología del Complejo Universitario Hospitalario de Albacete.” Adhering to the Declaration of Helsinki guidelines, the study received approval from the Ethics Committee (04/2015). Of these participants, 54 were diagnosed with PD, and 16 served as healthy controls (HC), with no neurological or psychiatric history (Table 1). The HC group had a lower mean age (62.38 ± 4.01) compared to the PD patients (69.46 ± 11.10), with a statistical significance of p = 0.0152. Within the PD cohort, ages ranged from 44 to 88 years. Specifically, 23 patients had early-onset PD (≤60 years), with an average age at onset of 48.74 ± 7.92 years and an onset age range of 33-60 years. Conversely, the 29 late-onset PD patients had an average age at onset of 68.55 ± 4.34 years. PD diagnosis was based on primary motor symptoms (rigidity, akinesia, and/or tremor) affecting one or both sides of the body. All participants provided written informed consent.
Characteristics of the Subjects of Study
The characteristics of subjects enrolled in this study, as well as the symptoms and the age of onset in patients with PD.
HC, healthy controls; M, mixed; PD, Parkinson's disease; RA, rigidity and akinesia; SD, standard deviation; T, Tremor; U, unclassified.
Genomic DNA extraction
Genomic DNA (gDNA) was extracted from the peripheral blood samples of both healthy subjects and PD patients for analyzing the VDR gene polymorphisms. The extraction was performed using the GeneJET Whole Blood gDNA Purification kit (K0782), following the manufacturer's instructions. Subsequently, we employed specific restriction enzymes from Thermo Scientific for the analysis: ApaI (FD1414), BsmI (ER0962), TaqI (FD0674), and FokI (FD2144), each being tested in the individual samples.
Restriction enzyme digestion and genotyping of SNPs
DNA fragments of the polymorphisms TaqI (rs731236), ApaI (rs7976091), BsmI (rs1544410), and FokI (rs10735810) were amplified using polymerase chain reaction (PCR) with specific primers. The primer sequences and melting temperatures (Tm) were as follows: ApaI: Forward (FW) 5′-CAACCAAGACTACAAGTACCGCGTCAGTGA-3′ and Reverse (RV) 5′-CACTTCCACAAGGGGCGTTAG-3′, Tm 59°C. BsmI: FW 5′-AGCTGGCCCTGGCACTGACTCTGCTCT-3′ and RV 5′-AACCAGCGGGAAGAGGTCAAGGG-3′, Tm 59°C. FokI: FW 5′-CAGAGTATGGACAGGGAGCAA-3′ and RV 5′-ATGGAAACACCTTGCTTCTTCTCCCTC-3′, Tm 60°C. TaqI: FW 5′-CAGAGTATGGACAGGGAGCAA-3′ and RV 5′-CACTTCGAGCACAAGGGGCGTTAG-3′, Tm 59°C.
The PCR products for ApaI (2000 base pairs [bp]), BsmI (825 bp), FokI (265 bp), and TaqI (501 bp) underwent restriction enzyme digestion for RFLP analysis, followed by gel electrophoresis separation. The alleles and genotypes for each site were designated as ApaI (T and G; TT, GG, TG), BsmI (A and G; AA, GG, AG), TaqI (T and C; TT, CC, TC), and FokI (C and T; CC, TT, CT). RFLP products were visualized under a UV transilluminator, and the sizes estimated using a DNA ladder PLUS™ of 100 bp (Gold Biotechnology, D003). Specific fragment sizes for RFLPs were as follows: ApaI [TT(2000 bp), GG(1700, 300 bp), TG(2000, 1700, 300 bp)], BsmI [AA(825 bp), GG(175, 650 bp), AG(175, 650, 825 bp)], FokI [CC(265 bp), TT(96, 169 bp), CT(96, 169, 265 bp)], and TaqI [TT(501 bp), CC(206,295 bp), TC(206, 295, 506 bp)].
Statistical analysis
Statistical analyses were conducted using SPSS22, Graphpad Prism8 (Version 8.8.2), and Excel software. The distribution of genotypes and alleles in PD patients and controls, as well as the differences in distribution between the two groups, was assessed using the Student's t-test and the Chi-square test. Odds ratios (OR) were calculated from https://www.medcalc.org/calc/odds_ratio.php to determine the association between genetic VDR polymorphisms and the risk of PD, as well as gender. Statistical significance was determined at p < 0.05, and the Bonferroni correction was applied to Tables 3 and 4, confirming significance when p ˂ corrected α (αc).
Limitations of the study
Limitations of this study include a small sample size and incomplete data, such as age, symptoms, and disease duration, which may impact the interpretation of observed results. Additionally, we did not assess the level of Vitamin D, preventing the establishment of an association with the studied SNPs.
Results
Gender differences
Our group of PD patients comprises 35 males and 17 females. While there was no significant difference in the age of disease onset between men and women (Table 2), we observed that in the categorization of the group into two subgroups—early-onset (age ≤60 years) and late-onset (age >60 years) of PD—there were more men than women in each subgroup. Notably, the mean age of PD females (42.86 ± 7.54) was lower than that of PD males (51.36 ± 6.66) in the early onset of PD (p = 0.01). No significant difference was found in the late onset subgroup. Our findings suggest that females seem to develop PD at an earlier age than males.
Early-Onset Parkinson's Disease in Females
The number of patients and the age at onset of PD in each gender group. A significant difference between males and females is observed in the age at onset under 60 years of age (p = 0.01).
Analysis of the VDR polymorphisms in PD patients
In this analytical study, we evaluated the VDR polymorphisms (ApaI, FokI, TaqI, and BsmI) in PD patients. Our findings were similar for ApaI, FokI, and TaqI polymorphisms, where no significant differences were noted in genotype or allele frequencies between PD patients and HC (Table 3). Specifically, the ApaI and FokI genotypes showed similar distributions (ORs of 1.466 and 0.533, respectively) and allele frequencies in both groups. Similarly, the TaqI polymorphism presented comparable genotype distributions (OR = 0.653) and allele frequencies. None of these polymorphisms showed significant associations with the age of disease onset or gender in PD patients.
FokI, BsmI, ApaI y TaqI Restriction Sites in Parkinson's Disease Cases and Controls
A comprehensive analysis of the genotypes and allele distributions of VDR ApaI, TaqI, FokI, and BsmI in PD patients compared to Controls. Statistical significance is indicated with p-values and chi-square (χ2) test results. A Bonferroni correction has been applied to adjust the significance p-value levels for multiple comparisons, with a αc threshold of 0.017. Notable findings are highlighted, including a significant association (*p = 0.004, χ2 = 8.26) in the observed genotypes between PD patients and controls in the VDR BsmI polymorphism, p < αc.
αc, corrected α; VDR, vitamin D receptor.
In contrast, the BsmI polymorphism exhibited a distinct pattern. A significant difference was observed in the BsmI genotypes between PD patients and controls (χ2 = 8.845, p = 0.012), with a notably higher genotype distribution (OR = 5.256, p = 0.0062 for AA+AG vs. GG) and A allele frequency (OR = 3.846, p = 0.0041) in PD patients. However, like the other polymorphisms, BsmI did not show a significant correlation with the age of disease onset. The gender association, though not statistically significant, was more pronounced in the BsmI polymorphism compared to others.
Combined BsmI and FokI polymorphisms
Given the significant association of BsmI genotypes with PD in our study population, we examined combinations of the BsmI SNP with those of FokI, TaqI, or ApaI. No significant findings were observed in the combined genotypes of these two SNPs, except for BsmI-FokI combination (χ2 = 12.50, p = 0.013). Interestingly, the genotype GGCT was significantly more prevalent in healthy subjects (p = 0.001) (Table 4). These results are consistent with the frequency of the BsmI GG genotype (Table 3), which was higher in HC (62.5%) compared to PD patients (21.4%).
Combination of BsmI and FokI Genotypes
Distribution of combined VDR BsmI and FokI genotypes in PD patients and Controls. AACT, AATT, AGTT, and GGTT were not observed in Controls, and they were removed from the contingency table. *p = 0.001, χ2 = 10.671, αc = 0.01 (Bonferroni correction), p < αc.
Discussion
PD is a neurological disorder characterized by the progressive onset of both nonmotor and motor symptoms (Tibar et al., 2018). Although nonmotor symptoms may precede motor symptoms (Kumar et al., 2022), the clinical diagnosis of PD primary relies on motor symptoms, such as tremors, rigidity, postural instability. Within our study population, 33.3% of patients exhibit tremors, 26% show rigidity, and 20% have mixed symptoms, while the rest remain unclassified (Table 1). Except for two individuals, the rest of PD patients have an asymmetrical onset of symptoms either on left or on the right. The average age of PD onset in Spanish patients is around 60 years old, which is consistent with the average age of disease onset in the Western world (Post et al., 2020). Nevertheless, 5.76% of individuals developed PD before the age of 40, with 3.84% of them being females. Patients beneath this age are considered young-onset PD patients (Post et al., 2020), which is a subset of early-onset PD (≤60 years). When the cut-off age is set at 50 years, the percentage of young-onset Parkinson's patients reached 23%.
Certain mutations in PARK genes have been associated with both sporadic and familial PD (Correia Guedes et al., 2020) and may be considered high risk factors for developing early-onset PD. However, genetic mutations account for only 10% of PD cases (Yakhine-Diop et al., 2016), suggesting that most cases results from a combination of genetic susceptibility and environmental factors (González-Polo et al., 2015).
Environmental factors, such as sunlight exposure and/or polymorphisms in VDR gene (Suzuki et al., 2012), can influence the regulation of vitamin D metabolism. Although the relationship between reduced sun exposure, vitamin D levels, and the severity of PD symptoms remains unclear (Peterson et al., 2013), it is evident that the level of 25[OH]D3 is lower in PD patients than in healthy subjects (Gezen-Ak et al., 2017). In our study population, no association was found between ApaI, TaqI, or FokI polymorphisms and PD. The four SNPs were not associated with the age of onset of PD nor with the occurrence of symptoms, such as tremors, akinetic rigidity, or mixed symptoms.
Notably, 50% of patients in the early-onset PD group presented tremor, while only 34% exhibited tremor in the late-onset PD group. Interestingly, lower levels of 25[OH]D3 have been reported in young PD patients with tremor symptoms compared to those with akinetic rigidity (Gezen-Ak et al., 2017). Unfortunately, our study did not measure vitamin D levels in patients' serum to establish an association with the four genetic variants of VDR gene. Gender differences in nonmotor symptoms, such as fatigue, depression, and sleep disorders, have been observed in PD patients (Haaxma et al., 2007). As previously reported (Haaxma et al., 2007), 64% of women experienced tremor, while only 31% of men did. Unlike to this report, the age of onset in females (57.35 years) was younger than in males (60.97 years). This age difference becomes particularly significant in the early-onset group, with women averaging 42.86 years and men 51.31 years.
In contrast to the other SNPs, BsmI was significantly associated with PD in this Spanish cohort (Table 3). This association was further strengthened when BsmI was combined with FokI. Our results suggest that the G allele is protective, with GGCT being the most frequently observed in the control group. In our study, the distribution of VDR BsmI genotypes in HC (AA 6.25%, AG 31.25%, GG 62.5%) was quite similar to that observed in healthy Korean (AA 1.3%, AG 26%, GG 72.7%). Surprisingly, the GG genotype was considered as a risk factor for PD in Korean population (Kim et al., 2005). However, in the Spanish population, the AG genotype for BsmI had the highest frequency. Therefore, the A allele was considered a genetic risk for PD and may double the incidence in men. Furthermore, the A allele was associated with an increased risk of vitamin D deficiency (Rahmadhani et al., 2017), which sustains the association of BsmI polymorphism with PD development.
Conclusions
BsmI genotypes and the A allele were significantly associated with PD, although not with the age of disease onset. In our study, the average age of Parkinson's patients is 60 years. Remarkably, two-thirds of the youngest patients, who presented with tremors, being women. This could be attributed to the fact that women were younger than men in this study. However, SNPs were not significantly associated with PD symptoms or gender. It is important to note that while the BsmI A allele increases the incidence of PD in men, the G allele appears to be protective.
Footnotes
Authors' Contributions
Conceptualization, M.F.G., J.M.F., and S.M.S.Y.-D.; methodology and analysis, S.C.-C., M.R.-A., M.F.G., J.J., and I.C.-N.; writing—original draft preparation, S.M.S.Y.-D.; writing—review and editing, S.C.-C., M.R.-A., I.C.-N., M.F.G., J.J., and J.M.F.; supervision, J.M.F. and S.M.S.Y.-D.; project administration, M.F.G. and J.M.F.; funding acquisition, J.M.F. and M.F.G. All authors have read and agreed to the published version of the article.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by Ethics Committee of clinical investigation 04/2015 del servicio de salud de castilla la mancha (sescam).
Informed Consent Statement
Informed consent was obtained from all subjects involved in this study and written informed consent has been obtained from the patient(s) to publish our results.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by CIBERNED (CB06/05/0041) and was also partially supported by the “Fondo Europeo de Desarrollo Regional” (FEDER) from the European Union. S.C.-C. was supported by FPU fellowships (FPU19/04435) from the Ministerio de Ciencia, Innovación y Universidades, Spain. J.M.F. received research support from the “Instituto de Salud Carlos” III, CIBERNED (CB06/05/0041). S.M.S.Y.-D. was supported by the “Maria Zambrano” Contract (“Plan de recuperación, Transformación y Resilencia” funded by European Union-NextGenerationEU).