
Abstract
Background:
RNA-binding motif proteins (RBMs) have been widely implicated in the tumorigenesis of multiple human cancers but rarely investigated in glioblastoma (GBM).
Methods:
The expression level of RBM47 and its correlation with prognosis of GBM were examined using bioinformatics, quantitative reverse transcription PCR, and Western blot analysis. The colony formation assay and Cell Counting Kit-8 assay were used to determine the biological role of RBM47 in GBM. To measure invasiveness we used the wound healing assay and transwell assay. The regulatory relationship between RBM47 and the epithelial-to-mesenchymal transition (EMT) was examined by Western blot analysis and bioinformatic analysis.
Results:
Through integrative analysis of clinical proteomic and genomic tumor datasets, we found that RBM47 is significantly upregulated in GBM mesenchymal subtype, and its high expression is correlated with poor prognosis. In in vitro biological experiments, we observed a significant inhibitory effect of RBM47 knockdown on colony formation and cell growth using GBM cell lines. Conversely, overexpression of RBM47 restored and accelerated these processes. Moreover, in vitro, wound healing assays demonstrated the role of RBM46 in promoting and cell migration and invasion. Mechanistically, RBM47 enhances invasive capacity through the activation of the EMT program. In RBM47-knockdown cells, the expression levels of Vimentin and CD44 were suppressed, and the level of E-cadherin was increased.
Conclusions:
Taken together our results demonstrate the tumor promoting characteristics of RBM46 and suggest that it could be used both as a therapeutic target and prognostically.
Introduction
Glioblastoma (GBM) remains one of the most aggressive and lethal malignancy in brain tumor, and the median survival following diagnosis is less than 15 months with current therapies (Ohgaki and Kleihues, 2005; Stupp et al., 2009). In the past decade, emerging treatments, such as immunotherapy, and combination therapy, have dramatically improved the outcomes for GBM. However, long-term survivors with 2-5 years are still very low (Stupp et al., 2005), innovative therapies are urgent needed to prolong the median survival. The heterogeneous cellular composition, diffuse invasiveness, fasciculate vasculature, and blood-brain barrier play a special role in GBM biology, which are also account for the resistance and high mortality. Elucidating new insights into these tumors and exploring predictive novel biomarker as well as core molecular mechanisms are indispensable to the improvement of better treatment and prognosis of patients with GBM.
Recent research has shown that epigenetic factors should be included in the creation of new GBM therapeutics (Neftel et al., 2019; Romani et al., 2018). Silencing LSD1 prevents it from interacting with CREBBP as an ATF4 stimulator, slowing the growth of GBM and increasing susceptibility to therapy (Faletti et al., 2021). EZH2 can attach to target gene promoters, inducing methylation and affecting gene expression. Because its overexpression results in a poor prognosis, it can be considered an independent prognostic factor in GBM (Paskeh et al., 2022). RNA-binding protein (RBP) DHX9 is involved in glioma progression and infiltration of TAMs through direct interaction between TCF12 and CSF1 (Liu et al., 2023).
Moreover, the PTEN/PI3K/AKT pathway has been linked to tumor resistance and epithelial-to-mesenchymal transition (EMT) in prostate cancer (Hashemi et al., 2023b). Interestingly, as an integrated molecular pathway, when GBM cells desire to increase their proliferation or induce EMT mechanism, they inhibit or suppress PTEN signaling, whereas PI3K/Akt signaling activation occurs in these processes (Hashemi et al., 2023a). As a result, understanding the significance of the role of molecular interactions in GBM progression is critical.
RNA-binding motif proteins (RBMs) belong to RBPs family, have been clarified to be involved in a series of biological events, including the regulation of RNA editing, splicing, and transcription stability (Bechara et al., 2013; Sutherland et al., 2005). Accumulating investigations have illuminated that RBMs are abnormally expressed in various types of cancers, which demonstrate that RBMs serve a potential role in tumorigenesis and progression (Di Cecilia et al., 2016; Jung et al., 2020; Pavlyukov et al., 2018). RBM47 is another member of RBMs family, which is composed of three RNA recognition motifs and an alanine-rich domain (Maris et al., 2005). RBM47 is ubiquitously expressed in many mouse or human tissues, and the expression level is variously different (Fossat et al., 2016). RBM47 plays an essential role in embryonic function, absence of RBM47 results in reduced litter size and perinatal lethality (Fossat et al., 2016). In addition, RBM47 is a C to U editor, it could pair with APOBEC1 to participate in C to U editing (Fossat et al., 2014).
Moreover, RBM47 is recently identified as an immune brake, RBM47 orchestrates immune-suppression process through promoting the production of interleukin (IL)-10 and inducing Foxp3+ regulator T cell (Wei et al., 2019). RBM47/SNHG5/FOXO3 axis forms a loop and regulates cell autophagy in thyroid cancer cells (Qin et al., 2022). Recently, RBM47 was thought to be a tumor-suppressor gene, it could inhibit cell proliferation and metastasis in breast cancer, colon cancer, thyroid cancer, and lung carcinoma (Kim et al., 2019; Sakurai et al., 2017; Vanharanta et al., 2014). Surprisingly, in nasopharyngeal carcinoma sample, RBM47 acts as an oncogenic gene; it promotes tumorigenesis through multiple pathways (Xu et al., 2021). However, little is known about the role of RBM47 in GBM, as well as its potential involvement in EMT-related mechanism. Here, we analyzed the expression level of RBM47 in normal and GBM tissue through integrative analysis of clinical tumor datasets, which reveal that RBM47 is vastly upregulated in GBM. We have further verified that RBM47 promotes cell proliferation and migration through the activation of EMT process. Our work strongly highlights a novel role of RBM47 in the development of GBM, and RBM47 might be a potent diagnostic marker and therapy target in clinical.
Materials and Methods
Cell lines and cell culture
Human GBM cell lines LN229, T98G, LN18, and human microglia HMC3 cell line were obtained from the American Type Culture Collection, human GBM cell line SF539 was obtained from the cell bank of the Chinese Academy of Science, and human astrocytes HA cell line was obtained from ScienCell. The cells were maintained in Dulbecco's Modified Eagle Medium (DMEM) (Cat. No. 11960044; Gibco) containing 10% fetal bovine serum (Cat. No. 10100147; Gibco), and 1% penicillin and streptomycin (Cat. No. 15070063; Gibco). Cells were maintained in 37°C incubator at 5% CO2.
RNA isolation and quantitative real-time PCR
Total RNA was isolated using the Total RNA Isolation Kit (Takara) according to manufacturer's instructions. cDNA was then obtained by reverse transcription of the RNA using HiScript II Q RT SuperMix kit (Vazyme). quantitative reverse transcription PCR (qRT-PCR) was performed using the cDNA as template. Experiments were performed with three biological replicates using the 7500 Fast Real-Time PCR System (Applied Biosystems). mRNA levels were normalized to β-actin mRNA levels of each sample. The primers used for qRT-PCR (RBM47-F: ATCAGCAATCCTTGGCTCAC; RBM47-R: CCTTGGGATTCCTCTGTTCA. GAPDH-F: GCACCGTCAAGGCTGAGAAC; GAPDH-R: TGGTGAAGACGCCAGTGGA).
Western blot
Total cells and tissues were lysed in the cell buffer containing 1 × protease inhibitor (Roche). The supernatants were collected and resolved on a 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gel, and then transferred to a nitrocellulose filter membrane. After blocking with 5% skimmed milk, the membrane was incubated with the indicated antibodies, including anti-RBM47 antibody (Cat. No. HPA006347; Sigma), anti-Vimentin antibody (Cat. No. 5741; Cell Signaling Technology), anti-CD44 antibody (Cat. No. 37259; Cell Signaling Technology), anti-E-cadherin antibody (Cat. No. 14472; Cell Signaling Technology), anti-Flag antibody (Cat. No. 66008-4-lg; Proteintech), and anti-GAPDH antibody (Cat. No. 60004-1-lg; Proteintech).
Immunohistochemistry
Immunohistochemistry (IHC) was performed as previously described (Luo et al., 2017). The paraffin-embedded tissue sections were deparaffinized with xylene and rehydrated, followed by antigen retrieval. The sections were then treated with 3% hydrogen peroxide to quench the endogenous peroxidase activity, followed by incubation with a primary antibody overnight at 4°C. After incubation with secondary antibodies for 1 h at room temperature, chromogenic immunolocalization was conducted through diaminobenzidine. All sections were counterstained with hematoxylin. IHC scores were calculated as the product based on the staining intensity and the percentage of cells stained.
Wound healing assay
GBM cells were seeded into the six-well plate until they reached complete monolayer confluence. Mitomycin C was supplemented before creating a defined cell-free gap. After washing twice with Hank's Balanced Salt Solution to carefully remove mitomycin C and detached cells, cells were then incubated in DMEM without serum, and images were captured at 0 and 48 h, using an inverted microscope (Olympus). The migration ability of the cells was calculated as the area measured in pixels between the edges of the scratching at the indicated time point relative to that of the starting time point.
Matrigel coating transwell assay
Cell transwell (8.0 μm pore size membrane; Corning) was first coated with matrigel (BD Bioscience) at a dilution of 3.3 ng/mL in a medium without serum. Subsequently, 5 × 104 cells, previously deprived of serum for 24 h, were seeded on the matrigel in the upper transwell chamber in a serum-free medium. As a chemoattractant, 20% fetal bovine serum was placed in the lower chamber. After 48 h, nonmotile cells at the top of the filter were removed, and the cells in the bottom chamber were fixed with methanol and stained with crystal violet and counted using immunofluorescence microscopy. Results represent the average number of cells in five fields.
Cell proliferation
For the cell growth, GBM cells were plated in a triplicate manner in a 96-well plate and were tested every 24, 48, and 72 h. Cell number and viability were determined using Cell Counting Kit-8 (CCK8) according to the manufacturer's protocol. Optical density of the wells was measured at 450 nm using a microplate reader.
Colony formation assay
For colony formation assay, 3 × 103 cells were seeded in a six-well plate. After growing for 7 days, cells were fixed in 4% paraformaldehyde for 20 min at room temperature and stained with 0.5% crystal violet solution for 15 min.
Knockdown and overexpression assay
The knockdown lentiviruses expressing shRNAs targeting RBM47 (shRBM47-1 and shRBM47-2) and negative control shRNA (shControl) were constructed with PLKO.1 lentiviral plasmid. The overexpression lentivirus expressing Flag-RBM47 and Flag-Vector were constructed with pcDNA3.1 vector. All lentiviruses were packaged in 293T cells and selected with puromycin. All shRNA oligos were commercially synthesized (Shanghai GenePharma Co., Ltd.), and the RBM47 sequence were cloned from GBM cell lines. The primer sequence and shRNA sequence (shRBM47-1-F: CCGGAAGCCTCATGATGGACTTTGACTCGAGTCAAAGTCCATCATGAGGCTTTTTTTG; shRBM47-1-R: AATTCAAAAAAAGCCTCATGATGGACTTTGACTCGAGTCAAAGTCCATCATGAGGCTT. shRBM47-2-F: CCGGAACGCCCAATAACTCCAGTATCTCGAGATACTGGAGTTATTGGGCGTTTTTTTG; shRBM47-2-R: AATTCAAAAAAACGCCCAATAACTCCAGTATCTCGAGATACTGGAGTTATTGGGCGTT).
Bioinformatics analysis
To comprehensively explore the expression pattern and prognostic implications of RBM47 in GBM, preprocessed RNA-seq and corresponding clinical data from CGGA, REMBRANDT, and TCGA cohort were downloaded from GlioVis data portal. For GBM subtyping, the most recent three-subtypes classification scheme was used. Differential RBM47 mRNA expression in GBM subtype was determined by nonparametric Mann-Whitney test.
For CPTAC-GBM proteomics data, unshared log-ratio data were normalized using the median centering method across total proteins to correct sample-loading differences. Differential RBM47 protein expression in GBM and normal control was determined by unpaired t test. Survival analysis was conducted via the “survminer” package. GBM patients were categorized into high and low RBM47 expression group using the optimal cutoff points determined using surv_cutpoint function, and survival curves were based on Kaplan-Meier estimates. The Pearson correlation between RBM47 mRNA and protein expression level with EMT-related marker genes was calculated. Statistical analyses were performed using the R statistical software.
Statistical analysis
Data were first evaluated using one-way analysis of variance (ANOVA). Multiple comparisons between the treatment groups and controls were performed using Dunnett's least significant difference (LSD) test. All statistical tests were performed with Prism software (version 9; Inc., La Jolla, CA, USA). p < 0.05 was considered to indicate a statistically significant difference.
Results
RBM47 is highly expressed in GBM and correlated with a poor prognosis
We first checked the protein expression level of RBM47 between normal and GBM tissue from the Clinical Proteomic Tumor Analysis Consortium (CPTAC) dataset, and we observed that the protein level of RBM47 is vastly upregulated in GBM (Fig. 1a). Consistently, we found RBM47 was highly expressed in GBM samples (n = 12) compared with normal brain tissue samples (n = 9) from the IHC assay (Fig. 1b). Our preliminary data reveal that RBM47 might be a potent oncogenic marker in GBM. Next, we analyzed the expression level of RBM47 between distinct GBM subtypes. Aberrations and gene expression of EGFR, NF1, and PDGFRA/IDH1 each define the classical, mesenchymal, and proneural subtypes, respectively (Verhaak et al., 2010). We found RBM47 expression in mesenchymal subtype were significantly upregulated than in other types (Fig. 1c), suggesting that RBM47 could contribute as a specific marker for the mesenchymal subtype.

In addition, we observed that the protein and mRNA level of RBM47 was variously improved compared with normal astrocytes HA cell line except for human HMC3 cell line, with human GBM cell lines LN229, T98G, and LN18 having the strongest expression (Fig. 1d, e). Kaplan-Meier survival curves revealed that patients with GBM who had a low expression of RBM47 had longer overall survival (Fig. 1f). Collectively, these data suggest that RBM47 might be an oncogenic factor and could be applied in a diagnostic marker for GBM.
RBM47 is positively associated with cell proliferation in human GBM cell lines
To further investigate the oncogenic function of RBM47 in GBM, we first knocked down the expression of RBM47 in human GBM cell lines LN229 and T98G using shRNAs (Fig. 2a). Cell number was sharply reduced in RBM47 knockdown cell lines after 48 and 72 h (Fig. 2b), and we observed a significant inhibitory effect of RBM47 knockdown on colony formation on both cell lines compared with the control settings (Fig. 2c). To further verify the function of RBM47 in cell proliferation in GBM cell lines, we overexpressed RBM47 in GBM cell lines, LN229 and T98G (Fig. 2d). As expected, cell number was increased in RBM47 overexpression cell lines after 48 and 72 h (Fig. 2e), and the number of colony formation was also increased in RBM47 overexpression cell lines (Fig. 2f). These observations suggested that RBM47 promotes cell proliferation in GBM cell lines.

RBM47 promotes cell migration and invasion
Cell migration is a hallmark of wound repair, cancer invasion, and metastasis. To investigate the function of RBM47 in cell migration, we designed a wound healing assay to simulate the cell migration and movement (Grada et al., 2017). The GBM cell lines were cultured in a two-dimensional confluent monolayer; then, the monolayer was scraped into a linear thin “wound” and subsequently capturing the images of the cells filling the thin gap. Strikingly, after 48 h, the gap was obviously wider in RBM47 knockdown cell lines and narrower in RBM47 overexpression cell lines compared with the control settings (Fig. 3a, b). These suggested that the expression level of RBM47 is positively correlated with cell migration.
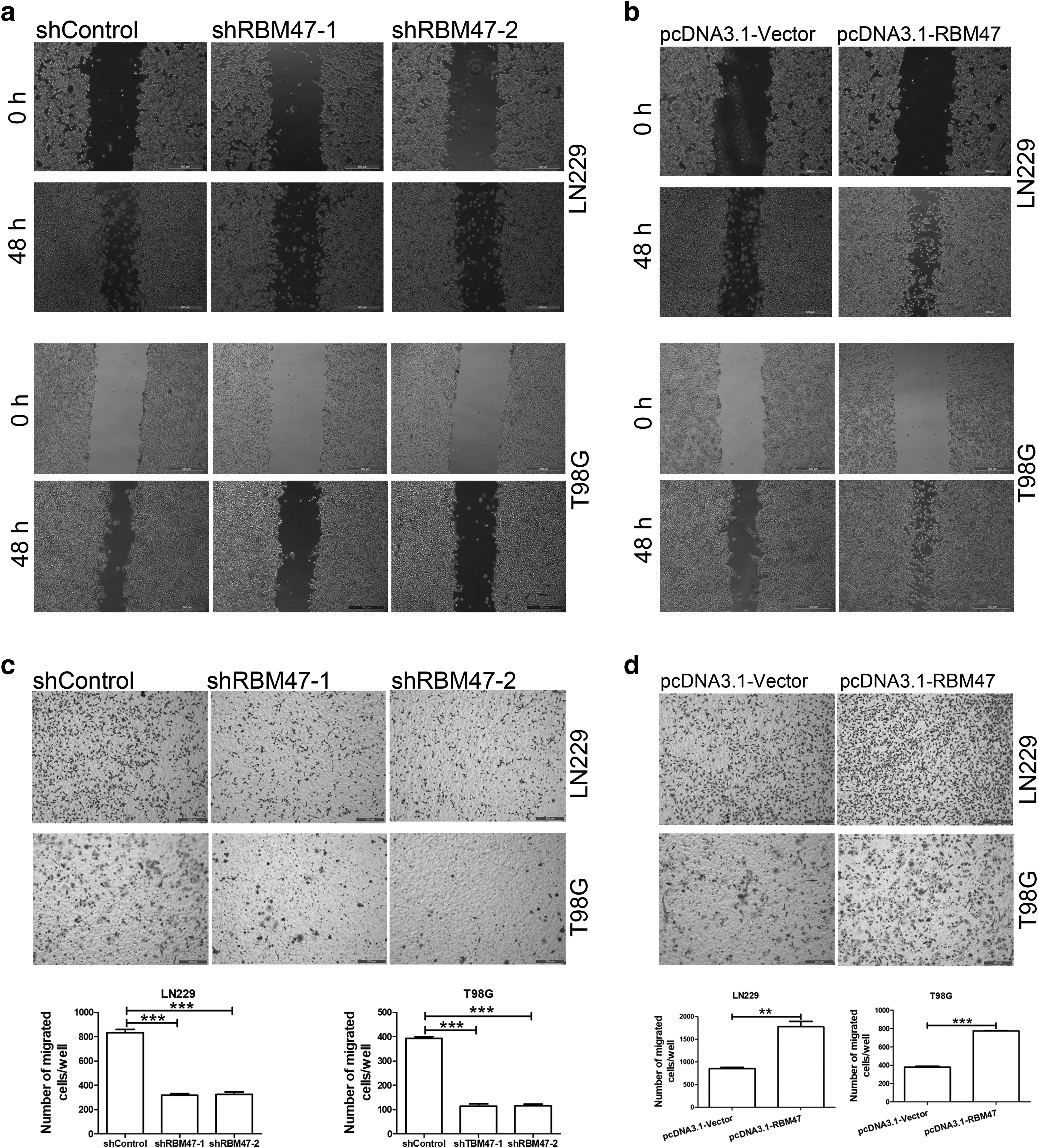
To further clarify the oncogenic role of RBM47 in GBM cell line, we performed a matrigel-coating experiment to imitate cell invasion and metastasis (Moroi et al., 1992). The invasive cells migrated through the row of matrigel-coated microgaps by breaking down the matrigel network. Our data showed that the number of migrated cells were increased in RBM47 upregulated cell lines and reduced in RBM47 downregulated cell lines compared with the control settings (Fig. 3c, d), which suggested that RBM47 could promote cell migration, invasion, and metastasis.
RBM47 promotes cell invasion and metastasis through activation the EMT process
To further investigate the role of RBM47 in cell invasion and metastasis, we had checked the effect of RBM47 in EMT process. During cancer progression, epithelial tumor cells undergo EMT, leading to loss of epithelial markers (E-cadherin), changes in cell polarity and intercellular junctions and increase of mesenchymal markers (fibronectin and vimentin) (Brabletz et al., 2018; Hugo et al., 2007; Kalluri, 2009; Zeisberg and Neilson, 2009). Here, we first checked the expression level of EMT-related marker genes from CGGA, REMBRANDT, TCGA, and CPTAC datasets. As we expected, RBM47 exhibits strong positive linear correlation with Vimentin and CD44, and weak negative correlation with E-cadherin (CDH1) (Fig. 4a).
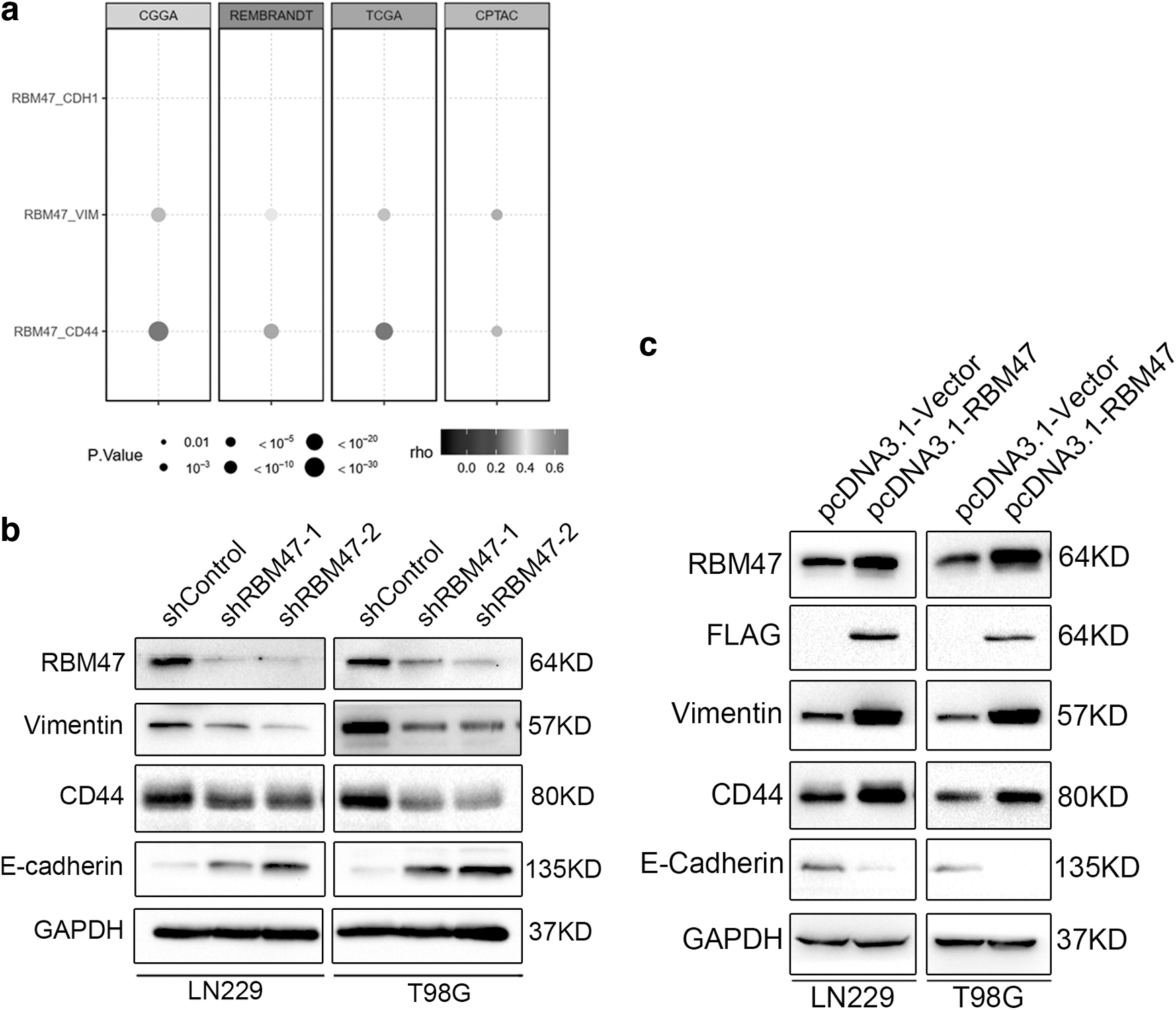
Next, we further checked the function of RBM47 in EMT process through knockdown and overexpression assay. In line with datasets, mesenchymal markers, including vimentin and CD44, were downregulated in RBM47 knockdown cell lines, and epithelial marker, including E-cadherin was upregulated in RBM47 knockdown cell lines (Fig. 4b). While in the case of RBM47 overexpression, these proteins' expression levels were exactly opposite (Fig. 4c). Taken together, RBM47 was an oncogenic gene in GBM and played an essential role in EMT process.
Discussion and Conclusion
Accumulating reports have demonstrated a close link between RBM47 and tumorigenesis. Some recent data have verified the tumor suppression function of RBM47 in breast, colon cancer, and lung cancer (Kim et al., 2019; Sakurai et al., 2017; Vanharanta et al., 2014), which is completely contrary to our point. In addition, the oncogenic effect of RBM47 in nasopharyngeal carcinoma have been verified (Xu et al., 2021), which is consistent with our point. We suspected that the oncogenic or anti-tumorigenic role of RBM47 might be distinct between tissues. Previous reports have revealed that RBM47 regulates thousands of different mRNA, including mRNA transcription, splicing, and stability, and these processes are closely associated with tumorigenesis and homeostasis.
The mechanisms that RBM47 affect tumor progression have been reported in many carcinomas. In lung adenocarcinoma, RBM47 inhibits Nrf2 activity to suppress tumor growth (Sakurai et al., 2017), and RBM47 stabilizes DKK1 and Wnt signal pathway to retard cell proliferation and alleviate tumor progression in breast cancer (Vanharanta et al., 2014). However, in nasopharyngeal carcinoma, RBM47 binds to the promoter of BCAT1 known as an oncogene to promote the transcription and tumorigenesis (Tönjes et al., 2013; Xu et al., 2021). Proteins including RBM47 are specifically or variously expressed in many tissues or organs, which might account for the double-edged role of RBM47 in tumorigenesis. Up to now, the specific regulation mechanisms of RBM47 in the occurrence and development of GBM remain unknown, and more experiments should be conducted to elucidate the precise regulation of RBM47 in cell proliferation in GBM.
Increasing evidence demonstrates the role of EMT in mediating drug resistance in different types of cancer cells. Cisplatin is a platinum chemical that has remarkable antitumor action against a variety of tumor types. Previous research has shown that cisplatin can induce EMT and promote resistance in cancer cells (Ashrafizadeh et al., 2020). Other frequent chemotherapy medications, such as paclitaxel and docetaxel, are also associated with EMT (Ashrafizadeh et al., 2021). Many cellular processes and pathways may synergistically induce EMT and promote chemoresistance in GBM. STAT3, as an upstream mediator of EMT, has been shown to promote EMT-mediated metastasis in GBM, and the interplay between STAT3 and EMT can influence drug-resistance in cancer cells (Sadrkhanloo et al., 2022).
Previous study proved that silencing of LncRNA-H19 decreases chemoresistance of human glioma cells to temozolomide (TMZ) by suppressing EMT via the Wnt/β-Catenin pathway (Jia et al., 2018). CD90 low glioma-associated mesenchymal stem cells promote TMZ resistance by activating FOXS1-mediated EMT in glioma cells (Xue et al., 2021). In addition, because autocrine transforming growth factor-β-signaling increases glioma proliferation, invasion, EMT, and treatment resistance, this complex regulatory network could be used as a therapeutic target for GBM. Thus, inhibition of EMT-induced drug resistance can be evaluated in further studies.
Recently, the effects of RBM47 in immune regulation have been identified, just as mentioned above; RBM47 exerts immune-suppression function through promoting the IL-10 production and inducing Foxp3+ regulatory T cells (Wei et al., 2019). However, a recent article has clarified that RBM47 could stabilize IFNAR1 mRNA to potentiate host antiviral activity (Wang et al., 2021). The role of RBM47 in immune response is controversial, just as in tumorigenesis. GBM microenvironment is highly heterogeneous filled with tumor-associated macrophage and other immune-regulated cells; we suspected that RBM47 might exert an immune-suppression function in GBM through promoting the polarization of TAM. Our data have showed that RBM47 could upregulate the level of CD44, a negative regulator of immune response to activation of EMT process. We will further explore the specific regulation mechanisms of RBM47 in GBM microenvironment.
In conclusion, our data have illuminated that RBM47 is vastly upregulated in GBM samples, and RBM47 is positively correlated with cell proliferation, invasion, and migration as well as EMT process. Our findings provide insights into understanding the important role of RBM47 in the development of GBM, which could be potentially used as a novel clinical marker and therapeutic target in further investigation.
Footnotes
Authors' Contributions
T.-Z.Z. conceived and designed the experiments. Y.-Q.X. and T.-Z.Z. performed all the experiments. All authors read and approved the final version of the article.
Ethical Approval
This work is not research involving human participants and/or animals.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was financed by the National Natural Science Foundation of China (project no. 81603448).