
Abstract
Objective:
To examine if METTL5 promotes the proliferation of nonsmall cell lung cancer (NSCLC) cells by interacting with IGF2BP3.
Methods:
The expression patterns of METTL5 and IGF2BP3 in NSCLC tissues, their relationship with survival rate, and their correlation were analyzed using bioinformatics and clinical sample analyses. The effects of METTL5 overexpression and IGF2BP3 knockdown, as well as those of METTL5 knockdown and IGF2BP3 overexpression, on the proliferation of NSCLC cells were analyzed by transfecting appropriate constructs. The interaction between METTL5 and IGF2BP3 was verified using the co-immunoprecipitation (Co-IP) assay. The in vivo effects of METTL5 and IGF2BP3 on NSCLC growth were analyzed using the tumor-bearing nude mouse model.
Results:
METTL5 and IGF2BP3 expression levels were positively correlated and were associated with poor clinical prognosis. The METTL5 and IGF2BP3 expression levels were upregulated in the clinical NSCLC samples. IGF2BP3 expression did not affect METTL5 expression but was regulated by METTL5. IGF2BP3 overexpression mitigated the METTL5 knockdown-induced impaired cell proliferation. Meanwhile, IGF2BP3 knockdown suppressed METTL5-mediated NSCLC cell proliferation. The Co-IP assay results revealed the interaction between METTL5 and IGF2BP3 in NSCLC cells. IGF2BP3 knockdown suppressed tumor growth, whereas IGF2BP3 overexpression enhanced tumor volume and quality.
Conclusion:
METTL5 induces NSCLC cell proliferation by interacting with IGF2BP3. Thus, METTL5 is a potential biomarker and a therapeutic target for NSCLC.
Introduction
Approximately 2.2 million new cases of nonsmall cell lung cancer (NSCLC) are reported each year worldwide. Lung cancer accounts for 11.4% of all tumors and is the second most common tumor in 2020 (Sung et al., 2021). Furthermore, lung cancer is the most common cause of cancer-related deaths, accounting for more than 1.7 million deaths (Sung et al., 2021). The survival rate of patients with stage I/II NSCLC is high. However, limited numbers of patients are diagnosed with stage I/II NSCLC owing to the obscure early symptoms and a lack of awareness of the need for physical examination (Zhou et al., 2022). The survival rates among patients with stage III and stage IV NSCLC are 20% and 1%, respectively (Caini et al., 2022). The mechanism underlying NSCLC cell proliferation has not been elucidated.
Methyltransferase like protein 5 (METTL5), a methyltransferase-like protein containing 267 amino acids, is involved in RNA modification (Turkalj and Vissers, 2022), especially in N6-methyladenosine (m6A) modification (Wang L et al., 2022). Although the function and mechanism of METTL5 protein are unclear, it is reported to be involved in the pathogenesis of various cancers (Zheng J et al., 2022). In gastric and renal cancer tissues, METTL5 expression is upregulated, indicates pathological progression, and predicts mortality (Wang Z et al., 2021) (Wei et al., 2022). METTL5 is also involved in regulating the proliferation of breast (Rong et al., 2020) and pancreatic cancer cells (Huang et al., 2022). However, the expression patterns, functions, and mechanisms of METTL5 in NSCLC have not been elucidated.
Similar to METTL5, Insulin like growth factor II mRNA binding protein 3 (IGF2BP3) is an m6A reader and an RNA-binding protein that regulates mRNA stability and translation (Yang et al., 2020). IGF2BP3, which binds to mRNA and regulates its trafficking and stability, is involved in human embryonic development and cell proliferation and transformation. In bladder (Huang et al., 2020), nasopharyngeal (Du et al., 2022), and prostate cancers (Yu et al., 2022), IGF2BP3 exerts an oncogenic effect. Both IGF2BP3 and METTL5 regulate RNA methylation in NSCLC (Zhang Z et al., 2021). However, the regulatory effect of IGF2BP3 on METTL5 in NSCLC is unclear.
This study analyzed the effect of METTL5 and IGF2BP3 on NSCLC and the mechanism through which they regulate NSCLC cell proliferation. Thus, this study aimed to provide a new theoretical basis for the diagnosis and treatment of NSCLC.
Materials and Methods
Bioinformatics methods
NSCLC datasets in The Cancer Genome Atlas database were analyzed using the Gene Expression Profiling Interactive Analysis tool. Differential analysis was performed with the data of 483 tumor tissues and 347 nontumorous tissues. The correlation between METTL5 and IGF2BP3 expression levels in NSCLC tissues was examined using the Pearson test. Kaplan-Meier survival analysis was performed using the data of 478 cases.
Clinical samples
In total, the samples of 83 NSCLC cases were collected from Tongjiang County People’s Hospital. Among these, 52 and 47 were METTL5-positive and IGF2BP3-positive cases, respectively. The clinical characteristics of patients are shown in Table 1 and include gender, smoking history, staging, T stage, N stage, and M stage. The patients were classified into METTL5-positive, METTL5-negative, IGF2BP3-positive, and IGF2BP3-negative groups. The significant differences in the clinical characteristics between the METTL5-positive and METTL5-negative groups, as well as between the IGF2BP3-positive and IGF2BP3-negative groups, were analyzed using the chi-square test. All patients agreed to be included in the study. This study was approved by the ethics committee of Tongjiang County People’s Hospital (No. L0008).
Clinical Characteristics of Patients with Nonsmall Cell Lung Cancer
Bold value (P<0.05) indicates statistical significant. NA, not applicable.
Cell culture
Human healthy lung cells (BEAS-2B cells) (MY-K5364, EK-Bioscience, China) and NSCLC cells [H1650 (SY4243, EK-Bioscience), H2023 (CM8504X, CMBIO, USA), A549 (MY-K6354, EK-Bioscience), H1437 (MY-K5687, EK-Bioscience), and H1299 cells (XY-0907, XYBio, China)] were maintained in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco, USA) supplemented with 10% fetal bovine serum. The cells were cultured in a 5% CO2 incubator at 37°C and 95% humidity.
Cell transfection
H1437 and H1299 cells were selected as they exhibited upregulated expression levels of METTL5 and IGF2BP3. pcDNA3.1-METTL5, pcDNA3.1-IGF2BP3, si-METTL5, si-IGF2BP3, and corresponding negative control (NC)/si-NC were purchased from GenePharma Co., Ltd. (China). The reagents were prepared as follows: reagent A, 50 pmol (0.67 μg) plasmid diluted in 25 μL serum-free DMEM; reagent B, 1 μL EntransterTM-R4000 (Engreen) and 24 μL serum-free DMEM mixed for 25 min. Reagent A (25 μL) and reagent B (25 μL) were mixed thoroughly (aspirated 10 times using a pipette) and left undisturbed for 15 min to obtain the transfection complex. Cells in 0.45 mL complete medium were transfected with 50 μL of transfection complex. NC/si-NC plasmids were used as control.
Quantitative real-time polymerase chain reaction (qRT-PCR) analysis
Cells or tissues were transferred to 1.5 mL EP tubes and incubated with 1 mL TRIzol on ice. Next, the samples were incubated with 0.2 mL of chloroform for every 1 mL of TRIzol, shaken vigorously for 15 s, incubated at 25°C for 3 min, and centrifuged at 12,000 g and 4°C for 15 min. The upper aqueous phase was transferred to a new tube and incubated with isopropanol to precipitate the RNA. The samples were centrifuged at 12,000 g and 4°C for 15 min. After discarding the supernatant, the RNA pellet was washed with 75% ethanol (at least 1 mL for every 1 mL of TRIzo1). The samples were centrifuged at 7500 g and 4°C for 5 min. The RNA pellet was allowed to dry at room temperature for approximately 3-5 min and dissolved in 25 µL of RNase-free water. The mRNA was reverse-transcribed into complementary DNA (cDNA) using the PrimeScript RT Kit (#DRR037A, Takara, Japan) and miRNeasy Mini Kit (#217004, Qiagen). qRT-PCR analysis was performed using SYBR Green reagent (#A2203XL, Yaji, China) and TaqMan miRNA (KMD122014, KEMOBio, China). The mRNA expression levels of METTL5 and IGF2BP3 were normalized to those of GAPDH using the 2 −ΔΔCt method. The following primers were used: METTL5, 5′-TTGGGTTAGCCGGGAGATCCT-3′ (forward) and 5′-CCACATCCTAGATCTGCAACGA-3′(reverse); IGF2BP3, 5′-AGACTGGTGGATGCGTTTGG-3′ (forward) and 5′-TTGTTCCCCTCGTCTTCTCG-3′ (reverse); GAPDH, 5′-GCACCGTCAAGGCTGAGAAC-3′ (forward) and 5′-TGGTGAAGACGCCAGTGGA-3′ (reverse).
Western blotting analysis
Cells or tissues were lysed using a mixture of 100 μL lysis buffer, 1 μL protease inhibitor, and 1 μL phenylmethylsulfonyl fluoride after removing the culture medium and washing. After centrifuging at 12,000 g and 4°C for 15 min, the supernatant was transferred to a new 1.5 mL tube. The protein concentration in 2.5 μL of sample in 22.5 μL of triple distilled water was measured using the bicinchoninic acid method. The proteins were resolved using sodium dodecyl sulfate-polyacrylamide gel electrophoresis with an 8-12% gel. The gel was cut according to the marker instructions and the position of the target band and soaked in transfer buffer for 15 min, and then transferred to a PVDF membrane for 90 min. After washing, the membrane was blocked with 5 mL of milk powder blocking solution overnight at 4°C with gentle shaking. Next, the membrane was incubated with rabbit antiprimary antibodies against METTL5 (1:2000, PA5-115571) or IGF2BP3 (1:2000, MA5-27481, ThermoFisher, USA) at 4°C overnight, followed by incubation with secondary antibody (1:2000, ab6721) for 2 h at 37°C. Immunoreactive signals were visualized using the enhanced chemiluminescence kit (Amersham Biotech) and IPP6.0. The relative expression level was determined based on the expression levels of the internal reference GAPDH.
Cell counting kit-8 (CCK-8) assay
Cell suspension (5 × 104/mL) (100 μL) was seeded in 96-well plates. After 1, 2, 3, 4, 5, and 6 days, the cells were incubated with CCK-8 solution (10 μL) on an orbital shaker for 1 min at 37°C to ensure uniform mixing. The incubation was continued for 2 h to allow dehydrogenation reaction. The optical density values at 450 nm wavelength were determined using a LEx808 microplate reader (25-315S, Lonza, USA).
Clone formation assay
After digestion and resuspension, 200 cells were evenly seeded in the medium and cultured under the previously mentioned conditions for 14 days. The medium was replaced once every 2-3 days until macroscopic colonies were obtained. The culture supernatant was discarded and carefully washed twice with phosphate-buffered saline (PBS). The colonies were fixed using 4% paraformaldehyde for 15 min. Next, the colonies were stained with Giemsa for 20 min. The samples were slowly washed to remove the staining solution and air dried. The plate was inverted, and a gridded transparent film was overlaid to count the number of colonies (with >20 cells).
Co-immunoprecipitation (Co-IP)
After removing the medium, the cells were washed thrice with PBS and lysed with an appropriate amount of lysis buffer for 15 min on ice. The lysate was centrifuged at 6000 g for 15 min to remove the precipitate. The supernatant was divided into three groups [positive control group, directly frozen for further analysis; negative control group, incubated with IgG; experimental group, incubated with anti-METTL5 (1:1000, PA5-58301) and anti-IGF2BP3 (1:1000, MA5-27481), ThermoFisher, USA] antibodies overnight at 4°C. Next, the samples were incubated with 40 μL of magnetic beads with rotation at 4°C for 1 h. The magnetic beads were washed with precooled PBS five times and incubated with protein loading buffer. The samples were subjected to western blotting to detect the target proteins.
Xenotransplantation assay
Male BALB/C nude mice (Charles River Co., Ltd, China) aged 4 weeks were used to evaluate cell tumorigenesis. The mice were housed under the following conditions: temperature, 24°C ± 1°C; relative humidity, 60% ± 5%. Cells transfected with pcDNA3.1-METTL5 and si-IGF2BP3 (5 × 106 cells) were resuspended in 200 μL PBS. The transfected cells were subcutaneously injected into the axillary region of nude mice. After 4 weeks, the mice were euthanized and the tumors were harvested and weighed. Animals were euthanized when the diameter of the tumor reached >2 cm, which was not observed in this study. The study was approved by the ethical committee of Tongjiang County People’s Hospital (No. L0008) and followed the guidelines for animal welfare and ethical considerations.
Statistical analysis
Data are expressed as mean ± standard deviation. Means between multiple groups were compared using one-way analysis of variance, followed by Tukey’s multiple comparison tests (GraphPad Prism version 7.0), whereas those between two groups were compared using the T test. The correlation between METTL5 and IGF2BP3 expression levels in NSCLC tissues was examined using the Pearson test. Differences were considered significant at p < 0.05.
Results
METTL5 in NSCLC and its relationship with IGF2BP3
METTL5 was expressed in both cancerous and noncancerous tissues. However, METTL5 expression was upregulated in most tumors, including NSCLC (Fig. 1A). IGF2BP3 was not expressed in noncancerous tissues but was markedly upregulated in cancerous tissues (Fig. 1B), especially in lung cancer (Fig. 1C). Additionally, METTL5 expression was positively correlated with IGF2BP3 expression in NSCLC (r = 0.300, p < 0.001) (Fig. 1D). Based on the median METTL5 expression level, 478 patients were divided into the low-METTL5 and high-METTL5 groups (n = 239). The survival rate in the high-METTL5 group was lower than that in the low-METTL5 group (p < 0.001) (Fig. 1E). The upregulation of IGF2BP3 predicted decreased survival rates (Fig. 1F). METTL5 and IGF2BP3 expression levels were positively correlated in NSCLC and were associated with poor clinical prognosis.
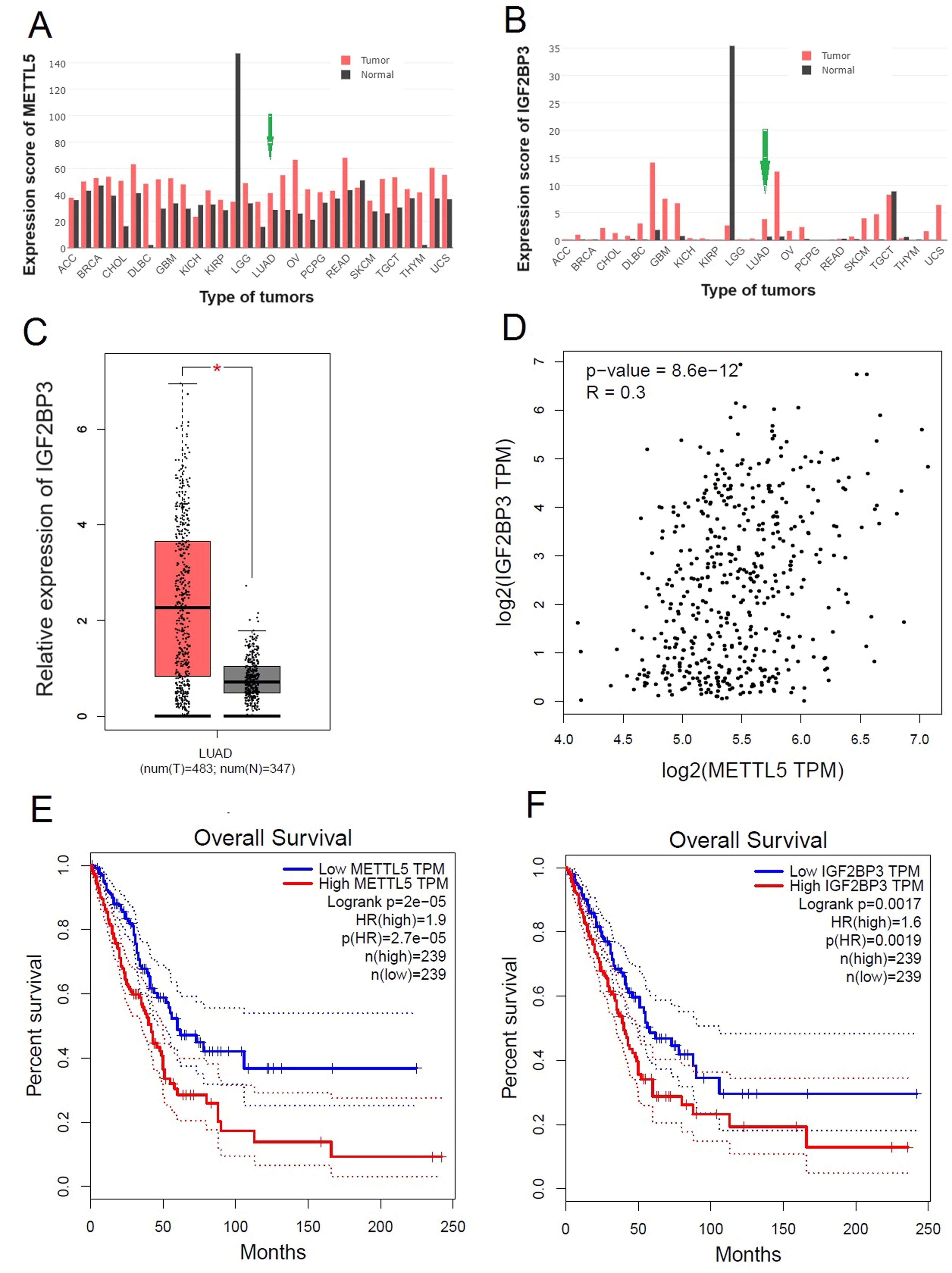
Correlation between METTL5 and IGF2BP3 expression levels in NSCLC.
Clinical significance of METTL5 and IGF2BP3
In total, 83 NSCLC cases were classified into METTL5-positive, METTL5-negative, IGF2BP3-positive, and IGF2BP3-negative groups. The clinical characteristics of patients are shown in Table 1. METTL5 and IGF2BP3 expression levels were significantly correlated with the tumor stages (p < 0.0001 for METTL5; p = 0.0284 for IGF2BP3). qRT-PCR analysis revealed that the METTL5 and IGF2BP3 mRNA levels were upregulated in NSCLC tissues (Fig. 2A, B). Consistent with the bioinformatics analysis results, the METTL5 and IGF2BP3 mRNA expression levels were positively correlated (R2 = 0.5789, p < 0.001) (Fig. 2C). Additionally, the protein levels of METTL5 and IGF2BP3 were upregulated in NSCLC tissues (Fig. 2D, E). These findings validated the results of the bioinformatics analysis at the clinical level.
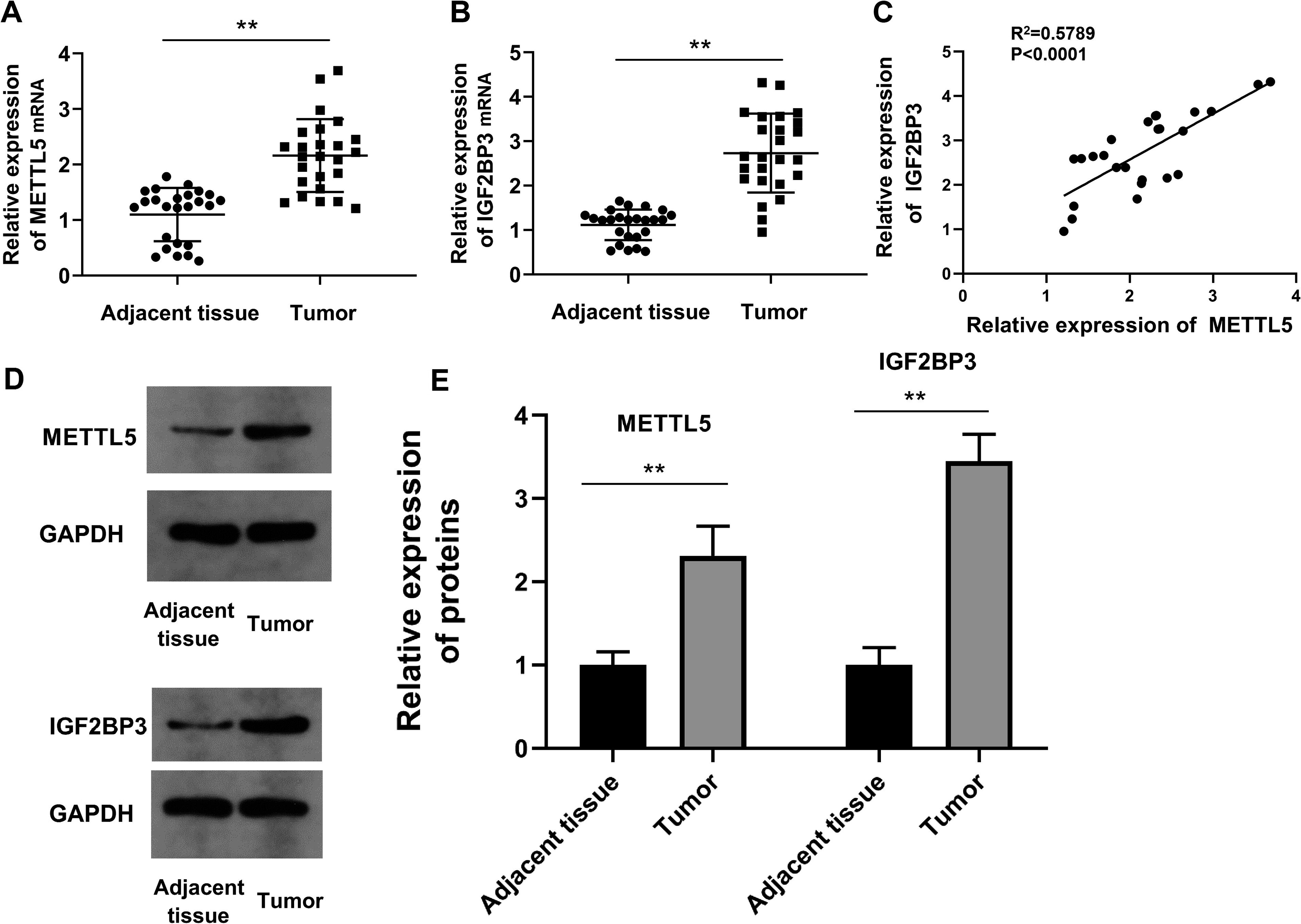
Clinical significance of METTL5 and IGF2BP3.
METTL5 and IGF2BP3 are upregulated in NSCLC cells
Next, the expression patterns of METTL5 and IGF2BP3 were examined in NSCLC cell lines. Lung epithelial cells (BEAS-2B cells) were used as a control. The METTL5 and IGF2BP3 mRNA levels were upregulated in NSCLC cell lines with the highest levels observed in H1473 and H1299 cells (Fig. 3A, B). Therefore, H1473 and H1299 cells were selected for follow-up studies. To evaluate the functions of METTL5 and IGF2BP3 in NSCLC, METTL5 and IGF2BP3 were knocked down or overexpressed in H1473 and H1229 cells. The transfection efficiency was confirmed at both the mRNA and protein levels (Fig. 3C, G, E, I, K, and L). The modulation of METTL5 expression altered the IGF2BP3 mRNA and protein levels (Fig. 3D, H, K, and L). However, the mRNA or protein levels of METTL5 were not altered upon modulation of IGF2BP3 expression (Fig. 3F, J, K, and L). These results confirmed the upregulation of METTL5 and IGF2BP3 in NSCLC at the cellular level and established the basis for subsequent functional verification experiments.
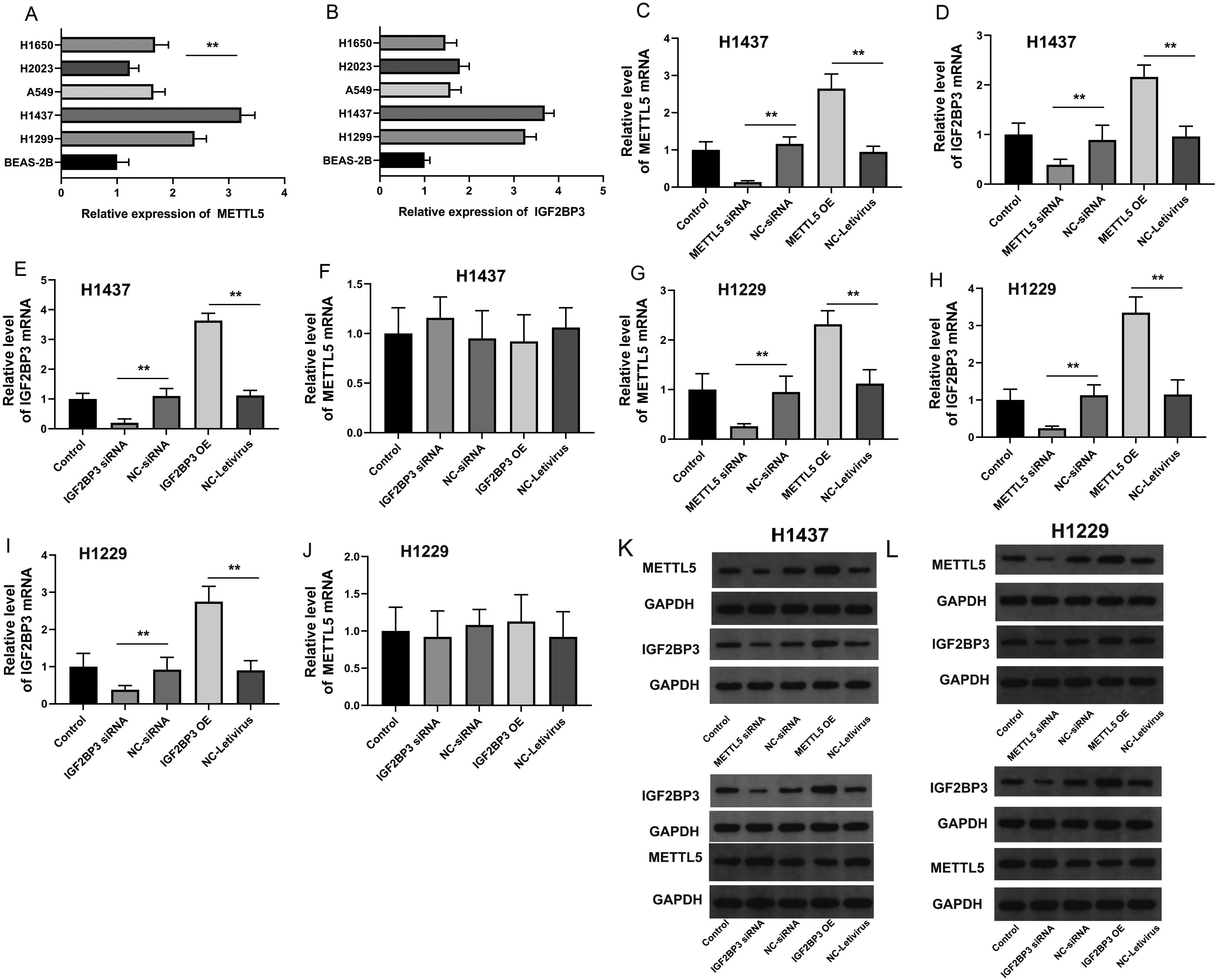
METTL5 and IGF2BP3 are upregulated in NSCLC cells.
METTL5 regulates the proliferation of NSCLC cells through IGF2BP3
To analyze the role of METTL5 and IGF2BP3 in NSCLC cell proliferation, H1473 cells were divided into the following three groups: control, si-METTL5, and si-METTL5 + IGF2BP3 groups. Transfection with si-METTL5 decreased cell viability. The viability of the si-METTL5 + IGF2BP3 group was higher than that of the si-METTL5 group (p < 0.05) (Fig. 4A). Next, this study examined if METTL5 affected the proliferation of NSCLC cells through IGF2BP3. H1473 cells were divided into the following three groups: control, METTL5, and METTL5 + si-IGF2BP3. Transfection with METTL5 overexpression construct increased cell viability. However, transfection with si-IGF2BP3 partially suppressed the METTL5 overexpression-induced enhanced cell viability (Fig. 4B). These experiments were also performed with H1299 cells with the same grouping. The findings revealed that IGF2BP3 overexpression suppressed the METTL5 knockdown-induced decreased cell viability (Fig. 4C). Similarly, IGF2BP3 knockdown mitigated the METTL5 overexpression-induced enhanced H1299 cell viability (Fig. 4D). This suggested that METTL5 enhances NSCLC cell viability through IGF2BP3.
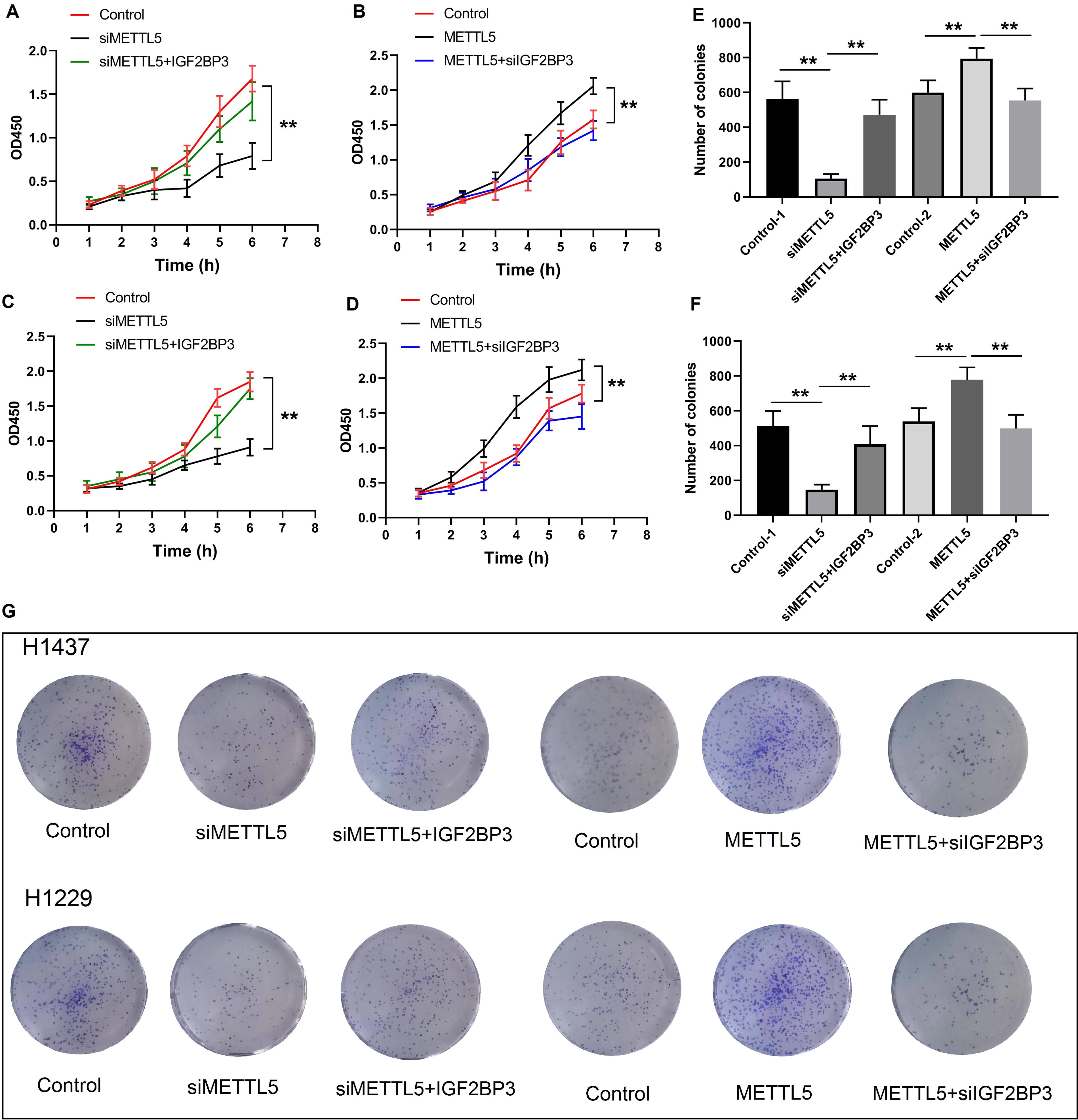
METTL5 regulates the proliferation of NSCLC cells through IGF2BP3.
The effects of METTL5 and IGF2BP3 on clonality were also examined. Cell clonality decreased upon METTL5 knockdown and increased upon METTL5 overexpression. Cotransfection with the IGF2BP3 overexpression vector and si-METTL5 restored the clonogenicity to the original level. IGF2BP3 knockdown suppressed the beneficial effects of METTL5 overexpression on clonogenicity (Fig. 4B-D). These findings indicate that METTL5 regulates NSCLC cell proliferation through IGF2BP3.
Interaction of METTL5 with IGF2BP3
The interaction between METTL5 and IGF2BP3 was verified using the Co-IP assay with METTL5 as the input group and IGF2BP3 as the immunoprecipitation (IP) group. Western blotting analysis revealed the presence of METTL5 in both input and IP groups. These findings indicate the presence of both METTL5 and IGF2BP3. IgG could not precipitate METTL5 and IGF2BP3, indicating that METTL5 and IGF2BP3 could not bind to IgG. The second band in the IP group indicated the precipitation of METTL5. This suggested an interaction between METTL5 and IGF2BP3 (Fig. 5A). IGF2BP3 was also used as the IP group. Consistent with previous results, the presence of METTL5 and IGF2BP3 and the inability of IgG to precipitate them were demonstrated. The addition of IGF2BP3 as the IP sample precipitated METTL5 (Fig. 5A). To perform bidirectional authentication, both Flag-METTL5 and HA-IGF2BP3 were incorporated, which was confirmed in the input sample. In Flag-METTL5, both METTL5 and IGF2BP3 were detected using immunoblotting. Similarly, in HA-IGF2BP3, both METTL5 and IGF2BP3 were detected using immunoblotting (Fig. 5B). This confirmed the interaction between METTL5 and IGF2BP3.

Interaction of METTL5 with IGF2BP3.
Effect of IGF2BP3 on NSCLC growth in vivo
To further demonstrate that METTL5 promotes NSCLC growth through IGF2BP3 in vivo, IGF2BP3 overexpression construct-transfected and si-IGF2BP3-transfected cells were used to establish the tumor-bearing nude mouse models (six mice per group). During the 20 days of modeling, the body weight of the experimental animals increased from 18 to 23 g. Additionally, the body weight was significantly different between the groups (Fig. 6A). The mortality rate in mice injected with si-IGF2BP3-transfected cells was lower than that in mice injected with IGF2BP3 overexpression-transfected cells (p < 0.05) (Fig. 6B). IGF2BP3 knockdown suppressed tumor growth. In contrast, IGF2BP3 overexpression enhanced tumor volume and quality (Fig. 6C, D). After euthanasia, the tumor was collected and displayed (Fig. 6E). These findings further suggest that IGF2BP3 promotes the growth of NSCLC.
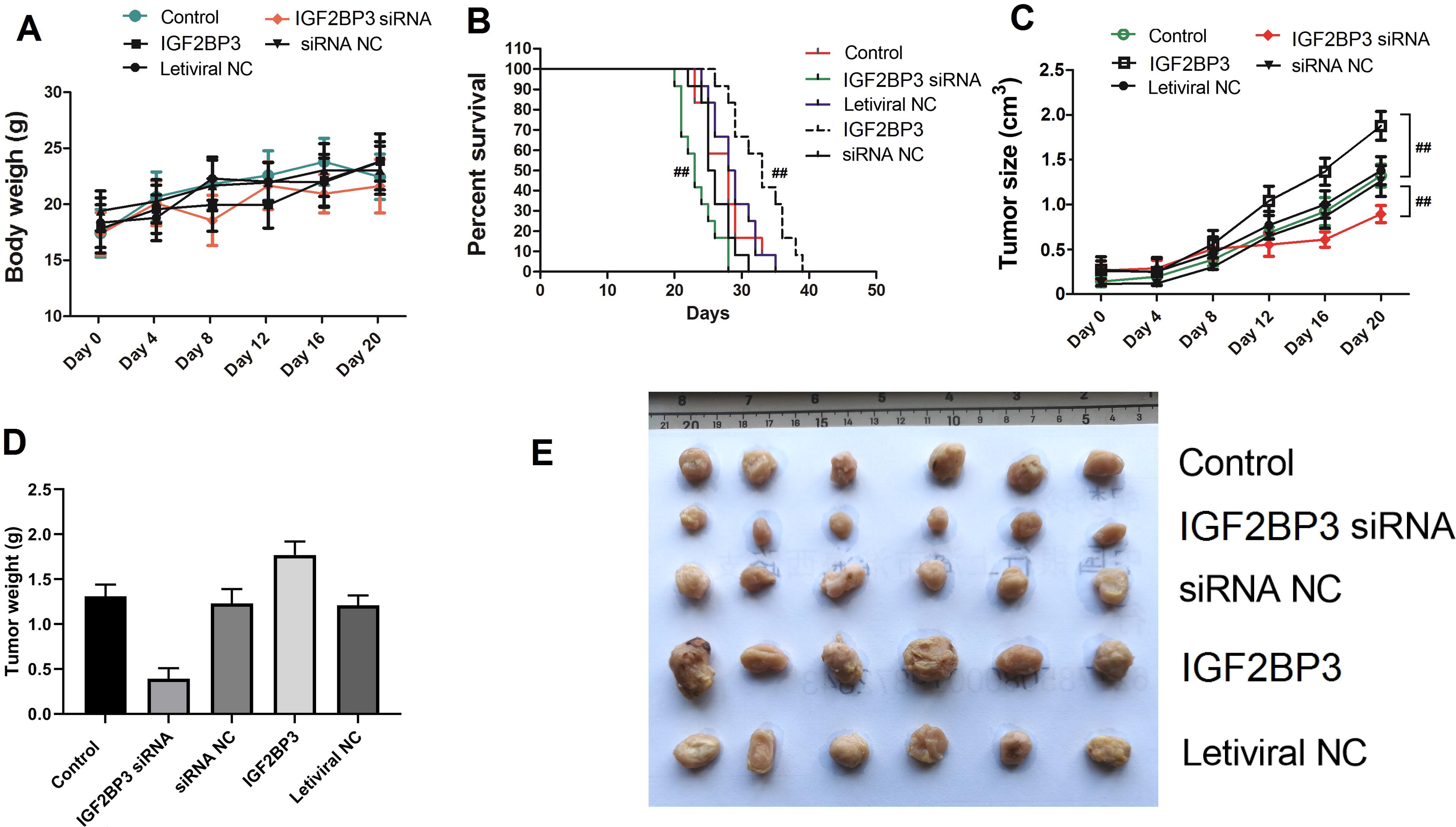
IGF2BP3 promotes NSCLC growth in vivo.
Discussion
In China, the incidence of NSCLC is approximately 54.3/10,000 and the average age of NSLC incidence is decreasing (Luo et al, 2021). The 5-year survival rate of patients with NSCLC, which is related to the timing of diagnosis, treatment methods, and physical condition of patients (Nie et al., 2021), is approximately 20% (Zheng W et al., 2020). No effective treatments have been developed for NSCLC as the mechanism underlying NSCLC occurrence and progression has not been elucidated.
METTL5, which contains a methyltransferase-like domain, regulates gene expression by modulating m6A modification and contributing to RNA stability (Han et al., 2022). Additionally, METTL5 is involved in various biological processes, such as cell cycle and protein translation regulation (Sepich-Poore et al., 2022). However, the specific function of METTL5 in human cells is controversial. Recent studies suggest that METTL5 can directly contribute to the pathogenesis of some cancers. For example, the expression of METTL5 is significantly upregulated in various malignant tumors, such as liver cancer and breast cancer, and is closely related to the prognosis and survival rate of patients (Xu W et al., 2023) (Peng et al., 2022). Additionally, the specific inhibition of METTL5 can effectively inhibit the proliferation and metastasis of cancer cells (Chen et al. 2023).
In this study, the role of METTL5 in NSCLC was examined using bioinformatics analysis. METTL5 upregulation was associated with poor prognosis of patients with NSCLC. Analysis of clinical samples confirmed the bioinformatics analysis results. The role of METTL5 in NSCLC cell proliferation was determined by silencing and overexpressing METTL5. Additionally, the role of METTL5 in the proliferation of NSCLC cells was assessed using the viability and clone formation assays. METTL5 regulates m6A modification, which is reported to be involved in the proliferation of NSCLC cells (Zhang Z et al., 2021). Furthermore, METTL5 is involved in the immune infiltration of NSCLC (Sun et al., 2020). Although studies on the role and mechanism of METTL5 in cancer are ongoing, METTL5 is suggested to be an important factor in the proliferation of NSCLC cells.
IGF2BP3, an RNA-binding protein, regulates RNA and is involved in functional regulation as a protein component (Mizutani et al., 2016). Recent studies have suggested that the functions of IGF2BP3 are similar to those of METTL5, including m6A modification (Zhang N et al., 2022). IGF2BP3 is upregulated in various cancers and is related to proliferation. IGF2BP3 promotes the expression of RCC2 (Zhang N et al., 2022), HDGF (Wang Q et al., 2020), and circARID1A (Ma et al., 2022) by regulating the translation and stability of mRNA and consequently promotes cell proliferation. Furthermore, IGF2BP3 modulates signaling pathways to regulate cancer stem cells (Conway et al., 2016) and drug resistance (Liu Y et al., 2017). IGF2BP3 overexpression enhances the resistance of tumors to chemotherapy drugs and targeted drugs, leading to increased risks of treatment failure and recurrence in patients (Yang et al., 2021) (Hsu et al., 2015). In NSCLC, IGF2BP3 can promote CD8+ T cell immunosuppression (Liu Z et al., 2021), drug resistance (Zhang Z et al., 2021), and ferroptosis (Xu X et al., 2022) and can predict prognosis (Guo et al., 2021). However, the regulatory mechanism of IGF2BP3 in NSCLC cell proliferation is unclear.
Bioinformatics and clinical sample analyses revealed that IGF2BP3 expression was upregulated in NSCLC and was positively correlated with METTL5 expression. Furthermore, IGF2BP3 did not affect the expression of METTL5, whereas METTL5 affected the expression of IGF2BP3. Thus, the findings of this study and previous studies indicate that METTL5 regulates the expression of IGF2BP3 by methylating specific sites in IGF2BP3 mRNA. Functional experiments revealed that the overexpression of IGF2BP3 reversed the METTL5 knockdown-induced impaired cell proliferation. Additionally, IGF2BP3 knockdown mitigated the beneficial effect of METTL5 on NSCLC cell proliferation. This suggested that IGF2BP3 modulates the regulatory effects of METTL5 on the proliferation of NSCLC. Additionally, the Co-IP assay results revealed the interaction between METTL5 and IGF2BP3 in NSCLC cells, indicating the critical roles of the two proteins. Next, the beneficial effect of METTL5 on NSCLC cell proliferation involving IGF2BP3 was confirmed in vivo. IGF2BP3 interacts with METTL3, a METTL5 homolog, to promote immunosuppression in breast cancer (Wan et al., 2022). These findings indicate that METTL5 promotes the proliferation of NSCLC cells by interacting with IGF2BP3. Additionally, METTL5 and IGF2BP3 may form a protein complex to regulate the proliferation of NSCLC cells. However, further clinical studies must be performed with a higher number of samples. Additionally, the molecular mechanism and function of METTL5 and IGF2BP3 must be further elucidated.
In conclusion, METTL5 promotes NSCLC cell proliferation by interacting with IGF2BP3. These results indicate that METTL5 can serve as a potential biomarker and a molecular therapeutic target for NSCLC. The elucidation of the mechanism of METTL5 and IGF2BP3 will enable the development of effective cancer treatments and drugs.
Footnotes
Authors’ Contributions
S.G. conceptualized the study. S.G. and H.L. performed the experiments and prepared the article. H.G. and W.S. analyzed the data and prepared the figures. All authors have read and agreed to the published version of the article.
Data Availability Statement
The authors declare that the data supporting the findings of this study are included in the article or were obtained from publicly available databases and online tools. The raw data can also be requested from the corresponding author.
Author Disclosure Statement
All authors declare no competing interest.
Funding Information
No funding was received for this article.