
Abstract
Background:
COVID-19 is one of the worst pandemics worldwide, and its diagnosis and treatment are of great importance. Recent evidence has shown that circular RNA (circRNA deregulation is involved in different infectious diseases. In the present study, we tried to investigate the expression of cirRNAs RasGEF1B (hsa_circ_0127052), HIPK3 (hsa_circ_100783), and GATAD2A (hsa_circ_0050236) in COVID-19 patients.
Methods:
Using quantitative real-time polymerase chain reaction, the expression profiles of candidate circRNAs were detected in 57 COVID-19 patients and 51 healthy controls. As part of the process of identifying a candidate circRNA that is sensitive and specific, receiver operating characteristic (ROC) curves were also utilized.
Results:
Our results showed higher expression levels of circRasGEF1B and circHIPK3 in COVID-19 patients, however, circGATAD2A showed no statistical difference between patients and controls. ROC curves showed that circRasGEF1B (hsa_circ_0127052), and HIPK3 (hsa_circ_100783) had favorable specificity and sensitivity, whereas GATAD2A (hsa_circ_0050236) did not.
Conclusion:
In summary, our study highlights the potential of CircRasGEF1B (hsa_circ_0127052) and HIPK3 (hsa_circ_100783) as biomarkers for COVID-19 diagnosis due to their high expression levels and demonstrated diagnostic accuracy. These findings suggest that circRNAs could play a crucial role in the development of diagnostic tools for COVID-19, providing a new avenue for early detection and management of the disease.
Introduction
Over the past two decades, coronaviruses have been associated with significant outbreaks in East Asia and the Middle East, including severe acute respiratory syndrome (SARS) in 2002 and Middle East Respiratory Syndrome (MERS) in 2012. Since late 2019, the novel coronavirus, severe acute respiratory syndrome (SARS-CoV-2), caused the global pandemic known as coronavirus disease 2019 (COVID-19), posing a major health threat worldwide (Holmes and Enjuanes, 2003). COVID-19 shows a wide range of symptoms in different people and is therefore classified into mild, moderate, and severe. Fever, a dry cough, and fatigue are the most common signs and symptoms of the disease. There were often reticular patterns, consolidation lesions, and ground-glass opacity during chest computed tomography (CT) (Chen et al., 2020; Shi et al., 2020). The virus infects host cells by using its spike proteins to bind to angiotensin-converting enzyme 2 (ACE-2) receptors, leading to cell damage, immune response activation, and, in severe cases, acute respiratory distress syndrome (ARDS), multiple organ failure, or death (Chugh et al., 2021).
Recent research has shown that interactions between noncoding RNAs (ncRNAs), including circular RNAs (circRNAs) and microRNAs (miRNAs), are connected to the SARS-CoV-2 infection through the development of the circRNA/miRNA/messenger RNA (mRNA) axis (Demirci and Saçar Demirci, 2021; Firoozi et al., 2022). CircRNAs are a type of endogenous ncRNAs with a closed-loop structure that makes them resistant to degradation by RNA exonucleases, providing them with greater stability than linear RNAs (Wu et al., 2019). This unique feature suggests that circRNAs could be valuable in clinical diagnostics and therapeutics.
Aberrant expression of circRNAs has been reported in various diseases, including autoimmune (Zhou et al., 2019), cardiovascular (Altesha et al., 2019), and neurological disorders (Lu et al., 2019), as well as nervous system illnesses (Chen et al., 2019). However, there is limited research on the role of circRNA in viral infections (Mohammadisoleimani et al., 2022; Xie et al., 2021).
CircRNAs are also detectable in bodily fluids such as blood, saliva, which indicates their potential as biomarkers for illness prediction (Salzman, 2016). However, the majority of circRNAs’ functions remain unclear, especially concerning their expression in COVID-19 patients. MiRNAs, which are key posttranscriptional regulators of gene expression in the competitive endogenous RNA network, can have their activity affected by circRNAs acting as miRNA sponges (Bezzi et al., 2017; Hansen et al., 2013). Although circRNA anomalies have been implicated in several diseases (Han et al., 2017; Li et al., 2022; Mo et al., 2021), not much research has yet looked at the expression of circRNAs in the blood of COVID-19 patients. Therefore, we aimed to investigate the association of certain less-known circRNAs with COVID-19.
For example, CircGATAD2A (hsa_circ_0050236) is located on chromosome 1 and is derived from the GATAD2A gene. Previous studies have shown that by preventing autophagy, circGATAD2A aided in the reproduction of the H1N1 virus (Yu et al., 2019). Given that influenza A virus (IAV) and COVID-19 share common symptoms, such as fever, cough, pneumonia, and ARDS, understanding the role of circGATAD2A in COVID-19 is crucial. Similarly, cirRNA 100783, also known as circHIPK3 (hsa_circ_000888), has been explored in the context of immunosenescence and may serve as a distinctive biomarker for the aging of CD28-associated CD8+ T cells (Bai et al., 2021; Wang et al., 2015). Thus, we sought to examine its role in COVID-19 patients. Furthermore, circRasGEF1B (hsa_circ_0127052), located on chromosome 5 and derived from the GEF1B gene, has been shown to regulate intercellular adhesion molecule-1 (ICAM-1) via the toll-like receptor 4/lipopolysaccharide (TLR4/LPS) pathway (Ng et al., 2016). We hypothesized that this circRNA might also influence COVID-19 pathogenesis. For these reasons, we decided to compare the expression pattern of these three circRNAs, complete blood count (CBC) parameters, and clinicopathological characteristics in COVID-19 patients and healthy controls.
Materials and Methods
Patients
Ethics Committee approval for this study was obtained from Fasa University of Medical Sciences (ethical code: IR.FUMS.REC.1399.066), and consent forms were signed by each patient. The study protocol complies with the Declaration of Helsinki. This study consists of 57 COVID-19 patients and 51 healthy controls. Quantitative real-time polymerase chain reaction (qRT-PCR) test was done to confirm the COVID-19 infection. Nasopharyngeal swabs were used for the COVID-19 sampling. Samples were collected during the peak of the Delta variant outbreak, and some were sent to the Pasteur Institute in Tehran, where the Delta variant was confirmed. Thus, it is likely that the majority of the patients were infected with this variant. The inclusion criteria for the control group required no history of COVID-19 infection, a negative qRT-PCR test for SARS-CoV-2 and influenza types A and B, no symptoms suggestive of COVID-19 at the time of recruitment, and no close contact with confirmed COVID-19 cases within the past 14 days. The recruitment of participants for this study was conducted at Vali-e-Asr and Shariati hospitals, located in Fasa, Fars, Iran. The blood samples were dispensed in ethylenediaminetetraacetic acid tubes within 3 h of collection and transported to the laboratory for RNA extraction.
Extraction of RNA and cDNA synthesis
Each participant had 6 mL of peripheral venous blood drawn, and peripheral blood mononuclear cells (PBMCs) were extracted within 2 h using Ficoll-Paque PLUS (GE Healthcare) density gradient centrifugation and kept at −80°C. TRIzol reagent was used to extract total RNA from PBMCs. The integrity of the RNA was verified using agarose gel electrophoresis after it was quantified by a spectrophotometer (NanoDrop BioTek, HTX multimode reader). Total RNA was collected for reverse transcription to create cDNA, as directed by the PrimeScript RT Reagent Kit (BioFact™, Cat.No: BR441-096). All reactions were performed according to 45 cycles at 95°C for 15 s, and then at 60°C for 30 s.
Quantitative polymerase chain reaction in real time
The RealQ Plus 2× Master Mix Green with High ROX (Amplicon, Cat.No: A325402-25) was used for RT-qPCR (ABI, USA). The reaction volume included 7.5 μL of SYBR Green PCR Master Mix, 0.75 μL of forward and reverse primers, 1 μL of cDNA, and as much ddH2O as needed to reach a total volume of 15 μL. All reactions were executed following a thermal cycling protocol of 45 cycles, consisting of denaturation at 95°C for 15 s, followed by annealing and extension at 60°C for 30 s. The primers are all displayed in Table 1 and were all created using the Circ primer software (BIOINF, 2024) . A backsplicing analysis predicts the presence of hsa_circ_0127052 at exons 2-4 of the GEF1B gene, hsa_circ_0050236 at exons 3-5 of the GATAD2A gene, and hsa_circ_100783 (hsa_circ_000888) at exons 2 and 3 of the HIPK3 gene locus. As an internal control, the B2M gene was used. The fold change in circRNA expression levels was calculated using the comparative CT (2−ΔΔCT) method. Sysmex XN-9000 hematological analyzers were used to perform routine CBC tests.
Specific Circular RNA Primers Used for Real-Time Polymerase Chain Reaction Analysis
circRNAs, circular RNAs.
Statistical analysis
SPSS Statistics software v.26 was used to conduct statistical analysis (SPSS, IL, USA), and GraphPad Prism version 9 software was utilized for the purpose of graphical visualization. The fold changes of circRNAs were calculated and compared separately in each group. The Mann-Whitney test was used for comparing circRNA expression levels. To evaluate the diagnostic value of circRNAs RasGEF1B (hsa_circ_0127052), HIPK3 (hsa_circ_100783), and GATAD2A (hsa_circ_0050236), receiver operating characteristic (ROC) curves were constructed, and CBC parameters were analyzed in patients with COVID-19 and healthy controls. The chi-square test was used to compare differences in sex and blood group between the groups, while one-way analysis of variance (ANOVA) was used to compare differences in age. Statistical significance was defined as a p value of equal or less than 0.05 (p ≤ 0.05).
Results
Patients’ basic information and demographic data
A total of 57 symptomatic COVID-19 patients, including 22 (38.6%) females and 35 (61.4%) males with a mean age of 50 years, and 51 healthy controls comprising 22 (43.1%) females and 29 (56.9%) males with a mean age of 43 years participated in this study. Patients and the control group matched in terms of age and sex. Among symptomatic COVID-19 patients, 6 (10.5%) had cardiovascular disease, 1 (1.8%) had kidney disease, 1 (1.8%) had diabetes, 1 (1.8%) had asthma, and 1 (1.8%) had immune system defects (Table 2). The different symptoms of patients with symptomatic COVID-19 are summarized in Table 3.
Participant Baseline and Demographic Information (p Value 1: In Comparison with Negative Control)
Age, mean SD, p value.
Negative control 43, 17 COVID-19 50 20 0.143.
Several Symptoms and Characteristics of COVID-19 Patients
BMI, body mass index.
Comparison of circRNA expression levels between healthy controls and COVID-19 patients
Profiles of circRNA expression showed that the expression of circRasGEF1B (hsa_circ_0127052) and circHIPK3 (hsa_circ_100783) was significantly upregulated in COVID-19 patients in comparison with healthy controls (Table 4, Fig. 1A, B). However, circGATAD2A (hsa_circ_0050236) did not show any significant difference between the two groups (Table 4, Fig. 1C).
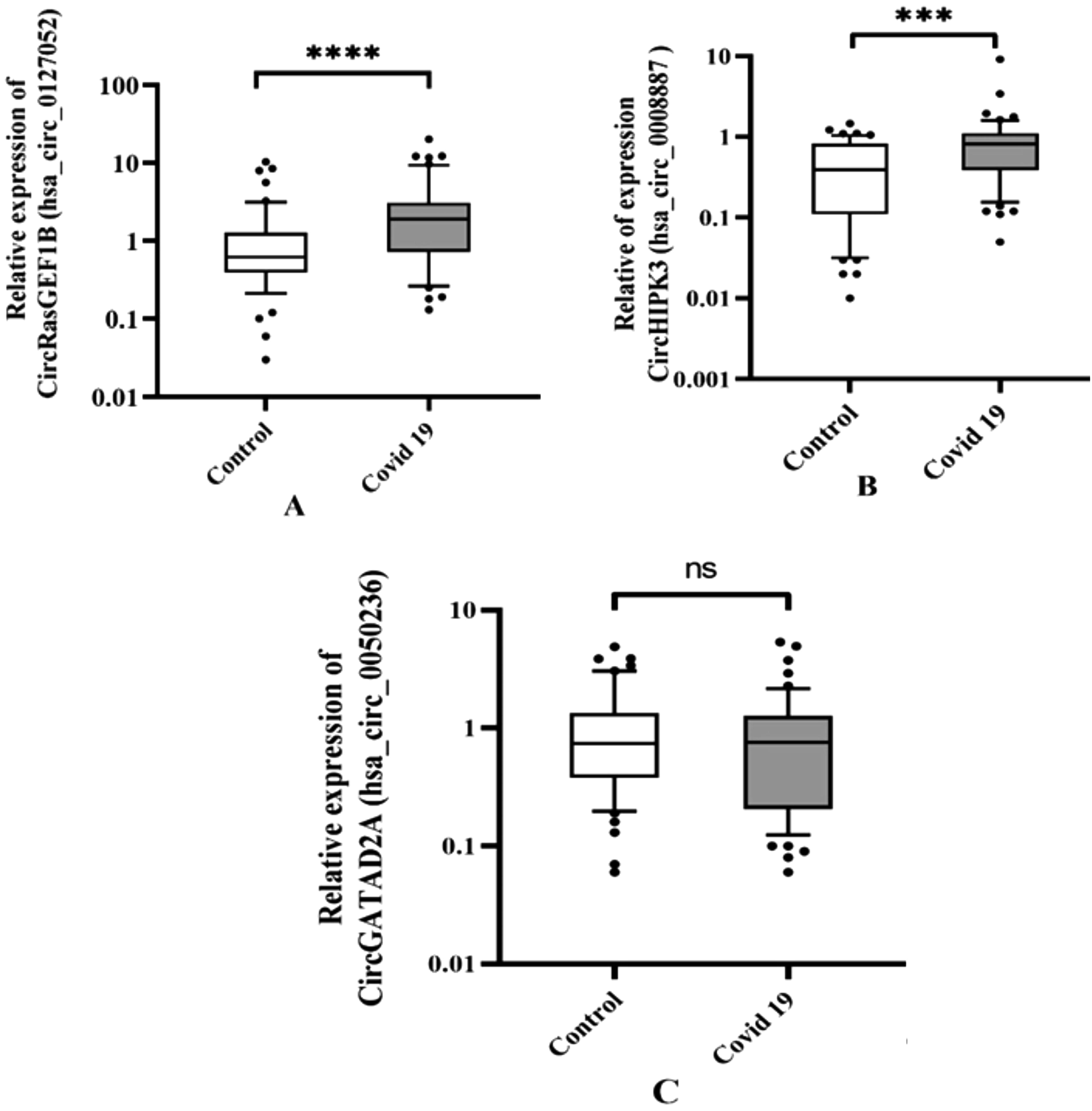
The relative levels of expression of hsa_circ_0127052 GEF1B and hsa_circ_0008887 HIPK3, hsa_circ_0050236 GATAD2 (ns: p > 0.05; *p ≤ 0.05; **p ≤ 0.01; ***p <0.001; ****p <0.0001).
A Comparison of Hsa_Circ_0127052 GEF1B, Hsa_Circ_0050236 GATAD2A, Hsa_Circ_100783 HIPK3 Expression Levels Between Negative Controls and COVID-19 Patients
ROC analysis of circRasGEF1B (hsa_circ_0127052), HIPK3 (hsa_circ_100783), and GATAD2A (hsa_circ_0050236) in patients with COVID-19
The area under the curve (AUC) of circRasGEF1B (hsa_circ_0127052), HIPK3 (hsa_circ_100783), and GATAD2A (hsa_circ_0050236) was 0.696 (0.593-0.798, p < 0.001), 0.717 (0.335-0.556, p < 0.001), and 0.445 (0.335-0.556, p = 0.335), respectively; the sensitivity and specificity were 82.5% and 59%, 83% and 55%, and 70.2% and 37%, respectively (Fig. 2).
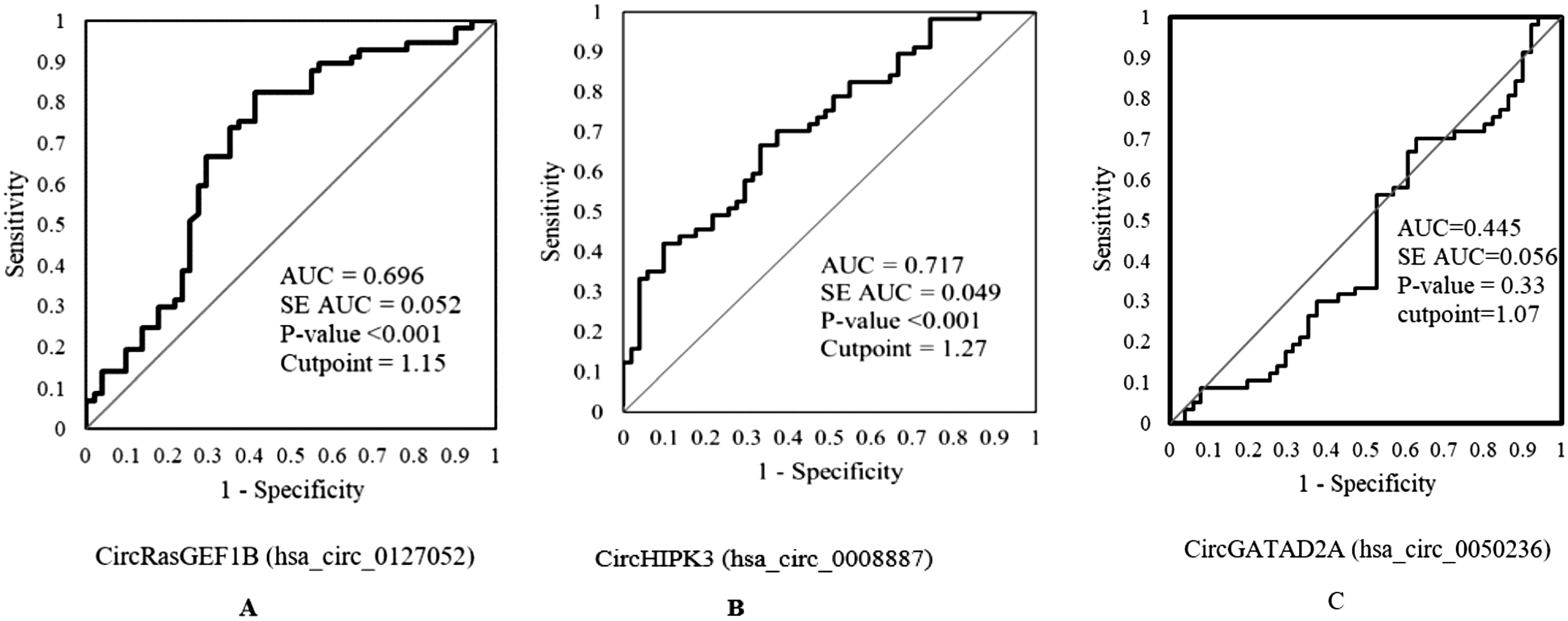
Potential diagnostic values of circRasGEF1B (hsa_circ_0127052), HIPK3 (hsa_circ_100783), and GATAD2A (hsa_circ_0050236) in COVID-19.
Results of the laboratory tests
A comparison is shown in Table 5 between COVID-19 patients and healthy controls in terms of CBC indices. According to our findings, the median values of hemoglobin, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, and red cell distribution width (RDW) are all significantly lower in COVID-19 patients than in healthy controls.
Hematological Parameters in COVID-19 Patients Compared with Controls
Hb, hemoglobin; RBCs, red blood cells; MCV, mean corpuscular volume; MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; HCT, hematocrit; WBC, white blood cells; LYMPH, lymphocytes; MONO, monocytes; GR, granulocytes; MPV, mean platelet volume; PLT, platelets; PCT, plateletcrit; PDW, platelet distribution width.
However, the red blood cell count, hematocrit, and mean corpuscular volume median values showed no significant difference between the two groups. Indicators of white blood cells and platelets in COVID-19 patients and healthy control are shown in Table 5. Compared with healthy control individuals, COVID-19 patients’ total white blood cell count, platelet count, lymphocyte count, and monocyte count median values and plateletcrit were significantly decreased. There was a significant increase in granulocytes in COVID-19 patients compared with controls. In contrast, mean platelet volume and platelet distribution width showed no significant differences between the two groups.
Discussion
COVID-19 is an infectious disease brought on by the SARS-CoV-2 virus. Based on the aforementioned functions, circRNAs are engaged in a number of pathophysiological processes. According to recent studies, circRNAs may be useful therapeutic targets in immunological disorders, including some conditions for which there is presently no cure. They may also serve as possible therapeutic targets and biomarkers for the diagnosis and prognosis of diseases. This study examined the link between COVID-19 and the expression of circRasGEF1B (hsa_circ_0127052), circGATAD2A (hsa_circ_0050236), circHIPK3 (hsa_circ_100783), and demographic and clinicopathological data.
The results of this study showed that patients with COVID-19 exhibit significantly higher levels of circRasGEF1B (hsa_circ_0127052) expression than healthy controls. Although our findings demonstrate potential roles for these circRNAs in COVID-19, it is important to note that the samples in our study were collected during the peak of the COVID-19 Delta variant outbreak. However, there was no association between circRasGEF1B expression and age, sex, or clinically relevant symptoms. In a study, researchers investigated the role of circRasGEF1B in the TLR4/LPS pathway, which plays a critical role in the innate immune response. They found that circRasGEF1B expression is induced by LPS in macrophages, indicating that it may play a role in the response to microbial infection (Ng et al., 2016). Knockdown of circRasGEF1B resulted in reduced expression of ICAM-1, a protein involved in immune cell adhesion and migration, in LPS-stimulated macrophages. Overexpression of circRasGEF1B increased ICAM-1 expression in both LPS-stimulated and unstimulated macrophages. The authors also found that circRasGEF1B can function as a sponge for miR-365, an miRNA that targets ICAM-1 mRNA. Overall, the study suggests that circRasGEF1B is a positive regulator of ICAM-1 expression in the TLR4/LPS pathway and may play a role in the immune response to bacterial infection (Ng et al., 2016). In another study, Chu et al. found that circRasGEF1B enhances antiviral immunity by regulating the miR-21-3p/MITA (mediator of interferon regulatory factor 3 activation) pathway. The miR-21-3p/MITA pathway is a signaling pathway that plays a role in regulating antiviral immunity. CircRasGEF1B acts as a sponge for miR-21-3p, which leads to reducing its levels and thereby increasing the expression of its target gene, MITA. This leads to the activation of downstream antiviral signaling pathways and enhances antiviral immunity (Chu et al., 2021). These findings suggest that circRasGEF1B may be a potential therapeutic target for enhancing antiviral immunity in viral infectious diseases.
Moreover, according to our findings, there was no significant difference between the expression of circGATAD2A in COVID-19 patients and healthy controls. Also, there is not any correlation between age or gender and the expression of circGATAD2A. A study investigated the effects of circGATAD2A on the A549 cell line IAV. Upon infection with the IAV, circGATAD2A expression soared, and when it was knocked down in A549 cells, autophagy was blocked and H1N1 proliferation was boosted. This study demonstrated that circGATAD2A controls LC3-II (microtubule-associated protein 1A/1B-light chain 3) production and upregulates p62 levels to control VPS34-dependent autophagy, which is crucial for destroying intracellular viruses. A rise in p62 levels raises the possibility that circGATAD2A is also in charge of suppressing autophagosome maturation (Yu et al., 2019). The relevance of the findings from other studies reporting alterations in circGATAD2A expression after infection lies in the potential significance of circGATAD2A in the defense against viral infection. Although circGATAD2A expression did not show any difference in our study, the previous findings suggest that circGATAD2A may play a role in the hosts’ defense mechanisms against viral pathogens. By exploring the probable roots of the difference between our results and those of other studies, we can gain insights into the complexity of circGATAD2A regulation during viral infections. Understanding the factors influencing circGATAD2A expression variations across different studies can help identify potential sources of discrepancies and determine future research directions. Moreover, highlighting the importance of circGATAD2A in the defense against viral infection emphasizes the need for further investigation to fully comprehend its functional significance. Future studies could delve into the specific mechanisms through which circGATAD2A may contribute to antiviral defense, exploring its potential roles in modulating immune responses, viral replication, or viral clearance.
This study examined circRNA100783 expression in patients diagnosed with COVID-19 and healthy individuals, and compared it with Wang et al. (2015), who found that circRNA100783 expression increased during aging and antigen exposure in CD8+ T cells. Our main finding was that circRNA100783 expression was significantly increased in COVID-19 patients compared with healthy controls, suggesting that this circRNA may be involved in the immune response to viral infection.
circRNAs play an important role in immune regulation and modulation, especially during aging and viral infections. In previous studies, circRNAs have been shown to regulate gene expression, interact with miRNAs, modulate signal transduction, and affect cellular functions such as proliferation, apoptosis, and differentiation. In addition, some circRNAs have been identified as potential biomarkers or therapeutic targets for autoimmune diseases, cancer, and viral infections (Carlos-Reyes et al., 2022; Tai and Chen, 2023; Wu et al., 2023).
Our findings also support Wang et al.’s hypothesis that circRNA100783 might function as a potential biomarker of immunosenescence. They discovered that the C1 and C4 groups of CD8+ T cells, which exhibit naive and senescent phenotypes, respectively, differed significantly in the expression of circRNA100783. To be more precise, the majority of the CD8+ T cells in the C1 group are naive, meaning they have never encountered an antigen. These cells stay dormant, waiting to react when coming into contact with their particular antigen. The C4 group, on the contrary, is made up of worn-out or senescent CD8+ T cells. These cells have lost their ability to function as a result of being repeatedly triggered over time, usually as a result of long-term exposure to antigens (Chen et al., 2024; Sheppard et al., 2014). In addition, they demonstrated that antigen exposure (C1) and time (C4) both had an impact on circRNA100783 expression during CD28-related CD8+ T cell aging. A circRNA-miRNA-gene network was also built, and it was proposed that circRNA100783 might control phosphoprotein-related signal transduction on CD28-dependent CD8+ T cell aging. CircRNA100783 may serve as an intracellular biomarker of immunosenescence, according to a long-term study.
The study has limitations, including a small sample size, cross-sectional nature, and lack of functional assays or mechanistic studies. Future research should explore circRNA100783′s role in immune aging and COVID-19.
In conclusion, our study revealed a significant increase in the expression of circRNA100783 in patients diagnosed with COVID-19 when compared with healthy individuals. This finding is consistent with previous studies that suggested that circRNAs are involved in immune regulation and modulation, especially in the context of aging and viral infections. Moreover, our finding supports the notion that circRNA100783 may serve as a novel biomarker of immunosenescence, as proposed by Wang et al. (2015). These collective insights highlight the significance of circRNAs as potential biomarkers and therapeutic targets in the context of immune dysregulation associated with aging and viral infections such as COVID-19.
According to ROC curve data, the assay’s sensitivity and specificity for circRasGEF1B (hsa_circ_0127052) and HIPK3 (hsa_circ_100783) indicated that this circRNA had a high potential as a diagnostic marker for COVID-19 patients (AUC = 0.696 and AUC = 717), respectively, so suggesting that they could be potential novel biomarkers. Nonetheless, GATAD2A (hsa_circ_0050236) was outside the acceptable range (AUC = 0.445).
Conclusion
In summary, our study findings indicate that COVID-19 patients exhibit elevated expression of hsa_circ_0127052 (GEF1B) and hsa_circ_100783 (HIPK3). However, the expression of hsa_circ_0050236 (GATAD2A) did not show any significant changes. These results suggest that hsa_circ_0127052 and hsa_circ_100783 may play a role in the pathogenesis or response to COVID-19, while hsa_circ_0050236 may not be directly involved in the disease process. Further investigations are warranted to elucidate the specific functions and mechanisms of these circRNAs in COVID-19 and their potential implications as diagnostic or therapeutic targets.
Author Confirmation Statement
E.M., A.S., B.M., J.K., M.F., Y.G., M.H.M., A.A., Z.F., and Y.M. are from Fasa University of Medical Sciences (Fasa, Iran), and A.D. is from Babol University of Medical Sciences (Babol, Iran), both where research and education are the primary functions.
Footnotes
Acknowledgments
The authors gratefully acknowledge the patients for their participation in this study. They express gratitude to Fasa University of Medical Sciences for providing funds and support in completing this study.
Authors’ Contributions
Y.M., E.M., A.S., A.D., and Z.F.: Study design, material preparation, data collection, and analysis. The first draft of the article was written by E.M. Laboratory experiments were performed by E.M., Z.F., and A.S. A.D., B.M., M.F., and M.H.M. were consultants for this project and critically revised the article and developed the idea. J.K., Y.G., and A.A. cooperated with the necessary permission to collect clinical data and samples and follow-up information and revised the article.
Data Availability Statement
All data will be available from the corresponding author, upon reasonable request.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This study was funded (ethical code: IR.FUMS.REC.1399.066) by Fasa University of Medical Sciences. Y.M. and Z.F. are the guarantors of this work and, as such, had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.