
Abstract
Abstract
Wagner, Peter D. Operation Everest II. High Alt. Med. Biol. 11:111–119, 2010.—In October 1985, 25 years ago, 8 subjects and 27 investigators met at the United States Army Research Institute for Environmental Medicine (USARIEM) altitude chambers in Natick, Massachusetts, to study human responses to a simulated 40-day ascent of Mt. Everest, termed Operation Everest II (OE II). Led by Charlie Houston, John Sutton, and Allen Cymerman, these investigators conducted a large number of investigations across several organ systems as the subjects were gradually decompressed over 40 days to the Everest summit equivalent. There the subjects reached a
Introduction

Left to right: Charlie Houston, Jack Reeves, John Sutton, Richard Riley, and Allen Cymerman. Photos all taken at USARIEM during OE II, October 1985.
The objective of OE II was to study a wide array of human biological responses along the way to, and then at, extreme simulated altitude as the subjects were gradually decompressed to a barometric pressure giving the equivalent inspired P

Decompression schedule showing simulated altitudes (calculated from the ICAO standard atmospheric pressure–altitude relationship) as a function of time.
OE II investigators studied cardiovascular, respiratory, metabolic, muscle, hematological, immunological, and neural responses (awake and asleep) in the 8 subjects. To date, these studies have resulted in at least 33 primary peer-reviewed research papers and 15 (16 if the present effort is included) reviews.
Unique elements of OE II
Given that there have been many research studies at high altitude carried out by many groups of investigators, both before and since, perhaps the primary question is what was unique about OE II. The uniqueness lies in two factors: (1) the remarkable breadth of interdisciplinary studies conducted both in parallel and in series by the large number of researchers on the same subject cohort and (2) the ability to make so many invasive and/or technically demanding measurements requiring catheter placement, tissue biopsy, complex electrical signal acquisition, and the like. The resulting accomplishments of OE II could never have been made in the field. For example, placement of both a systemic arterial and pulmonary arterial catheter was accomplished, allowing blood sampling and pressure measurements to be performed during maximal exercise, not just at sea level at the start of OE II, but also at intermediate stops: P
Chamber studies and field expeditions
It is probably useful to deal here with the inevitable argument about chamber versus field studies of altitude-related physiology and then to set that aside to allow focus on what OE II achieved. There is no denying that field studies examine the “real thing.” They encompass multidimensional insults to the body: hypoxia, exercise, cold, wind, stresses such as anxiety, fear, potential nutritional insufficiencies, potentially interrupted sleep, coping with injuries, illnesses, and so on. Rewards of course occur as well: the sense of accomplishment, the benefits of camaraderie, and the inherent beauty of the mountains. The insults additional to hypoxia may well affect the primary research, which focuses mostly on hypoxia, and produce results that might be different from those at similar hypoxic levels, where the above additional stresses might differ. They (weather, injury, illness) may even result in failure of the expedition. Additionally, not all subjects can make the summit of Everest breathing only ambient air, and time spent breathing oxygen may give rise to intermittent hypoxic exposure that may produce separate physiological effects not encountered in chamber studies, where ambient air is always breathed. Similarly, in field studies requiring ascents and descents before the summit is reached, altitude is not gained in a monotonic fashion, and this too may alter the kinetics of response compared with what would be seen in a steadily progressive ascent. That said, in OE II, some nights involved slight “descent.”
Chamber studies generally simulate only the hypoxia. To a lesser extent, they include highly regulated exercise, different from climbing, on an ergometer in a cramped environment, and different stresses: suffering the intrusions of a horde of investigators (who are bent on catheterizing, sampling, and measuring all they can with procedures that have potential risk), boredom, forced close contact with fellow subjects, and confinement within a small, sealed chamber. There is no climbing accomplishment, and the beauty of the mountains is certainly absent. On the other hand, there is a bed to sleep on, a toilet to sit on, showers, good food always at hand, and, in case of illness, the chamber door can be opened and the subject managed within seconds. The real benefit in chamber studies is that vastly more complex research can be performed than could be managed in the field and that control of extraneous factors such as cold or injury can be assured.
Field and chamber studies clearly have their pros and cons. This writer believes in their complementarity: both are valuable. To the extent that hypoxia dominates the findings, they can be similar, but the other differences need to be allowed for when results are examined. For OE II, a limitation as a chamber study was the speed of decompression. As Fig. 2 shows, subjects “summited” within 40 days, quite a bit faster than the ascent profile used in AMREE. One consequence of the rapid decompression profile in OE II was that, when passing through 20,000 ft, most subjects developed evidence of high altitude pulmonary edema (HAPE), and the “ascent” had to be interrupted with nighttime “descents” until subjects returned to normal. Time-dependent altitude responses may well have been affected by the rapid schedule, and, indeed, much has been written about the likely lesser degree of ventilatory acclimatization in OE II than in actual Everest ascent studies such as AMREE, described in this volume by Dr. West (West, 2010). That said, it will be pointed out later in this review that the similarities seem greater than the differences for many variables. Another limitation of OE II was that, because of the frequent need for taking blood by many of the investigators, the hypoxia-induced increase in Hb concentration was somewhat attenuated, average values increasing from 13.3 to only 16.9 g/dL (Groves et al., 1987). In AMREE, for example, Hb concentration averaged 18.4 g/dL at 8050 m (n = 2) (Winslow et al., 1984). To the extent that high hemoglobin at altitude is physiologically beneficial, which is not necessarily agreed to be the case (Calbet et al., 2002), OE II probably produced conservative results, especially in the context of maximal exercise and cardiac function.
Suffice it to say that OE II produced many historic firsts, and these and other important findings are now highlighted, with apologies to the original authors, in the remainder of this review. The order of presentation should not be assumed to reflect relative importance; further, it should be recognized that it is not possible within the allotted space to describe any of the results in detail or how the changes progressed with every step of increasing decompression. Indeed, not every finding can be discussed here. The focus will generally be on the most positive and novel findings, especially at the summit equivalent. Fifteen of the 30 primary research publications generated by OE II are cited in the text; the remaining 15 appear separately in the reference list along with 5 review articles.
Exercise capacity
As has been known for many years, exercise capacity, measured as
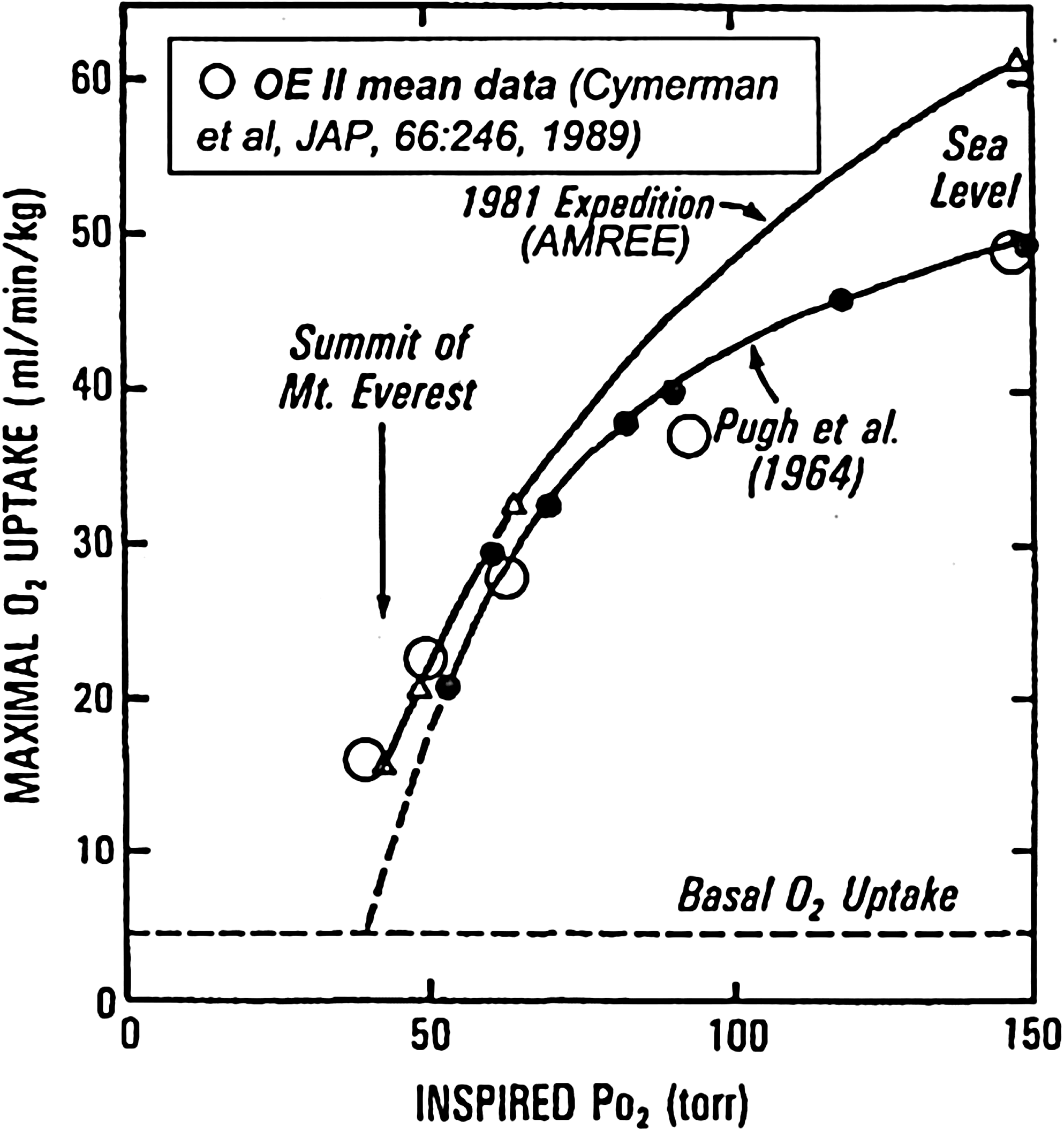
Maximal O2 uptake against inspired P
At the “summit”,
Left ventricular function
It is well known that cardiac output at maximal exercise is lower in chronic hypoxia than at sea level. This was found in OE II also. At maximal exercise at P
In OE II, we attempted to investigate, at least in part, the cause of this fall in maximal cardiac output. Potential explanations include (1) hypoxic ventricular dysfunction, (2) reduced venous return (potentially from hypovolemia) and thus lower ventricular filling pressures, (3) altered autonomic input to the heart, or (4) reduced demand for blood flow secondary to reduced O2 demand from other effects of hypoxia on O2 transport. Figure 4, using data from Reeves and colleagues (1987), shows that, whether related to left or right atrial pressures as estimates of their respective ventricular filling pressures, cardiac output is if anything higher, and not lower, at any given filling pressure as altitude is gained. The same is found when stroke volume is examined rather than cardiac output. This occurs in the face of increasing average systemic vascular resistance with increasing altitude (Reeves et al., 1987). In fact, using echocardiography, Suarez and colleagues (1987) found that ventricular function at all altitudes was similar to that at sea level, or even slightly enhanced.

Cardiac output from rest to exercise as a function of left and right atrial mean pressures (mean data). With increasing altitude, there is no suggestion that cardiac output is lower at any filling pressure. If anything, the opposite is seen (Reeves et al., 1987).
No interventions with fluid volume expansion or autonomic interruption were done in OE II to directly explore the possible roles of reduced filling pressures or altered autonomic control, respectively, but the data clearly showed that cardiac function was not at all impaired at any altitude. Thus, hypoxic impairment of cardiac function was not seen and cannot explain the reduction in maximal cardiac output. In this chamber study with good nutritional support, hypovolemia sufficient to reduce cardiac output by one-third seems unlikely, and other studies have shown that restoring the reduced maximal heart rate to sea-level values by vagal inhibition (Boushel et al., 2001) does not increase maximal cardiac output at altitude. This leaves the most likely explanation as hypothesis (4) above: reduced metabolic demand. What is clear is that the heart functions well at an arterial P
Right ventricular function and pulmonary vascular responses
Right ventricular function was also well preserved at altitude in spite of substantial pulmonary arterial hypertension (e.g., mean pressure 34 mmHg at P
Hypoxic pulmonary vasoconstriction is a well-established consequence of altitude, resulting in pulmonary arterial hypertension and, eventually, right ventricular hypertrophy. Pulmonary vascular resistance indeed rose progressively with altitude (Fig. 5), except during the acute hypoxic change from P

Mean values for pulmonary vascular resistance (PVR) at rest and exercise as a function of altitude (represented by air-breathing alveolar P
Pulmonary gas exchange
Pulmonary gas exchange can be examined in several ways. The major response to hypoxia is well known to be hyperventilation, and the degree of hyperventilation is reflected in alveolar P
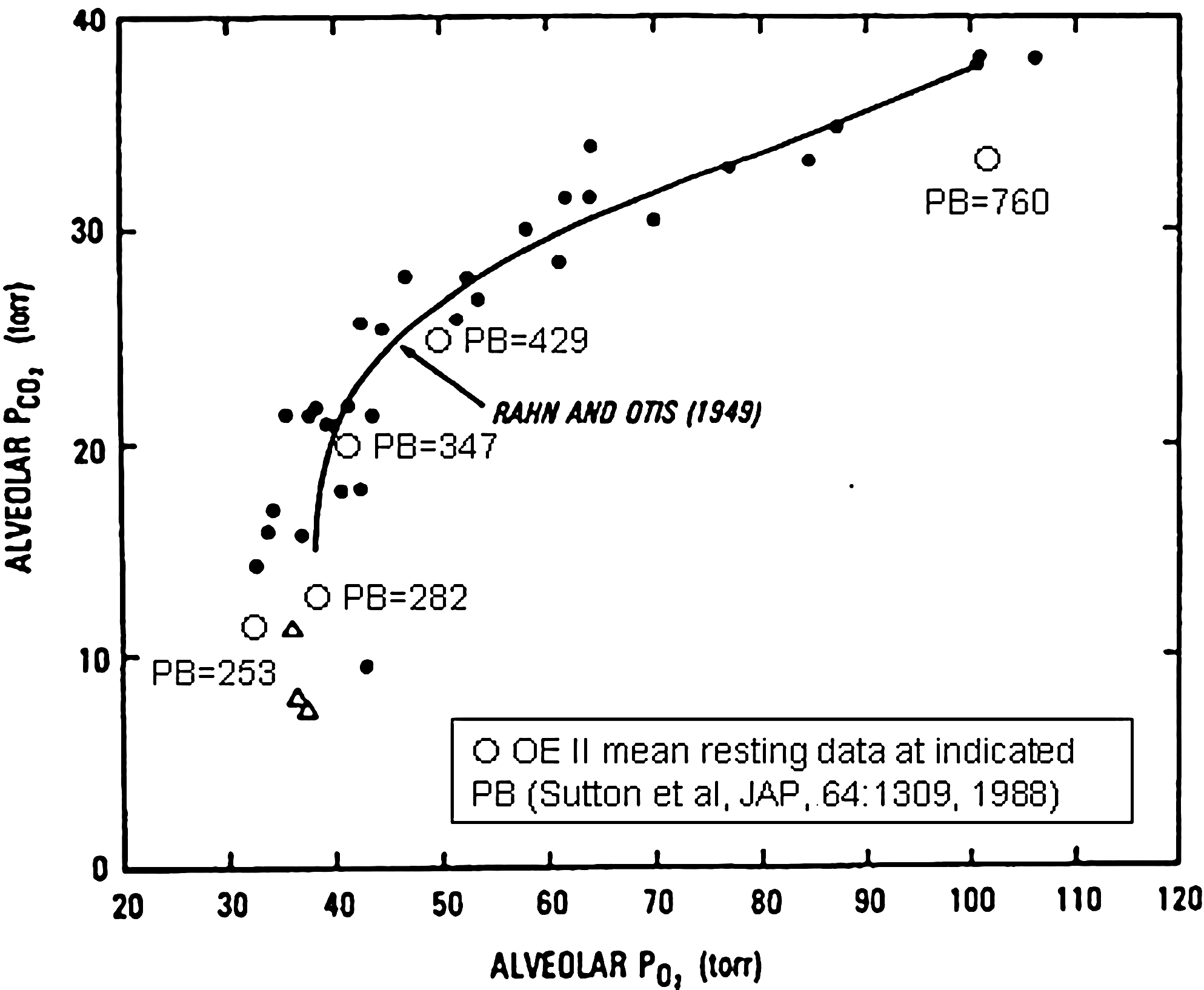
O2–CO2 diagram showing alveolar P
Gas exchange is also affected by ventilation–perfusion (
Finally, alveolar–capillary diffusion limitation has been predicted to have a major effect on O2 transfer and arterial saturation, increasing as altitude is gained (West and Wagner, 1980). This was indeed found to be the case in OE II (Wagner et al., 1987b), where on the summit at maximal exercise, diffusion limitation accounted for almost all the 6-mm Hg alveolar–arterial P
As a result of alterations in P
Acid–base balance
Although not a major focus of OE II, acid–base changes with altitude were examined in the course of exercise studies when arterial catheters were in place (sea level and altitudes corresponding to P
Peripheral gas exchange
Peripheral gas exchange was not a topic of research proposed for OE II. Nonetheless, it was remarkable to find how high mixed venous P
Skeletal muscle function
The great interest in O2 transport at altitude occurs partly because of the O2 needs of the skeletal muscles, which need O2 to allow climbing in the first place. Thus, studying skeletal muscle adaptation to extreme altitude became a focus of OE II. Green and colleagues (1989b) biopsied vastus lateralis at sea level pre-OE II, at P
OE II was a unique opportunity to gain further insight into the lactate paradox (the observations that, in spite of extreme hypoxia, blood lactate at a given exercise level rises less after spending time at altitude than immediately on ascent, and that blood lactate levels at exhaustion are progressively lower, and not higher, as altitude is gained). From muscle biopsies, it was found that muscle lactate levels at exhaustion were greatly reduced at altitude, compatible with lower power outputs (Green et al., 1989a). Thus, low blood lactate levels appear to reflect low muscle lactate levels.
Muscle structure was affected (MacDougall et al., 1991). The cross-sectional area of both type I and II fibers was significantly reduced, compatible with the well-known propensity for loss of muscle mass and body weight at extreme altitude. Muscle capillary density was increased, but only as a result of the loss of fiber area. The number of muscle capillaries was not altered. This is surprising insofar as muscle angiogenesis is believed to be driven at least in part by hypoxia-sensitive genes such as VEGF, possibly activated by HIF-1α (Forsythe et al., 1996).
Neuromuscular function was also studied in OE II (Garner et al., 1990) using a small- muscle mass group (ankle dorsiflexors). Effects of extreme altitude were mild to nonexistent, with force generation capacity and neuromuscular transmission preserved. There was evidence of slightly greater muscle fatigability.
In all, muscle changes other than loss of muscle mass were surprisingly mild or absent.
Neurological function
Although neuromuscular function was preserved (see above), neuropsychological testing revealed clear deficits resulting from exposure to extreme hypoxia in OE II. Hornbein and colleagues (1989) found persisting visual and verbal long-term memory deficits after subjects had returned to sea level. A provocative finding, also noted by Hornbein to be evident in the AMREE data, was that subjects with higher hypoxic ventilatory responsiveness displayed greater neuropsychological deficits. This was not expected, as higher ventilation should lead to greater arterial and thus cerebral oxygenation. However, Hornbein proposed that the greater degree of hypocapnia associated with higher ventilation may have caused sufficient cerebral vasoconstriction to more than offset the higher arterial P
Immunological function
A variety of in vivo and in vitro immune alterations were identified from 7 subjects during the ascent in OE II. Peripheral blood IgA and IgM immunoglobulin levels were increased at 25,000 ft, whereas in vitro peripheral blood mononuclear cell spontaneous and pokeweed mitogen-stimulated IgG/A/M production was unchanged. Spontaneous and stimulated in vitro production of interferon and natural killer (NK) cytotoxicity against the K562 cell line was unimpaired, and no increase in the generation of antinuclear antibodies was observed in the peripheral blood from any subject. Mucosal immunity was unimpaired, as determined by nasal pharyngeal washing measurements of IgA and lysozyme levels. In contrast, significant impairment of in vitro T cell function was documented by a 50% reduction in PHA-stimulated protein synthesis at 24 h of culture, followed by a 33% reduction in proliferation by 72 h by thymidine uptake. Simultaneous peripheral blood mononuclear cell subset analysis by flow cytometry indicated an increase in circulating monocytes without changes in B cells or T cell subsets. These findings suggest hypoxic exposure most likely alters normal immune regulation and function, T cell function more than B cell, or mucosal immunity through unidentified stress-mediated alterations in normal adaptive immunity, possibly mediated by monocyte-derived cytokines. Similar in vivo and in vitro immunologic alterations have been observed from subjects exposed to other environmental stressors, including space flight and military and mountaineering expeditions (Meehan et al., 1992; Meehan et al., 2001).
Metabolic and hormonal responses; nutrition and body composition
It has long been known that spending time at extreme altitude results in loss of body weight. This may come from reduction in fat mass, fat-free mass, or both, and causative factors may include hypoxia, poor diet, malabsorption, increased physical activity, dehydration, or stresses such as those mentioned in the introduction. In OE II, some of these factors could be excluded. Rose and colleagues (1988) showed that subjects lost 9% of their body weight on a palatable ad lib diet that included more than 3000 kcal/day of their favorite foods. In spite of food availability, intake fell progressively and substantially with altitude, from 3136 to 1789 kcal/day. However, even this reduction in intake, given the levels of physical activity (∼715 kcal), should have caused only a 2% weight loss, according to Rose. Dehydration can contribute to weight loss, but a rough fluid balance study (fluid intake vs. urine output) showed the subjects were maintaining hydration; but fluid losses from feces, hyperventilation, and excessive sweating were not measured. Computed tomography scans showed most of the weight was lost from fat-free (67%) sources, especially muscle. Malabsorption was not assessed, but these data suggest that hypoxia alone explains the majority of the findings.
Young and colleagues (1989) measured plasma lipid and certain hormonal profiles and found substantial changes over the course of OE II, although their mechanisms, significance, and longevity remain to be established. Fasting cholesterol levels fell by about 30% and triglyceride and insulin levels increased twofold, without changes in glucagon. Norepinephrine levels increased by a factor of 3 (with no change in epinephrine). We can only speculate that increased metabolism might have caused the weight loss.
Conclusions
In conclusion, OE II was a remarkable study, unique in its breadth and depth of topics studied and methods used. We found that at extreme altitude the heart does well, but the lungs do not (although the lungs can still support massive hyperventilation, there is substantial ventilation–perfusion inequality and, during exercise, diffusion limitation). Renal compensation for hypocapnia is far from complete after 40 days of progressive hypoxia. The muscles atrophy, accounting for most of the weight loss, but the hypoxia does not promote muscle angiogenesis or metabolic adaptations sufficient to overcome the loss of oxygen. The blood shows evidence of altered lipid metabolism and immune function. Cognitive function is impaired, but at the end of the day, normal subjects can still function and even exercise at 100 W (the equivalent of a moderately slow horizontal walk at sea level), in spite of an arterial P
Footnotes
Acknowledgments
This work was supported by the National Institutes of Health P01 HL091830. The author wishes to thank Dr. Madeleine Rose and Dr. Richard Meehan for help in describing the nutritional and immunological findings in OE II, respectively.
Disclosures
The author has no conflicts of interest or financial ties to disclose.