
Abstract
Abstract
Chen, Yu-Ching, Shin-Da Lee, Low-Tone Ho, and Cha-Hua Kuo. The effects of altitude training on the AMPK-related glucose transport pathway in the red skeletal muscle of both lean and obese Zucker rats. High Alt. Med. Biol. 12:371–378.—
Introduction
When skeletal muscles face physiological challenges (e.g., exercise and hypoxia), large amounts of AMP are produced. An increase in the AMP:ATP ratio generates a conformational change in AMPK. AMPK then becomes more easily phosphorylated by its major upstream kinase LKB1, a constitutively active enzyme that phosphorylates AMPK at position Thr172. LKB1 can be phosphorylated by a number of protein kinases, but the impact of such phosphorylation on LKB1 activity is still unclear (Woods et al., 2003). Recent studies have shown that phosphorylated AMPK can phosphorylate and activate AS160 (Akt substrate 160). AS160 is a newly identified potential trigger factor that is involved in the regulation of the translocation of intracellular GLUT4 from the intracellular pool to the plasma membrane (Kramer et al., 2006). GLUT4 is the major glucose transporter expressed in skeletal muscle (MacLean et al., 2002; Rodnick et al., 1992). A causal relationship between GLUT4 protein content and whole-body insulin sensitivity has been well established in transgenic models (Galuska et al., 1998). The responses sent through this chain of proteins (LKB1-AMPK-AS160-GLUT4) help muscles quickly absorb large amounts of useful glucose.
Previous studies have indicated that AMPK phosphorylation levels are abnormal in skeletal muscles. One study using obese Zucker rats has revealed abnormalities in AMPK activity in the epitrochlearis muscle before and after electronic stimulation (Barnes et al., 2002). People with body mass indexes (BMIs) greater than 30 also show blunted AMPK and AS160 activation in the basal vastus lateralis muscle after moderate exercise (Sriwijitkamol et al., 2007). In 2005, Sriwijitkamol et al. discovered that the basal levels of LKB1 and AMPK phosphorylation display decreasing trends in the gastrocnemius muscle of obese, insulin-resistant Zucker rats. Sriwijitkamol et al. (2006) have also shown that 5 weeks of running exercise does not reverse the abnormal basal pAMPK expression in obese rats.
According to our previous study, both exercise training and 8 hours of 14% daily hypoxia can improve insulin sensitivity and increase the skeletal muscle GLUT4 expression in normal rats (Chiu et al., 2004). Combined exercise and hypoxia (the altitude training) is known to increase the endurance of red muscles in athletes (Stray-Gundersen and Levine, 1999; Stray-Gundersen et al., 2001). Based on these previous findings, we investigated whether either hypoxia or altitude training can improve the LKB1-AMPK- AS160-GLUT4 pathway.
Therefore, in this study, we compared the effects of exercise, hypoxia, and a combination of exercise plus hypoxia (altitude) training on the expression levels of the proteins involved in the LKB1-AMPK-AS160-GLUT4 pathway in skeletal muscle from both lean (normal) and obese Zucker rats. Because the red gastrocnemius muscle, a slow-twitch muscle that is responsible for daily activities, is the most important muscle type for chronic endurance training (Ruegg, 1987; Sica and McComas, 1971). we chose this muscle for the analysis of protein expression. Both lean and obese Zucker rats were divided into control (n=7), exercise (n=7), hypoxia (n=7), and exercise plus hypoxia (altitude training, n=7) groups. After 6 weeks of training, the red gastrocnemius muscles were collected and analyzed to determine the effects of training on expression of the proteins involved in the LKB1-AMPK-AS160-GLUT4 pathway.
Methods
Animal models
Lean (Fa/fa or Fa/Fa, encodes for glutamine) and obese (fa/fa, encodes for proline) male Zucker rats (n=7 for each group) were housed in a temperature-controlled room (21±1°C) with a photoperiod of 12 hours of light and 12 hours of darkness. All of the rats were provided with unlimited water and chow (PMI Nutrition International, Brentwood, MO). This study was approved by the Animal Management Committee of Taipei Veterans General Hospital and conformed to the guidelines for the use of laboratory animals published by the ROC Department of Agriculture.
Experimental design and training protocols
Lean and obese Zucker rats (14 weeks of age) were randomly divided into four groups: control (n=7), exercise (n=7), hypoxia (n=7), and exercise plus hypoxia (altitude training, n=7). The exercise and hypoxia protocols were adapted from versions used in a previous study (Chiu et al., 2004) and described in the following paragraph. The treatments were administered every day for a period of 6 weeks.
The exercise protocol consisted of swimming in several specially-designed plastic containers (water depth, 70 cm; water temperature, 29±1°C). Two days before the test, all of the rats, including the controls, were put into the swimming containers for 10–15 min once a day to acclimate to the water. During the training process, the rats were swimming with a load equivalent to 2%–5 % of their body weight so that the rats could not float. Hypoxia was generated by placing the rats in a 4-room isobaric chamber (56 cm long, 43 cm wide, and 39 cm tall) infused with a constant 14% fraction of inspired oxygen and controlled by a hypoxic air machine (GAO2 Altitude, Australia). The oxygen concentration in the isobaric chamber was monitored throughout the experimental period with an oxygen sensor equipped with an alarm (GAO2 Altitude). Water was available throughout the duration of the hypoxic treatment.
The exercise training followed our own previously published protocol (Chiu et al., 2004). The swimming treatment was administered during the dark cycle (active phase). The swimming duration was initially 30 min, but was gradually extended to 90 min during the second week. In later weeks, the duration was further prolonged to 180 min by adding 20 min/day. The hypoxic treatment lasted for 8 h/day and was administered during the light cycle (sleep phase) with water remaining accessible. In the exercise plus hypoxia group (altitude training group), the rats were trained in a “living-high/training-low” (LHTL) manner. The progressive daily exercise training (during the dark cycle) was always administered for 4 h after 8 h of sleep in a 14% hypoxia chamber (during the light cycle). During the treatment, the rats in the control group were prevented from eating, but water remained available. To avoid the acute effects of the last treatment, rats in all of the groups were allowed to recover for at least 48 h with food and water accessible prior to tissue collection. At the end of the treatments, all of the rats were anesthetized with an intraperitoneal injection of pentobarbital sodium (65.0 mg/kg body wt). The red gastrocnemius muscles were then excised and frozen with liquid N 2 using forceps for Western blotting.
Fasting glucose and insulin
After the completion of 5 weeks of training, fasting glucose and insulin levels were assessed in all of the rats after 12 h of fasting. The measurements were taken during the light cycle (sleep phase) 16 h after the last episode of training. Blood samples (0.6 ml) were collected from the abdominal vena cava and used to measure whole-blood glucose content and serum insulin content. The whole-blood content was measured using a one-touch digital sugar analyzer (Roche, Precision Sof-Tact™). Serum insulin levels were detected using a commercially available kit (Mercodia, Rat Insulin ELISA 10-1250-01).
Western blot analysis
Approximately 50 mg of skeletal muscle was homogenized (1:20 w/v) in HES buffer with a 1% protease inhibitor cocktail (Sigma, P2714) and a 1% phosphatase inhibitor cocktail (Sigma, P2850) using a Polytron system (Brinkmann Instruments, Littau, Switzerland). The homogenates were then centrifuged at 100 g for 10 min. The supernatants were collected, and the protein concentrations were determined using the BioRad Protein Assay reagent (Richmond, CA), according to the manufacturer's instructions. The sample homogenates and standards were diluted 1:1 with Laemmli sample buffer (125 mM Tris, 20% glycerol, 2% SDS, and 0.008% bromophenol blue, pH 6.8) and separated on a 10% SDS-polyacrylamide gel by electrophoresis at 120 V for 90 min. The fractionated proteins from the sample homogenates were then transferred to a polyvinylidene fluoride (PVDF) membrane. The membranes were blocked for 60 min in TTBS (Tris-buffered saline with 0.16 % Tween 20) containing 7% nonfat milk and then incubated overnight at 4°C with the following primary antibodies: anti-LKB1 (Cell Signaling, Beverly, MA), anti-AMPKα (Cell Signaling), anti-phospho-AMPK (Thr172; Cell Signaling), anti-AS160 (Upstate Biotechnology, Lake Placid, NY), anti-pAS160 (Upstate Biotechnology), anti-GLUT4 (Chemicon, Temecula, CA), and anti-β-actin (Cell Signaling). The membranes were then incubated with the appropriate anti-rabbit secondary antibody, diluted in TTBS. The muscle proteins were visualized using the ECL Western Blot Detection Kit (Amersham, Arlington Heights, IL) on Kodak film according to the manufacturer's instructions.
Statistical analysis
Two-way analysis of variance (two-way ANOVA) was used to analyze all of the variables across the experimental groups. Fisher's protected least significant difference test, which holds the value of a type I error constant for each test, was then performed to distinguish significant differences between pairs of groups. P<0.05 was considered statistically significant for all tests. Data are expressed as the mean±SEM.
Results
Body weight
Figure 1 shows the changes in body weight during the treatment period. At the beginning of training, the obese Zucker rats were significantly heavier than the lean rats (Fig. 1B). In the lean Zucker rats, no differences in weight were observed among the control, exercise, hypoxia, and exercise plus hypoxia (altitude training) groups during the treatment period (Fig. 1A). In the obese Zucker rats, the weight gain of the rats in the exercise plus hypoxia group (altitude training group) was attenuated compared with the control group. The mean body weight of the exercise plus hypoxia (altitude training) group of obese rats was significantly lower compared with their respective control group, after 25 days of training (Fig. 1B). No differences in weight were observed among the control, exercise, and hypoxia groups (Fig. 1B).
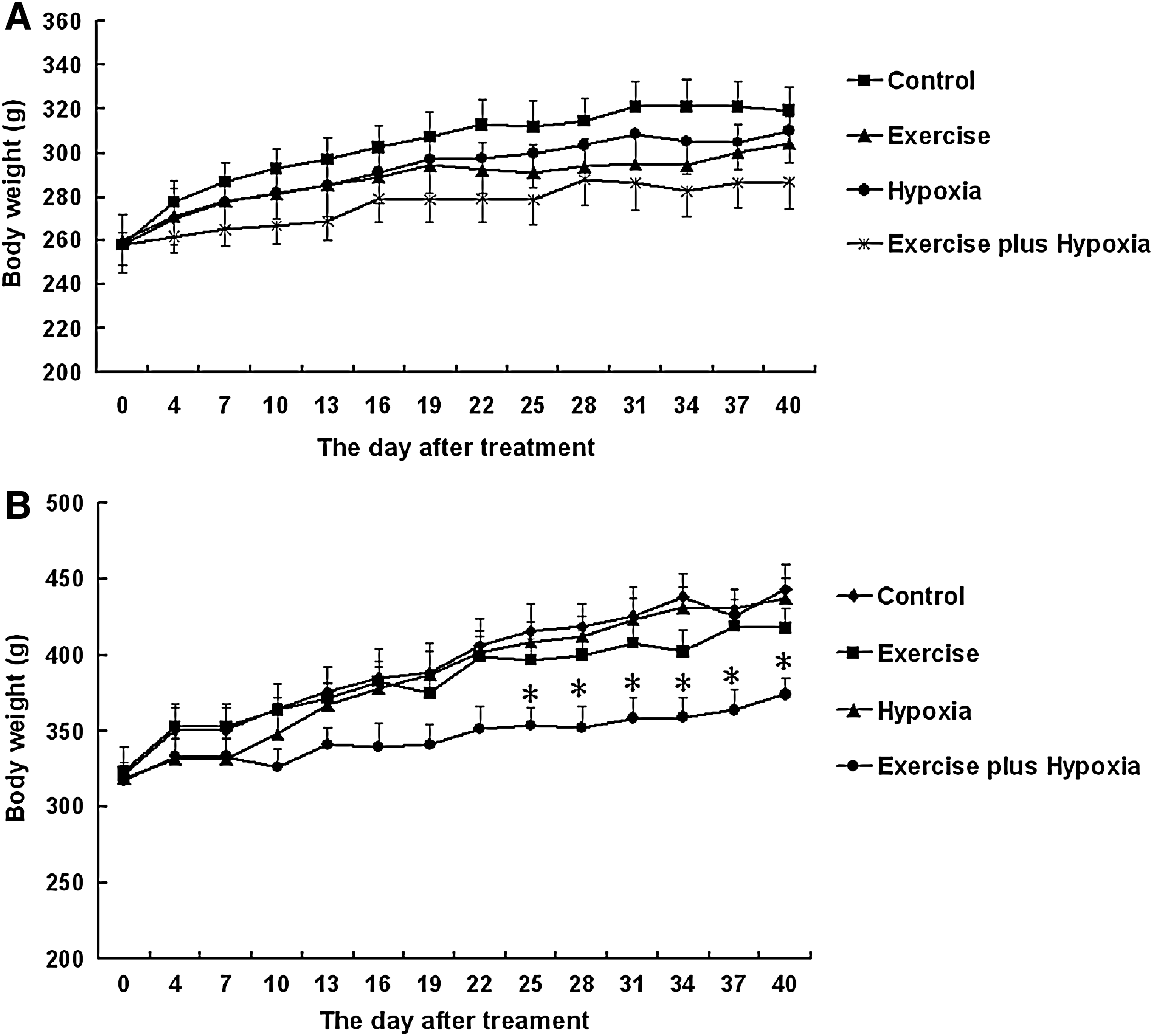
The body weights of lean
Fasting glucose levels
In the control groups, there were no differences in fasting glucose levels between the lean and obese Zucker rats (103.1±4.4 mg/dL vs. 109.3±7.2 mg/dL). In both the lean and obese groups, the exercise plus hypoxia groups (altitude training groups) had lower fasting glucose levels compared with their respective control groups (Fig. 2B). In the exercise and hypoxia groups of both lean and obese rats, there was an insignificant trend toward a decrease in blood glucose levels compared with their respective control groups.
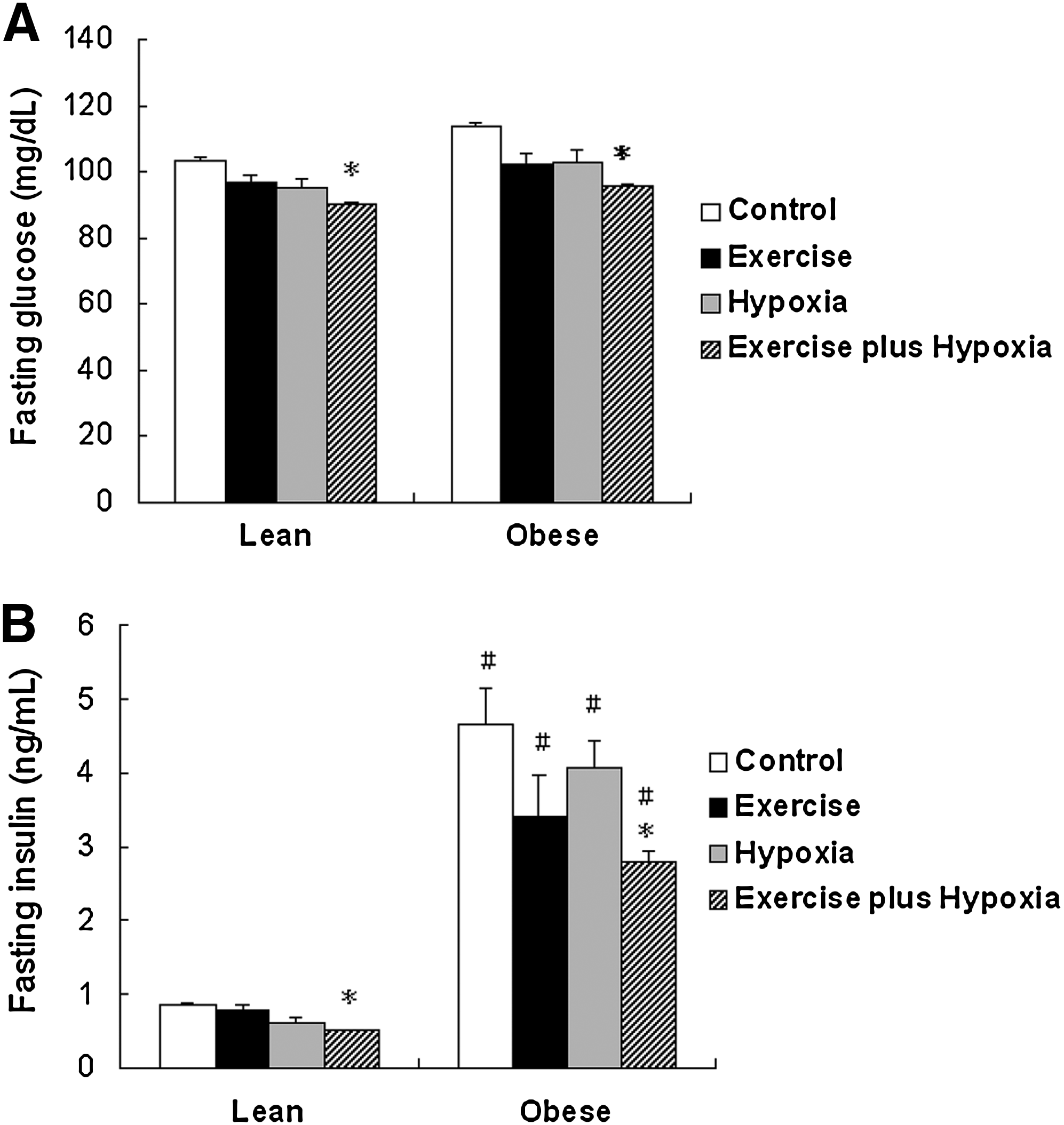
The fasting insulin and glucose levels of lean
Fasting insulin levels
In the control groups, the obese Zucker rats exhibited 5-fold higher fasting insulin levels compared to the lean rats (4.65±0.49 ng/mL vs. 0.87±0.02 ng/mL, p<0.05). In both the lean and obese Zucker rats, the exercise plus hypoxia group (altitude training group) had significantly lower insulin levels than their respective control groups (Fig. 2B). In the exercise and hypoxia groups of both lean and obese rats, there was an insignificant trend toward a decrease in fasting insulin levels compared with their respective control groups.
A comparison of basal LKB1, AMPK, AS160, and GLUT4 protein expression levels between lean and obese rats
In the control groups, there were no significant differences in the protein levels of the AMPK kinase-LKB1, AMPK, AS160, and GLUT4 in skeletal muscles of the lean and obese Zucker rats (Figs. 3, 4B, 5B, and 5D). The levels of AMPK phosphorylation, pAMPK-to-AMPK ratio, AS160 phosphorylation, and pAS160-to-AS160 ratio were significantly higher in the obese Zucker rats, compared to the lean Zucker rats (Figs. 4A, 4C, 5A, and 5C).
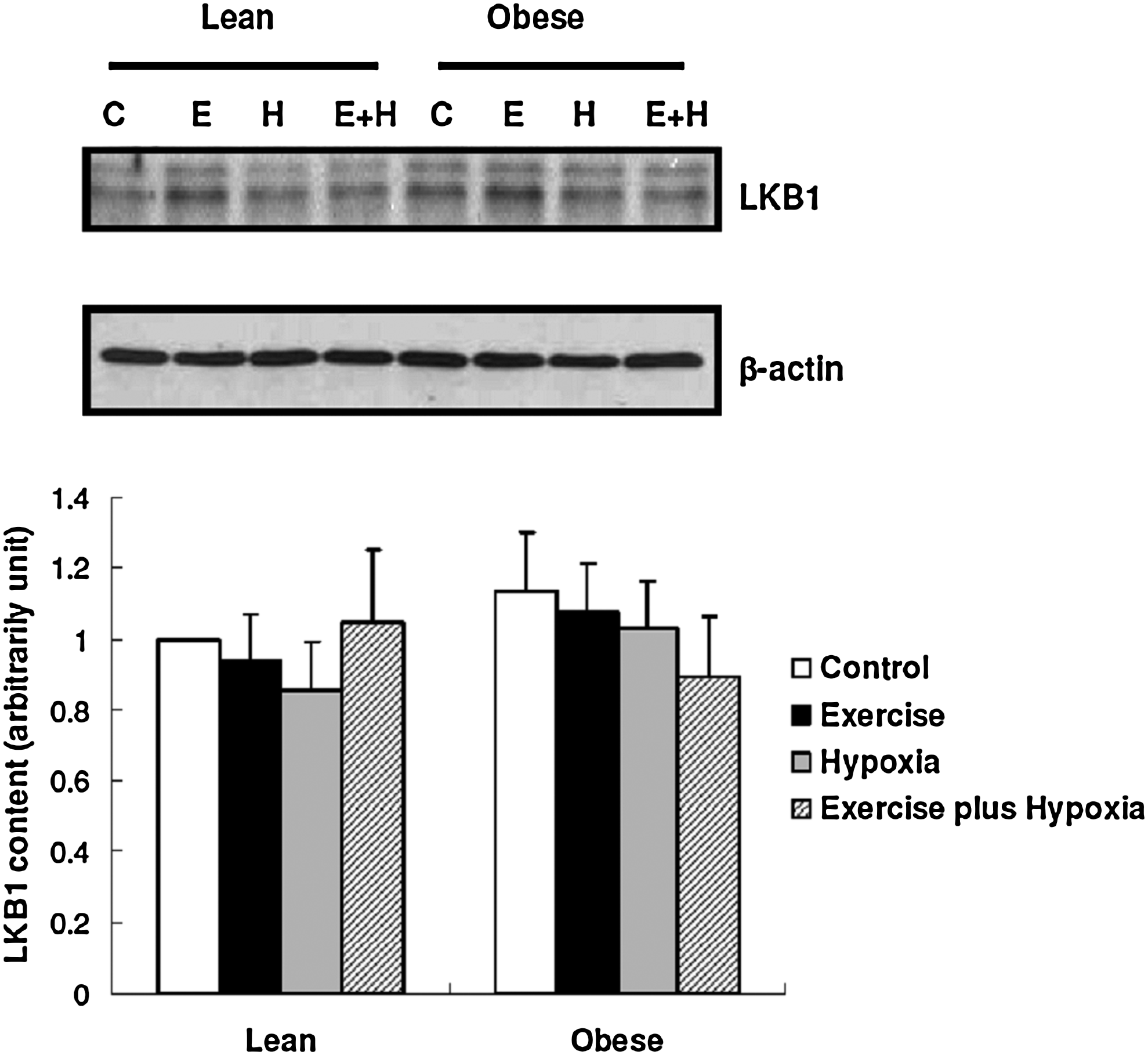
The levels of LKB1: Data are presented as mean±SEM.

AMPK phosphorylation

The levels of AS160
LKB1, AMPK, AS160, and GLUT4 protein expression in the lean and obese training groups
For both lean and obese Zucker rats, there were no significant differences in LKB1, AMPK, and AS160 expression between the groups. However, GLUT4 levels were increased in both of the exercise plus hypoxia groups (altitude training groups) compared to their respective control groups (Figs. 3, 4B, 5B, and 5D).
For lean Zucker rats, AMPK phosphorylation and the pAMPK-to-AMPK ratio were only significantly increased in the exercise plus hypoxia group (altitude training group) compared to its control group (Figs. 4A and 4C). AS160 phosphorylation and the pAS160-to-AS160 ratio were significantly increased in the exercise and the exercise plus hypoxia (altitude training) groups compared to their control groups (Figs. 5A and 5C).
For obese Zucker rats, AMPK phosphorylation and the pAMPK-to-AMPK ratio were significantly decreased in the exercise and hypoxia groups compared with those in the control groups (Figs. 4A and 4C). The relevance of comparing the AMPK phosphorylation and the pAMPK-to-AMPK ratio in the exercise and hypoxia groups did not mirror the changes in body weight and fasting glucose and insulin (Figs. 1B, 2A, 2B, 4A, and 4C). On the other hand, AMPK phosphorylation, the pAMPK-to-AMPK ratio, AS160 phosphorylation, and the pAS160-to-AS160 ratio were significantly decreased in the exercise plus hypoxia group (altitude training group) compared with those in the control group (Figs. 4A, 4C, 5A, and 5C). AS160 expression displayed an insignificant increase in the exercise plus hypoxia group compared with those in the control group (Fig. 5B).
Discussion
As previously shown, obese Zucker rats that were resistant to insulin exhibited abnormal levels of AMPK phosphorylation, and this abnormality was not corrected with exercise training (Sriwijitkamol et al., 2006). The present study further revealed that there was an increase in the phosphorylation of AS160, the newly discovered downstream regulator of AMPK that triggers GLUT4 translocation, in the muscles of obese Zucker rats. Most importantly, this study demonstrated that the altitude training model developed in our laboratory, which is similar to the popular “living-high training-low” (LHTL) training model used by endurance athletes, could successfully reverse the increased phosphorylation of AMPK and AS160 that has been observed in the muscles of obese Zucker rats to normal levels. This altitude training also suppressed the rate of weight gain, lowered fasting glucose and insulin levels, and increased GLUT4 protein levels in the muscles of the obese rats by approximately 50%. Moreover, in the lean (normal) Zucker rats, the altitude training lowered fasting glucose and insulin levels and increased the phosphorylation of AMPK and AS160, as well as GLUT4 protein levels, in the muscle. Our study is the first to suggest that altitude training may be a potentially effective nonpharmacological strategy for the prevention and treatment of metabolic syndrome based on its effects on the AMPK-related glucose transport pathway.
In this study, we also observed that either the 8-hour daily hypoxia (14% O2) treatment or progressive swimming-exercise training alone were able to reverse the increases in AMPK phosphorylation observed in the obese rats to the levels observed in the lean rats. These results are the first to demonstrate that long-term mild intermittent hypoxia has the same effect as progressive swimming-exercise training on the reversal of the increased phosphorylation of AMPK that has been observed in obese subjects. Mild intermittent hypoxia may be an appropriate treatment for obese subjects whose mobility issues make exercise difficult.
According to our findings, the protein levels of LKB1, a major upstream kinase of AMPK, in the red gastrocnemius muscles of the control group of obese Zucker rats were no different than those of the lean controls. This finding conflicts with a previous report by Sriwijitkamol et al. (2006), who observed lower LKB1 levels in the mixed gastrocnemius muscles of obese Zucker rats. Some potential reasons for this difference include differences in 1. animal age; 2. muscle type; 3. fasting time before muscle tissue collection, and the most likely, 4. muscle fiber composition. The gastrocnemius muscle contains a combination of fiber types. The superficial portion of the muscle is made up of white (glycolytic) fibers, which express substantially less LKB1 than the deeper portion of the muscle (red muscle) (Taylor et al., 2004). Whether obesity selectively influences LKB1 expression in different muscle fibers is currently unknown.
In the obese Zucker rats from the control group of our study, the basal levels of AMPK phosphorylation in the red gastrocnemius muscles were increased compared to the lean controls. This result is inconsistent with the findings of Sriwijitkamol et al. (2006), who observed that obese Zucker rats had lower levels of AMPK phosphorylation in the mixed gastrocnemius muscle compared to the lean rats. However, Sriwijitkamol et al. used fasted animals in their study, while the rats in our study had access to chow. It has been reported that a reduction in the availability of glucose can increase AMPK phosphorylation to facilitate energy utilization (de Lange et al., 2007). Thus, we posit that the low pAMPK levels observed by Sriwijitkamol et al. are due to the inability of AMPK in the muscles of obese rats to be phosphorylated during conditions of hunger.
Phospho-AS160, the active form of AS160, is an important downstream regulator of AMPK that triggers GLUT4 translocation from the intracellular pool to the plasma membrane for glucose transport. We observed that the baseline phospho-AS160 levels were higher in the obese Zucker rats than the lean rats from the control groups and inferred that this increase may be a result of the overexpression of phospho- AMPK, the AS160 upstream kinase.
According to our data, the lean Zucker rats of the exercise plus hypoxia group (altitude training group) had increased levels of AMPK and AS160 phosphorylation, lower fasting insulin and glucose levels, and increased GLUT4 expression compared with the lean controls. We speculate that the training-induced increases in the levels of AMPK and AS160 phosphorylation aided in the regulation of glucose absorption in the muscle and the maintenance of glucose homeostasis. The reduction in fasting insulin and glucose levels, along with the increase in GLUT4 protein expression in the muscle, provide evidence that altitude training may attenuate insulin resistance. These results supplement previous research on training-induced alterations in metabolic pathways.
In the muscles of the lean rats of the exercise group, we observed a significant increase in phospho-AS160 levels, compared to the lean controls. The phospho-AMPK and GLUT4 expression levels in this group showed an insignificant trend toward an increase, compared with the lean controls. This finding conflicts with past studies reporting that exercise training can significantly increase phospho-AMPK and GLUT4 expression in normal rats (Frosig et al., 2004; Langfort et al., 2003; Winder et al., 2006). These conflicting results may be due to the intensity of the exercise. Our progressive swimming-exercise training protocol may have been less intense than those used in past studies. However, in the lean rats of hypoxia group, there was an insignificant trend toward an increase GLUT4 protein expression, compared with the lean controls. This increasing trend agrees with the results from our previously published study, which showed that 4 weeks of 8-h/day of hypoxia increased GLUT4 expression in normal Sprague- Dawley rats (Chiu et al., 2004). Our present data reaffirms the effects of intermittent hypoxia on muscle GLUT4 expression.
A previous study showed that the commonly observed effects of exercise training on the normalization of AMPK phosphorylation levels are blunt in the skeletal muscle of genetically obese Zucker rats (Sriwijitkamol et al., 2006). The results of our study provide further support for the hypothesis that the combination of hypoxia and exercise training (altitude training), which is frequently used by endurance athletes, can lower the levels of red muscle AMPK and AS160 phosphorylation, partially improve the fasting insulin and glucose levels, and increase GLUT4 expression in the red muscles of obese Zucker rats. We also observed that, in the lean Zucker rats of the control group, the altitude training increased the levels of AMPK and AS160 phosphorylation in the red muscle, lowered fasting glucose and insulin levels, and increased GLUT4 protein expression. Several recent studies have shown that exercise training under hypoxic conditions may improve glucose metabolism and mitochondrial function in humans (Lecoultre et al., 2009; Schmutz et al., 2010). It would be interesting to investigate whether altitude training can be used as a therapy for humans with metabolic syndromes. In conclusion, the findings of our study provide strong evidence that lifestyle intervention can both prevent insulin resistance and correct metabolic conditions caused by genetic defects.
Footnotes
Acknowledgments
This study was supported by the V98C1-006 Grant from the Taipei Veterans General Hospital and the NSC 97-2314-B075-013-MY3 Grant from the National Science Council.
Author Disclosure Statement
No competing financial interests exist.