
Abstract
Abstract
Tissot van Patot, Martha C. and Max Gassman. Hypoxia: Adapting to high altitude by mutating EPAS-1, the gene encoding HIF-2α. High Alt. Med. Biol. 12:157–167, 2011.—Living at high altitude is demanding and thus drives adaptational mechanisms. The Tibetan population has had a longer evolutionary period to adapt to high altitude than other mountain populations such as Andeans. As a result, some Tibetans living at high altitudes do not show markedly elevated red blood cell production as compared to South American high altitude natives such as Quechuas or Aymaras, thereby avoiding high blood viscosity creating cardiovascular risk. Unexpectedly, the responsible mutation(s) reducing red blood cell production do not involve either the gene encoding the blood hormone erythropoietin (Epo), or the corresponding regulatory sequences flanking the Epo gene. Similarly, functional mutations in the hypoxia-inducible transcription factor 1α (HIF-1α) gene that represents the oxygen-dependent subunit of the HIF-1 heterodimer, the latter being the main regulator of over 100 hypoxia-inducible genes, have not been described so far. It was not until very recently that three independent groups showed that the gene encoding HIF-2α, EPAS-1 (Wenger et al. 1997), represents a key gene mutated in Tibetan populations adapted to living at high altitudes (Beall et al. 2010, Yi et al. 2010, Simonson et al. 2010).
Hypoxia-inducible transcription factors were first identified by the description of HIF-1 (Semenza et al. 1991, 1992), which was subsequently found to enhance transcription of multiple genes that encode proteins necessary for rescuing from hypoxic exposure, including erythropoietic, angiogenic and glycolytic proteins. Then HIF-2 was identified (Ema et al. 1997; Flamme et al. 1997; Hogenesch et al. 1997; and Tian et al. 1997) and although it is highly similar to HIF-1 and has the potential to bind (Camenisch et al. 2001) and mediate (Mole et al. 2009) many of the same genes as HIF-1, its biological actions in response to hypoxia are distinct from those of HIF-1 (reviewed by Loboda et al. 2010). By now, several of these HIF-2 mediated processes have been implicated in the human response to high altitude exposure including erythropoiesis (Kapitsinou et al. 2010), iron homeostasis (Peyssonnaux et al. 2008), metabolism (Shohet et al. 2007; Tormos et al. 2010; Biswas et al. 2010; Rankin et al. 2009) and vascular permeability (Chen et al. 2009; Tanaka et al. 2005), among others. Clearly, mutation of EPAS-1 has the potential to bring far more advantage when adapting to high altitude than solely mutating the Epo gene.
Introduction
The goal of this review is to highlight the biological processes in which both HIF-2 and human response to acute and chronic hypobaric hypoxia are implicated (Table 1). Focus is on the molecular pathways by which HIF-2 may be involved in acclimatization, adaptation or the failure of these processes that might lead to acute mountain sickness (AMS), high altitude pulmonary edema (HAPE), high altitude cerebral edema (HACE) or chronic mountain sickness (CMS). Because it has been demonstrated that renal HIF-2 is responsible for hypoxia-induced transcription of Epo, and the resulting erythropoiesis is an integral part of the human response to hypoxia, we begin with a brief history of the events that have led to the discovery of the pathway responsible for hypoxia-induced erythropoiesis. Subsequently, we describe the possible impact of HIF-2 on adaptation and acclimatization to high altitude in humans.
High Altitude, HIF-2, Epo and Erythropoiesis
Understanding of the hematopoietic response to acute high altitude exposure has eloquently unfolded over the past 122 years. In 1878, Paul Bert and his mentor Denis Jourdanet discovered the relationship between low pO2 and higher concentrations of red blood cells and hemoglobin while conducting studies on natives with chronic mountain sickness and animals in the highlands of Peru (Bert 1878). They proposed that the increased hematocrit was a familial trait inherited over generations. In 1890, Francois-Gilbert Viault traveled from France to Morococha, Peru (4500 m) and found polycythemia in his own blood and that of his companions following 23 days at altitude, concluding that low pO2 stimulated erythropoiesis (Viault 1890) and was not dependent on an inherited genetic factor. During an expedition to Everest in 1924, Somervell noted that the hemoglobin level in highly adapted Tibetan natives was much lower than that in members of the expedition team (Somervell 1925). Therefore, by 1925 it was known that hemoglobin and hematocrit were elevated in lowlanders ascending to high altitude, and in comparison native healthy highlanders generally had low hemoglobin and hematocrit at high altitude. It was also apparent that native highlanders who became ill at altitude had higher hemoglobin than lowlanders at high altitude.
At this same time, it was just beginning to be realized that there was a humoral factor that stimulated erythropoiesis during hypoxia (Carnot et al. 1906, Carnot et al. 1906, and Bonsdorff et al. 1948), but clear evidence of Epo's existence wasn't reported until 1953 (Erslev 1953). In 1966, Siri and colleagues (Siri et al. 1966) were the first to measure plasma Epo in humans following ascent to high altitude, reporting the now well-known response of an initial spike and then reduction to approximately twice sea level values. This was followed by data from Faura et al. in 1969 demonstrating that urinary Epo in Andean highlanders living at high altitude was low, similar to concentrations found in lowlanders at sea level. Interestingly, in these studies there were two highlanders with very high hematocrit (72-73%) and urine Epo (approx. 15–25 U/day) levels, who may have been developing CMS.
In 1991, the hypoxia-inducible transcription factor (HIF-1) was discovered by its ability to stimulate transcription of the Epo gene during hypoxia (Semenza et al. 1991 and 1992). For several years, it was assumed that the primary stimulus for Epo production in response to acute hypoxia was HIF-1. The discovery of a second hypoxia-inducible transcription factor termed HIF-2 in 1997, (Ema et al. 1997, Flamme et al. 1997 and Tian et al. 1997), led to further understanding of the hematopoietic response to hypoxia. In the years 2003 and 2004, data from in vivo (animal model, Morita et al. 2003) and in vitro (Warnecke et al. 2004) experiments suggested that despite the fact that HIF-1 was discovered by its ability to bind the hypoxic response element (HRE) of the Epo gene in response to hypoxia in human hepatoma cells, in vivo HIF-2 seems to be the primary mediator of Epo expression in kidneys in response to hypoxia. And indeed, over a decade after its discovery, Haase and colleagues (Kapitsinou et al. 2010) clearly demonstrated the primary role of renal HIF-2 in promoting the hypoxic renal Epo response. At this time, the only report involving investigations of HIF-2 at high altitude in human subjects was published by Robach and colleagues (Robach et al. 2007). They reported a 3.3 fold increase in HIF-2α mRNA in the skeletal muscle of human subjects following 7-9 days at 4559 m, compared with a modest 2.2 fold increase in HIF-1α mRNA. These results were associated with elevated Epo plasma levels. In a separate report on the same biopsies used by these authors (Viganò et al. 2008), no change in HIF-1α protein was found with ascent to altitude strongly supporting the notion that HIF-2 had been responsible for the rise in Epo plasma levels.
The most recent discovery, that alterations (SNPs) in EPAS-1 are unique to native Tibetan highland populations (Beall et al. 2010; Yi et al. 2010; and Simonson et al. 2010) and are associated with lower hemoglobin concentrations (Beall et al. 2010), has added a new chapter to the hematological response to high altitude. This observation provides convincing evidence for HIF-2α's role in the adaptation of native populations to high altitude. However, many questions remain regarding the role of HIF-2α in the different erythropoietic response to high altitude between lowlanders, Tibetans, Andean populations and non-native highlanders who develop CMS. To visualize these differences, data from different reports have been depicted in Figure 1 that shows hematocrit (Hct), hemoglobin (Hb) and Epo levels in these populations. The most intriguing question is whether HIF-2 regulates the hematological response to acute high altitude in lowlanders. Moreover, as South American highlanders show a much higher incidence of CMS (Leon-Velarde et al. 2005), one might speculate that the described EPAS-1 mutations prevent most Tibetans from CMS. Accordingly, the question arises if manipulation of the HIF-2 and/or Epo pathway might be feasible to manipulate or treat CMS.
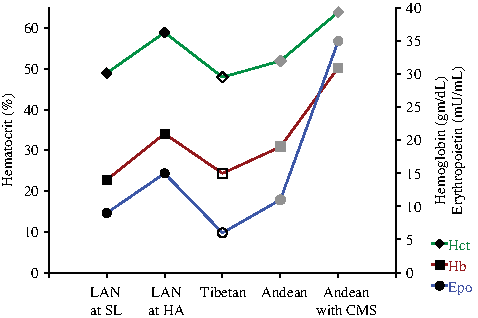
Previously reported hematocrit (Hct), hemoglobin (Hb), and plasma erythropoietin (Epo) values for low altitude natives (LAN) from India at sea level (SL) and high altitude (HA) (Basu et al. 2007), Tibetans and Andeans at HA (Beall et al, 1998; Winslow et al. 1989 and Andeans suffering from chronic mountain sickness (CMS) at HA ( Leon-Velarde, Villafuerte, & Richalet 2010; Gonzales et al. 2009). Note that the erythropoietic values between LAN at SL and Tibetans at high altitude are similar. Black markers: populations without EPAS-1 mutations (e.g. no SNPs); open markers: populations with mutations in the EPAS-1 gene; grey markers: populations in which EPAS-1 SNP's have not (yet) been reported.
Of note, apart from its role in regulating red blood cell production, Epo has been shown to be expressed in brain (Marti et al. 1996, Digicaylioglu et al. 1995) and retina (Grimm et al. 2002) and to protect these and other organs from injuries (Gassmann et al. 2003). Apart form its protective function against pathological events such as stroke (Wiessner et al. 2001), Epo has been found to enhance the neuronal control of the ventilatory response to hypoxia (Gassmann et al. 2009, Soliz et al. 2005). The question arises as to whether the control of cerebral Epo expression is mediated—in analogy to the kidney—by HIF-2. If so, one would expect altered Epo levels in the brain of Tibetans when exposed to reduced oxygenation. Thus, investigations further delineating the role of the HIF-2/EPO pathway will likely contribute valuable information to the understanding of human response to acute and chronic hypoxia.
High Altitude, HIF-2 and Iron Homeostasis
In order to fully understand erythropoiesis at high altitude, it is crucial to gain an understanding of iron homeostasis under these conditions. In 1959, Reynafarje et al. reported a 3-fold rise in intestinal absorption of iron in lowlanders ascending to high altitude, but normal intestinal absorption in native highlanders. Shortly after, Hornbein demonstrated that the increased iron absorption appeared to be sufficient to support erythropoiesis during high altitude ascent without needing to supplement dietary iron (Hornbein 1962). However, this study was performed in subjects with unlimited access to food containing sufficient iron content. Note that studies from Bolivia suggested that mothers and children living at high altitude could not consume enough iron to meet the hemoglobin demand (Berger et al. 1997), however it was subsequently shown that the low iron in these subjects was due to reduced food availability and not altitude exposure (Cook et al. 2005).
Because HIF-2 plays a key role in red blood cell production, the latter requiring a considerable amount of iron, it was expected that HIF-2 would also have an impact in iron homeostasis. Indeed, recent evidence indicates that intestinal HIF-2 promotes the uptake of iron from nutritive sources via transcriptional control of the genes encoding duodenal cytochrome b (DcytB) and divalent metal transporter 2 (DMT) (Mastrogiannaki et al. 2009 and Shah et al. 2009). DcytB reduces dietary ferric iron (Fe3+) to ferrous iron (Fe2+) and then DMT absorbs Fe2+ at the apical membrane in the intestinal brush border (reviewed by Shah et al. 2009). During normoxia, HIF-2α expression is inhibited by the binding of the iron regulatory protein a (IRP1) to an iron responsive element (IRE) in the 5’-untranslated region of the HIF-2α message (Zimmer et al. 2008). Hypoxia causes repression of the IRP1 – IRE binding, releasing HIF-2α to be translated and allowing the HIF-2 mediated hypoxic response (Zimmer et al. 2008). Taken together, ascent to high altitude increases iron demand in association with elevated erythropoiesis, and it is possible that HIF-2 stimulates increased dietary iron absorption. At present, there is little evidence, however, to suggest that highlanders have higher dietary absorption of iron as compared to lowlanders, although this aspect needs further investigation (Zhou et al. 2006). Interestingly, the increase in dietary iron consumption with acute ascent to high altitude may not be sufficient to fuel the increased erythropoiesis, as suggested by a recent report in which myoglobin content of human skeletal muscle was reduced following ascent to high altitude (Robach et al. 2007).
High Altitude, HIF-2 and Pulmonary Hypertension/Remodeling
Chronic exposure to high altitude can cause the development of pulmonary hypertension and remodeling of the pulmonary vasculature involving smooth muscle cell proliferation (Maggiorini et al. 2003 and Alexander et al. 1963). Not unexpectedly, HIF-2 has been implicated in pulmonary hypertension and in smooth muscle cell proliferation (Diebold et al. 2010 and Brusselmans et al. 2003). It is of interest to note that the recent finding that EPAS-1 is mutated in Tibetans, but not low altitude Han Chinese population corresponds well with the fact that pulmonary artery pressure is higher in the Han as compared with the Tibetans at high altitude. Furthermore, Tibetans have lower pulmonary artery pressure at high altitude than native Andean highlanders and North Americans, as well as a lower hypoxic pulmonary vascular response (reviewed by Maggiorini et al. 2003).
High Altitude, HIF-2 and Metabolism
In response to acute hypoxic exposure, human metabolism is characterized by increased glycolysis and elevated lactate production (reviewed by Hoppeler et al. 2003). Mitochondrial response to hypoxia has been a controversial subject with various investigators demonstrating increased (Schmutz et al. 2010) or unchanged (Lundby et al. 2004) mitochondrial concentration in skeletal muscle from human subjects exposed to high altitude. However, there is also significant evidence that mitochondrial function is altered with ascent to high altitude (reviewed by Hoppeler et al. 2003). Naturally, reduced oxygen delivery to cells initiates a change in metabolism, presumably to preserve life of the cell and organism. A complex series of events begins in which reactive oxygen species (ROS) are produced by the hypoxic mitochondria and stimulate the activation of HIF-1 and HIF-2. In turn, HIF-1 mediates the initiation of glycolysis (Hu et al. 2003) and reduces mitochondrial respiration (reviewed by Tormos et al. 2010), while HIF-2 directly reduces mitochondrial production of ROS and obviously preserves mitochondrial homeostasis (Kojima et al. 2007 and Scortegagna et al. 2003). HIF-2 reduces mitochondrial ROS production by promoting transcription of genes encoding anti-oxidants superoxide dismutase 1 (SOD1), manganese SOD 2 (SOD2), glutathione peroxidase type 1 (GPX1) and catalase, with SOD2 appearing to be the primary mediator of reduced mitochondrial ROS (Kojima et al. 2007 and Scortegagna et al. 2003). HIF-2 also protects the respiratory control ratio through a complex pathway resulting in the preservation of Krebs tricarboxylic acid (TCA) cycle activity (Oktay et al. 2007). Specifically, HIF-2 promotes transcription of the gene for frataxin, a protein that protects stores of iron-sulfur cluster from disassembly during oxidative stress. The iron-sulfur cluster is necessary for activation of the mitochondrial aconitase enzyme that in the TCA converts citrate to isocitrate cycle (Oktay et al. 2007 and Bulteau et al. 2004). Thus HIF-2 is instrumental in protecting the cells from mitochondrial dysfunction during hypoxic exposure.
Lipid metabolism is also increased following chronic exposure to high altitude although the exact mechanisms by which this is mediated are unknown (Ou et al. 2004 and McClelland et al. 2001). Recently, HIF-2 has been implicated in lipid metabolism in the liver, specifically associated with impaired fatty acid β-oxidation, reduced lipogenic gene expression and elevated lipid storage capacity (Kapitsinou et al. 2010), suggesting HIF-2 may also play a role in lipid metabolic response to high altitude exposure.
High Altitude, HIF-2 and Vascular Permeability
Rapid ascent to high altitude is associated with elevated risk for the development of AMS, HAPE, and HACE, all of these diseases being associated with leaking capillaries (Maggiorini et al. 2001; Schoene et al. 1988). In response to hypoxia vascular endothelial growth factor (VEGF) increases vascular permeability as a first step in angiogenesis (Issbrucker et al. 2003) and can be mediated by HIF-1 and HIF-2 (Ema et al. 1997 and Forsythe et al. 1996). Initial investigations reported that plasma VEGF concentrations in groups of subjects acutely ascending to high altitude were no different from that of subjects at sea level (Walter et al. 2001 and Maloney et al. 2000). However, a subsequent study in which individual variation was controlled by having each subject serve as his/her own sea level control reported a significant rise in plasma VEGF in subjects who developed AMS, but not in healthy subjects (Tissot van Patot et al. 2005). Soluble VEGF receptor-1 (sFlt-1) can be mediated by HIF-1 or HIF-2 (Kendall et al. 1996 and Eubank et al. 2010) that can bind circulating VEGF and prevent its induction of capillary leak (Hastings et al. 2003). Therefore, investigators also determined plasma sFlt-1 concentration in the same subjects. Healthy subjects had a greater rise in plasma sFlt-1 than those who developed AMS. This is especially relevant in light of new evidence that HIF-1 preferentially increases VEGF, while HIF-2 preferentially induces transcription of sFlt-1 in response to acute hypoxia (Eubank et al. 2010). It is tempting to hypothesize that a strong HIF-2 response may be important for acclimatization to acute ascent, as opposed to the genetic evidence suggesting that prolonged HIF-2 activity in high altitude residents may not be beneficial for adaptation.
Gene Conservation: HIF-1 vs. HIF-2
There is 90% conservation of the amino acid sequence and 80% conservation of the nucleotide sequence in the proximal promoter of the genes encoding human and mouse HIF-1α subunits (Iyer et al. 1998). Comparison of the human α–subunits between HIF-1 and HIF-2 reveals significant differences in splice site locations, the location of each of their 14 introns (Iyer et al. 1998 and Tian et al. 1997). In lieu of recent data demonstrating genetic variation in EPAS-1 resulting in phenotypic differences (Yi et al. 2010; Beall et al. 2010; Bigham et al. 2009; Percy et al. 2008), it is possible that the gene encoding HIF-1α gene was replicated at some point in evolution giving rise to EPAS-1 and HIF-2α. Such gene replications might allow, in turn, greater plasticity of the organism in the hypoxic response.
Mutations (SNPs) in EPAS-1
Few functional mutations of human EPAS-1 have been reported, yet those that have been identified are all associated with changes in hemoglobin and circulating red blood cell concentration (Yi et al. 2010; Beall et al. 2010; Bigham et al. 2009; Percy et al. 2008). The first report of a mutation in EPAS-1 described a gain of function mutation in a family with erythrocytosis and high serum hemoglobin concentrations (Percy et al. 2008). This report was followed by a description of a mutation in EPAS-1 linked with erythrocytosis and pulmonary hypertension (Gale et al. 2008), both of which are associated with high altitude exposure (reviewed by Maggiorini et al. 2003 and Monge et al. 1991). Another gain of function mutation of EPAS-1 was reported in a family with erythrocytosis, and in association with elevated mRNA for Epo receptor, VEGF, transferrin receptor, adrenomedullin and N-myc downstream regulated gene 1 (Martini et al. 2008). More recently, a very minor amino acid substitution in the human EPAS-1 resulted in a gain of function mutation associated with erythrocytosis (van Wijk et al. 2010). Thus, there are four reports of a mutation in EPAS-1 associated with clinical cases of erythrocytosis. This makes the current reports of mutated EPAS-1 in the Tibetan population unique in the fact that these mutations are associated with loss of function or low hemoglobin concentrations in healthy individuals described very recently (Yi et al. 2010; Beall et al. 2010; Bigham et al. 2009). Note that not all Tibetans carry the mutated EPAS-1 gene, which may explain why CMS is present, albeit with low incidence, also in the Tibetean population (Wu, et al. 1998).
While the erythropoietic effects of EPAS-1 mutation are easily noted, there may be other effects of the mutations that are not as clearly determined. Evidence to support this hypothesis comes from the association of pulmonary hypertension (Gale et al. 2008) and elevated mRNA encoding hypoxia-mediated proteins (Martini et al. 2008) in human patients. Further evidence of more far reaching effects of EPAS-1 mutation comes from animal and cell culture studies.
Mutating EPAS-1 in mice and cells
Initial attempts to knock out EPAS-1 in mice resulted in embryonic lethality due to inhibition of catecholamine activation in mid-gestation (Tian et al. 1998) and impaired angiogenesis (Peng et al. 2000). However, Scortegagna et al. (2003) succeeded in breeding a small number of surviving HIF-2α null mice whose phenotype included hypoglycemia, lactic acidosis, altered Krebs cycle function and dysregulated fatty acid oxidation. It was hypothesized that these phenotypes were the result of the reduction in anti-oxidant enzymes present in these animals. Recent studies in MCF-7 breast cancer cells revealed that HIF-1 and HIF-2 bind to the same core-binding motif (RCGTG) with similar affinities and therefore have the potential to mediate transcription of the same genes (Mole et al. 2009). However, acute hypoxia primarily induced HIF-1 binding and not HIF-2. There is ample evidence to suggest that HIF-2 responds more effectively to prolonged hypoxia, while HIF-1 is an acute responder (proposed in (Patel et al. 2008)). This is counter-intuitive in light of recent evidence for EPAS-1 mutation in Tibetan highlanders, however an intact EPAS-1 gene may be beneficial for acclimatization, but suboptimal for adaptation to high altitude.
Transcriptional Co-Activators Specifically Involved in HIF-2 Activation
Although more definitive proof is needed, there are a few proteins that have been linked specifically to HIF-2 activation, and not HIF-1 (Table 2). For example eukaryotic translation initiation factor 3 subunit E (eIF-3 p48), encoded by EIF3E (also known as INT6), interacts specifically with HIF-2α, resulting in proteosome-dependent degradation (Chen et al. 2007). This is an oxygen and pVHL independent degradation process and when this interaction is inhibited there is a subsequent induction of angiogenic proteins angiopoietin, fetal liver kinase 1, Tie-2, and VEGF, stronger than that mediated by HIF-1. Prolyl hydroxylase inhibitor 3 (PHD-3) is appears to preferentially bind HIF-2α, targeting it for proteosomal degradation (Appelhoff et al. 2004). Possible co-factors that appear to be necessary and specific for HIF-2 DNA-binding include ETS-1 (p54), ELK-1 and NEMO (Hu et al. 2007; Aprelikova et al. 2006; Bracken et al. 2005; Elvert et al. 2003). Recently, a hypoxia associated factor (HAF) has been demonstrated to inhibit HIF-1 activation by promoting HIF-1α ubiquitination and simultaneously promote HIF-2 transactivation in response to prolonged hypoxia in tumor cell cultures (Koh et al. 2011). Take together, one might speculate that mutations in EPAS-1 in Tibetans adapted to high altitude are associated with mutations of those genes encoding the transcriptional co-factors or expression of the co-factors of HIF-2 and thus may be useful in marker specific gene analyses in altitude studies.
Parentheses indicate the common abbreviation or alternate name for the protein or gene.
Caveats
There are some caveats to this review that we would like to emphasize. HIF-2 plays a critical role in embyrogenesis, specifically with regard to angiogenesis (reviewed by Patel et al. 2008), and in tumorigenesis (reviewed by Pietras et al. 2010). These topics are beyond the scope of this review; however it is quite likely that the role of HIF-2 during development is of critical importance to high altitude adaptation. Another important consideration is that the data discussed is from a wide variety of studies emphasizing human subject research, but also including studies in animal and cell models. As emphasized in a recent review on metabolomics and hypoxia (Serkova et al. 2008), great care must be taken when considering responses to hypoxia from differing cell types, organs and species. The information presented in this review is meant to present strong associations between the human global response to hypoxia and the extensive biologic systems in which a role for HIF-2 has been implicated. Despite only being mentioned now, one should keep in mind that epigenetic aspects regulating the cell's oxygen sensing mechanism, including HIFs and PHDs, might represent a major player in adaptation to high altitude. In summary, this review highlights how the EPAS-1 gene is central to physiology. Careful functional dissection of the Tibetan adaptation provides an opportunity to examine in detail how the SNPs in the Tibetan allele have managed to tweak the spectrum of this regulation without otherwise harming this tough and physically active population.
Footnotes
Disclosures
Drs. Tissot van Patot and Gassmann have no conflicts of interest or financial ties to disclose.