
Abstract
Abstract
Külköylüoğlu, Okan, Necmettin Sari, Derya Akdemir, Mehmet Yavuzatmaca, Ceren Altinbağ. Distribution of sexual and asexual ostracoda (crustacean) from different altitudinal ranges in the Ordu region of Turkey: Testing the Rapoport Rule. High Alt. Med. Biol. 13:126–137.—We evaluated Rapoport's rule which states a negative correlation between species richness and altitude. To understand the relationship between altitude and reproductive modes (a/sexual) of non-marine ostracods, 166 aquatic bodies in Ordu region, Turkey were randomly sampled from July 11 to July 16, 2010. A total of 26 species of ostracods were found from 133 out of 166 sites. Except for one species (Heterocypris incongruens), the other 25 species were new reports for the region. Candona improvisa was also a new report for Turkish ostracod fauna. Three species (Psychrodromus olivaceus, H. incongruens, and C. neglecta) occurred most frequently as 43, 46, and 76 times, respectively. Canonical correspondence analyses exhibited two variables [Habitat type (p=0.014; F=2.171) and water temperature (p=0.018; F=2.248)] as having the most effect on species. Correlation of species' reproductive modes to those of environmental variables measured was not significant. UPGMA dendrogram displayed 15 most frequently occurring species into four clusters where most species (11) were asexual. Although a small group (asexual species without swimming setae) showed a tendency to habitat type and electrical conductivity, such variables are believed to play secondary role on species distribution. Highest species diversity (13 species) was observed at the range of 1200 and 1400 m (a.s.l.), where numbers of stations sampled was not the highest (22). Numbers of asexual species (19) were higher than the sexual (11) but there were no significant differences (p>0.05) between the frequencies of their occurrences at different altitudinal ranges. Accordingly, our findings do not support the Rapoport Rule. Results yield that reproductive modes of species (sexual and asexual) was not directly correlated to altitude or any environmental variables measured during this study. A better explanation of ostracod diversity appears to be suitability of habitats.
Introduction
The relationship between reproduction and altitude has been the subject of a long-standing question. Reproduction and reproductive characteristics of organisms at high altitudes have been studied in different taxonomic groups, including humans (Crognier et al., 2002; Okumura et al., 2003), waterbirds (Lu, 2011), and plants (Hautier et al., 2009; Zhao et al., 2007). Further, Zhao et al. (2010) showed that biomass was correlated with altitude for various zooplankton and phytoplankton. However, almost nothing is known about ostracod reproduction modes at different altitudinal ranges (but see Mezquita et al., 1999; Külköylüoğlu et al. (2012)). However, contemporary studies show that increasing altitude does not reduce or limit ostracod distribution and diversity (Külköylüoğlu et al., 2012; Laprida et al., 2006). Similar results have already been determined in other taxonomic groups. For example, the study of Wettstein and Schmid (1999) on grasshopper diversity in wetlands of Switzerland exhibited declining diversity with high altitude. In contrast, lichen diversity in northeast India were generally more diverse in the mid altitudinal ranges (Pinokiyo et al., 2008).
Rapoport's Rule states that species diversity and altitude are negatively correlated, meaning that species richness should decrease with increasing elevation (Fleishman et al., 1998; Stevens, 1992). Although the rule has been argued with many studies since its introduction (Bhattarai and Vetaas, 2006; Colwell and Hurt, 1994; Gaston et al., 1998; Rohde, 1992; Veech, 2000), it has not been tested with ostracods in Turkey. Thus, the aims of this study are: (a) to determine ostracod reproductive modes at different altitudinal ranges in Ordu region, Turkey; (b) to document ostracod diversity and distribution along with their ecology; and (c) to evaluate if there is/are any correlation(s) between ostracod reproductive modes and altitudinal distribution along with testing Rapoport rule.
Materials and Methods
Site description and sampling
The city of Ordu is located on the Black Sea coast in northern Turkey. Its surface area covers about 5963 km2, including 18 towns. There has been no extensive sampling from the area so far, except that Kılıç (2001) reported two ostracods (Cypridopsis vidua from Çınarsu stream and Cyprinotus inaequivalvis (now Heterocypris incongruens) from Ünye.
Ostracod samples were randomly collected from 166 aquatic sites from 15 of 18 towns during July 11 to July 16, 2010 (Fig. 1). Samples (ca. 200 ml) were collected with a handmade plankton net (125 μm in mesh size) from about 100 cm of depth and fixed in 70% ethanol in situ. In laboratory, materials were sieved using three standardized sieves (0.125, 1.00, and 2.00 mesh size). Ostracods were separated from debris under a stereomicroscope (Olympus BX51) and fixed in 70% ethanol for further analysis. Ostracod species were identified using the keys provided by Meisch (2000), but some other related works were also consulted.
166 sampling sites in the Ordu region of Turkey sampled during this study.
Physicochemical variables of sampling sites were measured before ostracod sampling. pH was measured using a Hanna HI-98150 pH/ORP meter; dissolved oxygen, temperature, and conductivity were measured with a YSI-85 oxygen-temperature meter, while GPS 12 XL was used to report data on altitude and coordinates.
Statistical analyses
Canonical Correspondence Analyses (CCA) with Monte Carlo test (499 permutation) after DCA (length of gradient>4) was used to show relationships between species' reproductive modes (i.e., bisexual species with males and females; parthenogenetic species with females only) and environmental variables by means of using CANOCO program (Ter Braak, 1987). Sampling sites were divided into five habitat types, such as lake, pond, creek, spring, and trough (Table 1). A t-test with different variances was applied to detect differences between the frequency of sexual and asexual species at different altitudinal ranges (Excel version 2003–2007). Unweighted pair group mean averages (UPGMA) were used to show clustering association among species (MVSP version 3.1, Kovach, 1998).
SHE (Redox Potential, mV), EC (Electrical Conductivity, μS/cm), TDS (Total Dissolved Solids, mg/l), TW (Water temperature, °C). Species codes: Candona candida, CC; C. improvisa, CI; C. neglecta, CN; C. sanociensis, CS; Candona sp., Cas; Cyclocypris ovum, COv; Cypria ophthalmica, CO; Darwinula stevensoni, DS; Eucypris virens, EV; Eucypris sp., Eus; Fabaeformiscandona breuli, FB; Herpetocypris brevicauda, HB; H. intermedia, Him; Heterocypris incongruens, HI; H. reptans, HR; Herpetocypris sp., Hrs; Heterocypris sp., Hes; Ilyocypris bradyi, IBr; I. gibba, IG; I. inermis, II; Ilyocypris sp., IIs; Potamocypris fallax, PF; P. fulva, PFu; P. pallida, PP; P. similis, PS; P. unicaudata, PU; P. villosa, PVi; P. zschokkei, PZ; Potamocypris sp., Pos; Prionocypris zenkeri, PZe; Psychrodromus olivaceus, PO; P. fontinalis, PFo; Psychrodromus sp., Psys; Pseudocandona sp., Psc. Note that
Results
A total of 26 species were found from 133 out of 166 sites (Table 1), of which, 25 species are newly reported for the area, while Candona improvisa is a new species for Turkish ostracod fauna. Maximum numbers of species was five when the mean numbers of species per site was 1.93. Psychrodromus olivaceus, H. incongruens, and Candona neglecta occurred most frequently, being observed at 43, 46 and 76 stations, respectively. CCA results did not correlate species' reproductive modes to certain environmental variables measured. Among four variables [habitat type (p=0.014; F=2.171); water temperature (p=0.018; F=2.248); pH (p=0.066; F=1.588); EC (p=0.088; F=1.577)], the first two had more influence on species than the others (Fig. 2a). Although a small group (asexual species without swimming setae) showed a tendency to habitat type and electrical conductivity (Fig. 2b), such variables are believed to play a secondary role on species distribution (see discussion). Cumulative percentage variation of the relationship between species and environmental variables was about 38.6%, while the first axis of CCA was able to show about 57% of the correlation (Table 2).
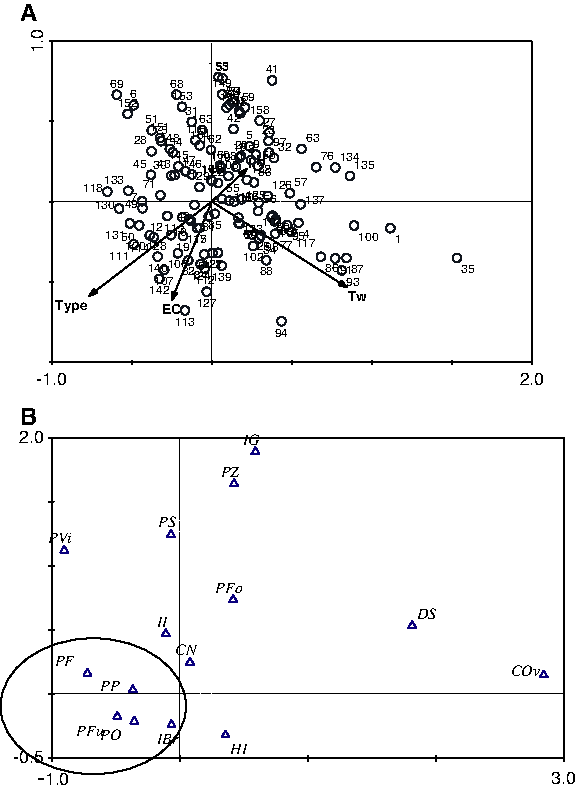
CCA diagram of
Species richness was highest (13 species) at the range of 1200 and 1400 m (above sea level) where numbers of stations was 22 (Fig. 3). Numbers of asexual species (19) were higher than the sexual (11) but there were no significant differences (t-test=0.88, p>0.05) between the frequencies of their occurrences at different altitudinal ranges (Fig. 4). Most of the asexual species were found without swimming setae, sexual species tend to have swimming setae (Fig. 5). According to UPGMA results, 11 of 15 most frequently occurring species were asexual without swimming setae (Fig. 6). Results showed that reproductive modes of species (sexual and asexual) were not significantly correlated to altitude or any of the other environmental variables measured during this study. Overall, a better explanatory factor on ostracod diversity and distribution seems to be suitability of habitats. Therefore, our findings do not support the Rapoport Rule where it is assumed that increasing altitude reduces the species diversity.
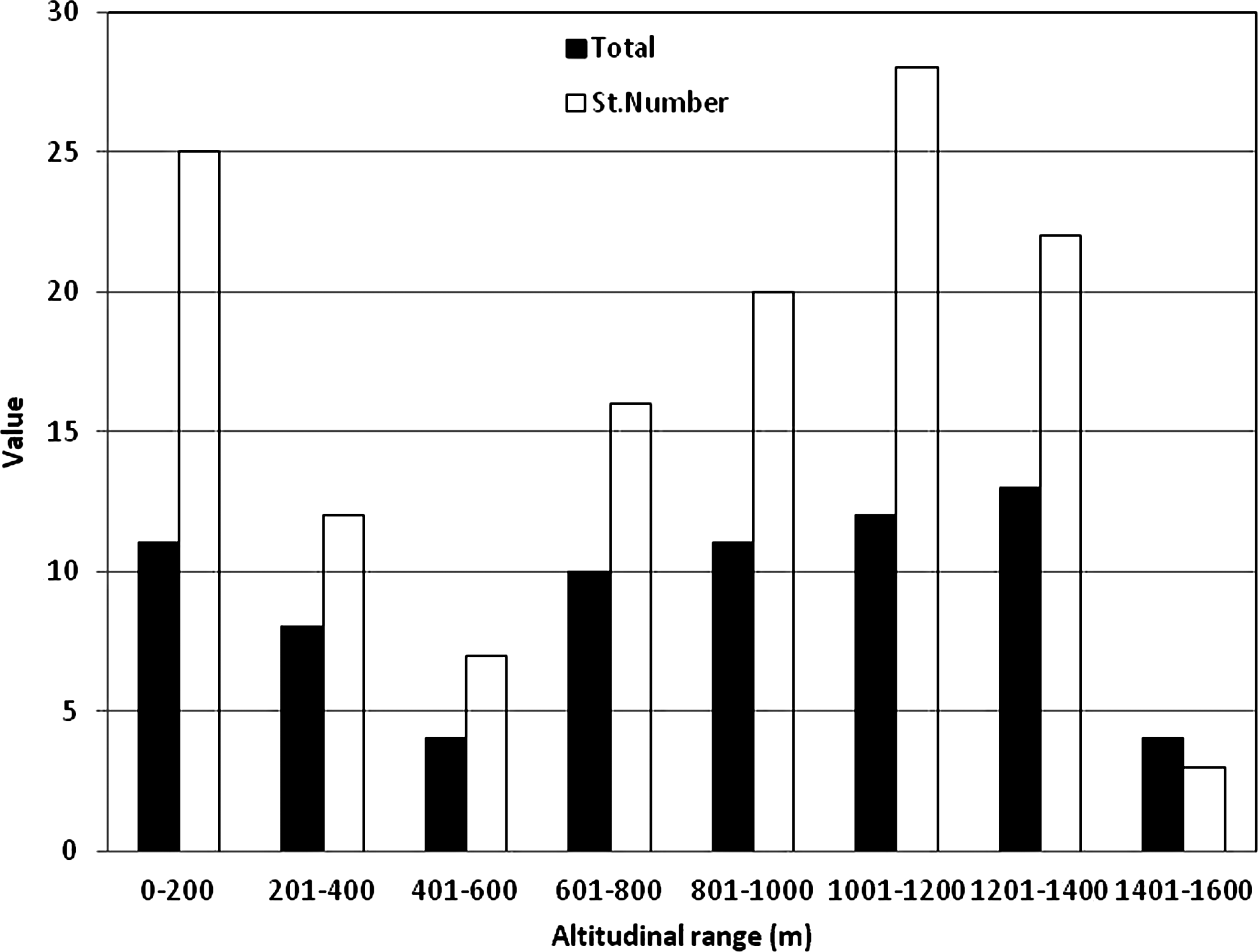
Highest species diversity (13 species) was observed at the range of 1200 and 1400 m, although number of stations sampled was not the highest (22).
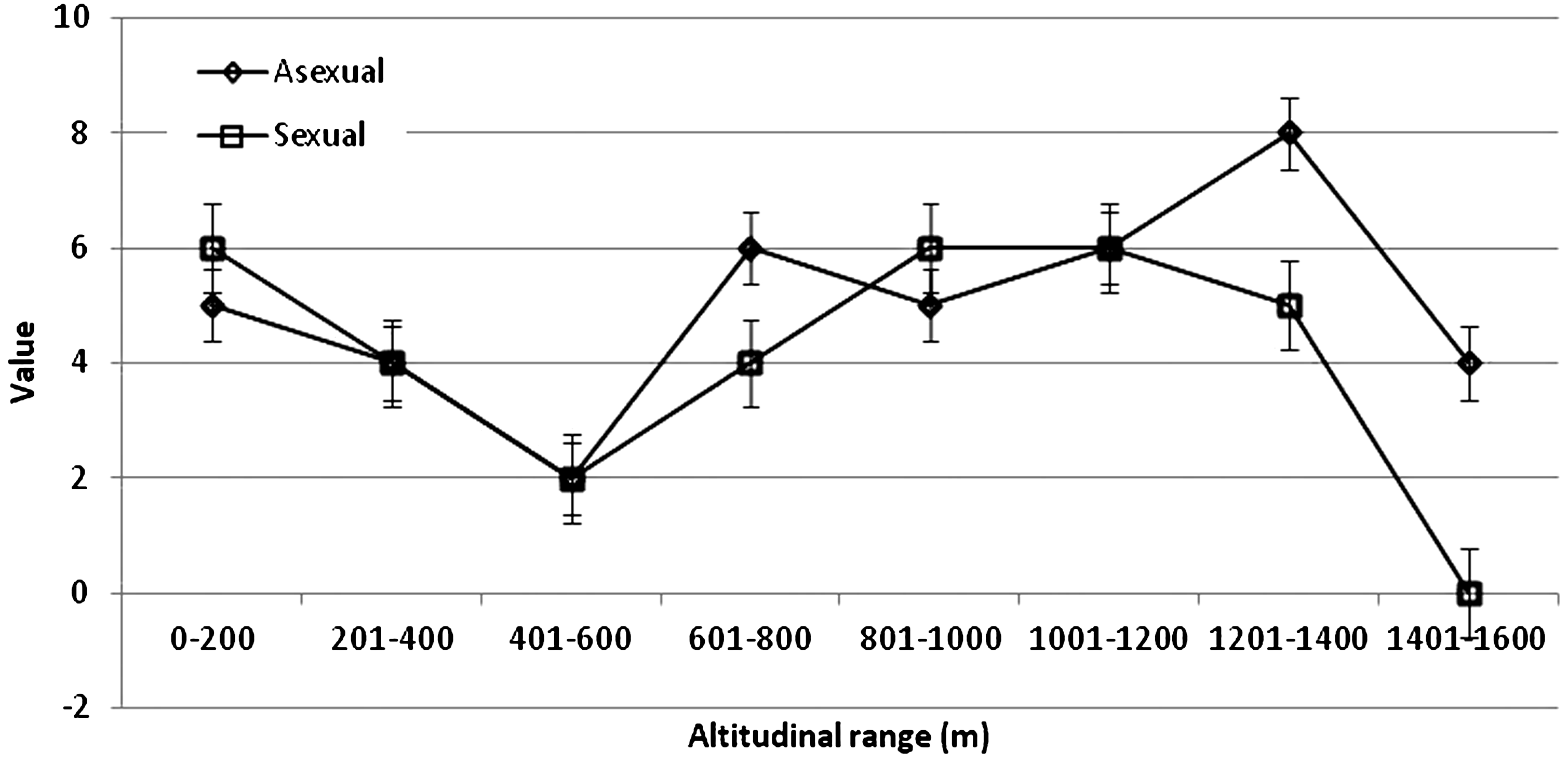
Numbers of sexual and asexual species from different altitudinal ranges.
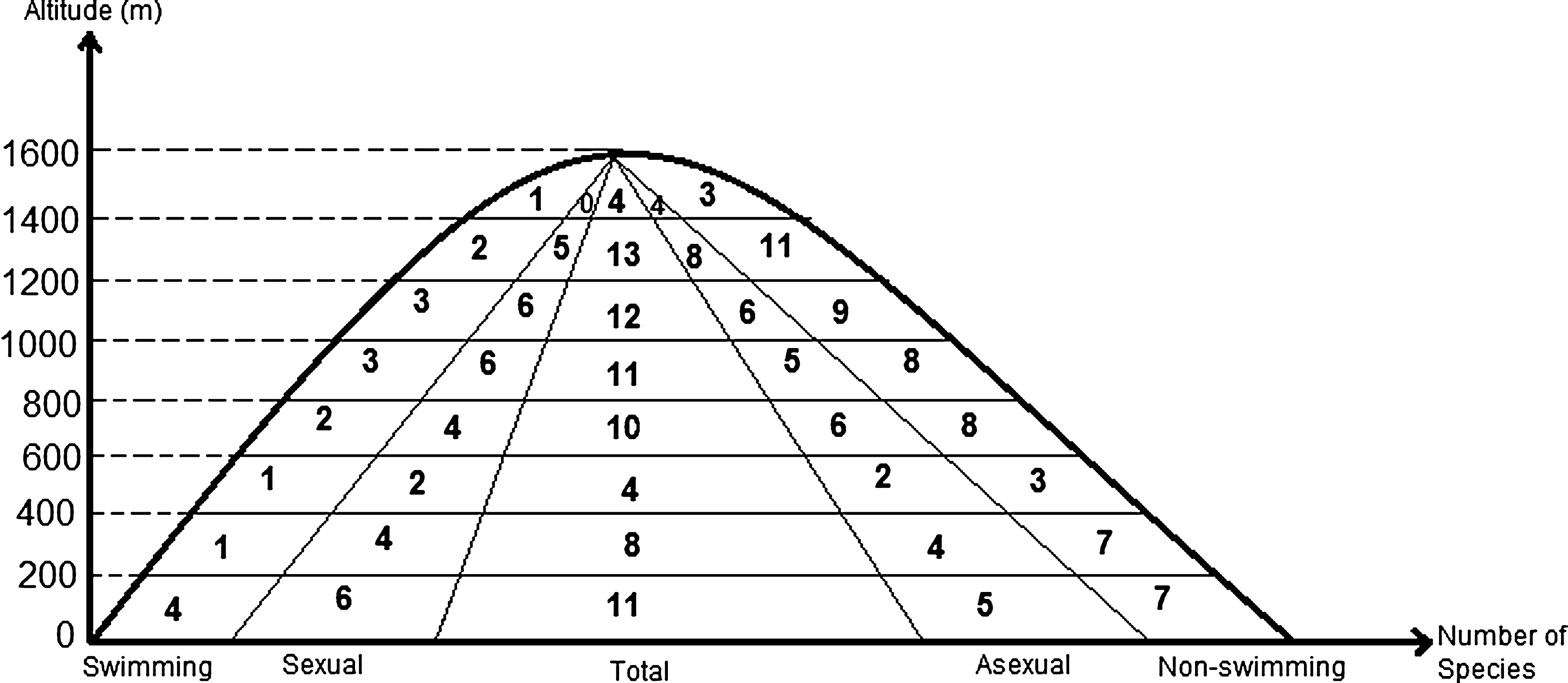
Numbers of species with and without swimming setae found from different altitudinal ranges (0–1600 m).
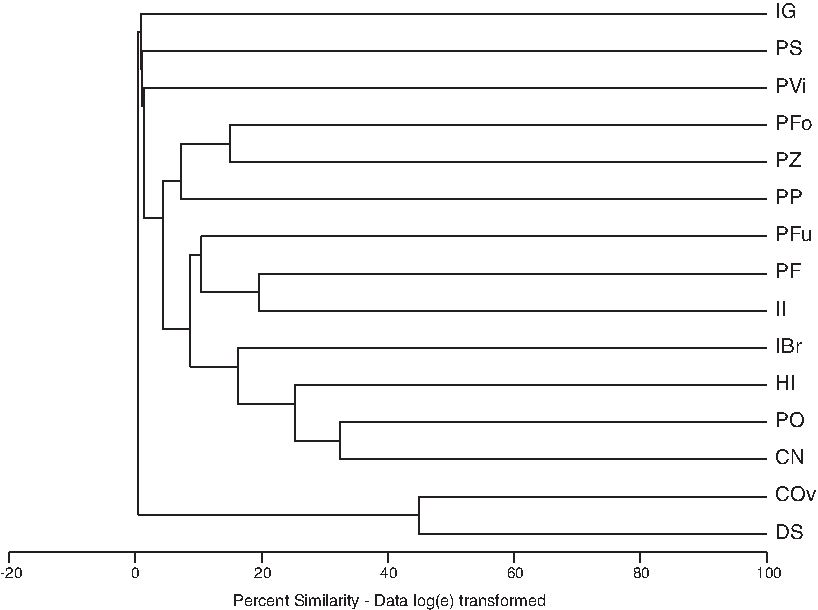
UPGMA showing no clear groupings among 15 species occurred at least two times. However, most species (11) showed asexual characteristics without swimming setae.
Discussion and Conclusion
Until recently, there was no detailed information about the ostracod fauna of the Ordu region, Turkey, except that a single study published by Kılıç (2001) reported two species (H. incongruens and C. vidua). The present study reports 25 species previously not observed in this area. Because our study was only about 7 days and covered several different types of habitats, this area is likely to have high ostracod species richness (Table 1). This is especially true at the range of 1200 and 1400 m where the highest species number (13) was accounted from 22 sites. Most recently, Külköylüoğlu et al. (2011) found a total of 29 species from 57 out of 78 sites at high elevational ranges (1659–2889 m) in the Northeastern part of Van region (Turkey) where 26 species were encountered from 38 sites between 1659 and 1750 m.
Additionally, Külköylüoğlu and Sarı (2012) reported 40 ostracod taxa from 141 different sites in Bolu (Turkey) while 62 and 54 species were collected from 156 and 48 sites (with 132 samples) of Belarus (Nagorskaya and Keyser, 2005) and Brazil (Higuti et al., 2010), respectively. Compared to the earlier studies, the mean numbers of species per site (1.93 spp.) in Ordu was lower than the mean numbers of species (13.2) per lake in Turkey (Külköylüoğlu, 2005) and species (2 spp.) from springs of Spain (Mezquita et al., 1999), but higher than species in springs of North America (0.58 spp.) (Külköylüoğlu and Vinyard, 2000) and wetlands of Northern Italy (1.19 spp.) (Rossetti et al., 2004).
Distribution of the asexual species (19 spp.) were also higher than the sexual (11 spp.) but there were no significant differences (p>0.05) between the frequencies of their occurrences at different altitudinal ranges. We did not find any clear correlation between reproductive modes of species (sexual and asexual) and altitude or any of the other environmental variables measured during this study. There can be several biotic and abiotic reasons for such differences in the mean numbers of species including the differences in habitat types, sampling time, water quality, occurrence of competition and/or predation. Unique structure of the habitats can be related to sexual modes of species. For example, when most of the asexual species (11 of 15) were found without swimming setae, sexual species tend to have swimming setae (Fig. 5). Using swimming setae may have advantages in finding food, mate, and movement in dynamic and unstable systems such as flowing waters. However, in small habitats with relatively stable conditions (e.g., troughs and springs), the need for swimming setae may not be necessary. Therefore, swimming setae may be advantageous for sexual ostracods of their higher energy demands and mate searching. Although it was claimed that most ostracods reproduce sexually (Cohen and Morin, 1990), in freshwater (nonmarine) ostracods, parthenogenesis appears to be exceptionally common (Schön and Martens, 1998) but the reason(s) is still unknown. In contrast, some of the ongoing studies and unpublished data exhibited that numbers of parthenogenetic and bisexual ostracods in freshwater habitats may not be very different from each other. For example, among ca. 159 species described from freshwater environments of western and central Europe (Meisch, 2000), there are 96 bisexual species (males present), 46 parthenogenetic (males absent), and about 17 species having with both males and females reported separately from different countries. Unlike earlier arguments, numbers of bisexual species are at least twice of the parthenogenetics. Similar patterns are also known from freshwater ostracods of Turkey where there are about equal numbers of bisexual and parthenogenetic populations (ca. 50 species each) and about 20 species with both reproductive modes (Külköylüoğlu, unpublished data). Besides, reporting males from different aquatic bodies and locations may imply that some species can have two different reproductive populations and/or mixed populations (Butlin et al., 1998). Indeed, several examples of those kinds of populations/species (Potamocypris villosa, Cavernocypris subterranean, and Eucypris virens) have already been reported in the literature (Külköylüoğlu and Vinyard, 1998; Martens and Meisch, 1985; Meisch, 2000, respectively). In a classical view, such differences in reproductive (or sexual) modes can be dependent on geographical and/or ecological isolation (Mayr and Ashlock, 1991). For example, Eucypris virens has parthenogenetic populations in northern Europe, but its bisexual populations are known from southern parts of Europe; mixed populations are also reported some areas in the middle (Horne et al., 1998; Martens et al., 1998). Most recently, Martins et al. (2009) confirmed the sympatric coexistence of both reproductive modes of E. virens in Spain and Poland with wide ecological tolerance. According to these authors, a/biotic factors may be important for changes in population dynamics of the species. For example, when males were present, the initial female-biased sex ratio was altered towards a balanced sex ratio by season. Implication of this study pinpoints the importance of environmental changes on reproductive modes of species. As pointed above, the present study did not cover a seasonal sampling period when E. virens was represented by one female and no bisexual populations reported from Turkey so far. Among the variables used in CCA, habitat type (p=0.014; F=2.171) and water temperature (p=0.018; F=2.248) were the two most effective factors on species. Geiger (1998) using the data of European database on ostracods (NODE) underlined that there was no significant correlation between habitat type and reproductive mode of the species, although there were more parthenogenetic species in unstable (fluctuating) habitats and more sexual in stable environments. This was actually opposite to the common view (see Chaplin et al. 1994). We again did not find a clear correlation between habitat type and mode of reproduction, although it was one of the most effective factors on species. As argued by Geiger (1998), some other factors (e.g., producing resting eggs or stages) along with habitat type can be of important for colonization into the unstable habitats. Because we did not study the production of resting stages, we could not analyze its importance herein. However, what Geiger suggested was opposite to the common view that more sexual forms can be found in unstable conditions than stable where parthenogenetic forms can be high (e.g., Innes, 1997). However, as implied by Geiger (1998), we consider some other additional ecologically important factors such as water temperature that was the second effective one on species. Our results did not show a clear correlation of reproductive mode to any of those environmental variables used; however, they clarified that the correlation may not be explained with one or two factors at all. Hence, it implies that multiple factors seem to corresponding to reproductive mode of the species. On the other hand, according to Gülen (1985) there could be high possibilities of finding more bisexual populations in Anatolia because such populations are formed under slightly variable environmental conditions (opposite to the common view). When these conditions are changed to unstable, parthenogenetic reproduction is favored. Accordingly, bisexual populations were able to survive in refuges (Macedonian “Northwestern refuge” and Caucasian “Northeastern refuge”) during the glazier age that they could be able to disperse when conditions improved. Gülen did not provide supportive evidence of the relationship between modes of reproduction and habitat types, but his approach was at least partially supported by Geiger (1998) above. In contrast to these views, one may argue that parthenogenetic species could also be placed in the refugia as well. Nevertheless, at the moment, we cannot answer why there are so many bisexual populations in Turkey. On the other hand, considering that we had at least ten different types of habitats (Table 1) sampled during this study, a better explanatory factor on ostracod diversity and distribution seems to be suitability of habitats. Since increasing altitude did not cause a significant reduction in species number up to some level, our results did not show a clear support for the Rapoport Rule: in fact, species richness was not correlated with altitude.
Footnotes
Acknowledgments
We would like to thank Mr. Kerim Altinbağ for his tremendous effort and help on field sampling. Also, thanks must be given to Dr. Craig Stockwell and Sujan Henkanaththe (North Dakota State University) for their critical review on the first draft of the manuscript. In addition, two of our graduate students (Elif Başak and Samet Uçak) (Abant Ġzzet Baysal University) are thanked for their help in laboratory work.
Disclosure Statement
No competing financial interests exist.