
Abstract
Abstract
Singh, Rashim Pal, Kristin Franke, and Ben Wielockx, Hypoxia-mediated regulation of stem cell fate. High Alt Med Biol.13:162–168, 2012.—Hypoxia-mediated regulation of stem cell fate, or reduced oxygen availability, is a prominent feature during mammalian development and under physiological and pathological conditions in adults. Oxygen-sensing is therefore indispensable as it enables the cells to adapt instantaneously to an inappropriate pO2. This machinery relies primarily on hypoxia inducible factor (HIF). Moreover, a growing body of evidence proposes that different types of stem cells exist in a very hypoxic microenvironment, which may be beneficial for the maintenance of these cells and ensures continuous replenishment of dead or damaged cells in virtually all tissues of the body. Recent reports have shown that HIF is a critical player in these responses. However, a better understanding of the different HIF-related mechanisms is of utmost importance for the improvement of therapeutic strategies for tissue regeneration as well as hematological malignancies.
Introduction
Hypoxia maintains the undifferentiated state of stem cells, including hematopoietic (Takubo et al., 2010), mesenchymal (Méndez-Ferrer et al., 2010), and neural stem cell (Mazumdar et al., 2010), and influences quiescence, proliferation, and cell-fate commitment. Moreover, stem cells are essentially defined by their ability to self-renew and maintain a lifelong production of mature cells and therefore have been found to reside mostly in hypoxic niches (Eliasson and Jönsson, 2009; Mohyeldin et al., 2010). Low oxygen tensions maintain the multipotency of stem cells through the regulation of a fine balance between proliferation and differentiation.
In this review, we will focus on some significant findings that highlight the importance of hypoxia and the HIF pathway in different types of stem cells which adds an important new facet to our traditional view of hypoxia and stem cell fate.
Hematopoietic Stem Cells and Hypoxia
Of all the known types of stem cells, hematopoietic stem cells (HSCs) are probably the best characterized. Based on the expression of different surface markers, these cells are well defined and categorized into either the most primitive quiescent HSCs (long-term HSCs) or more mature and proliferating multipotent progenitors (MPPs). Both of these cell types are found in a specific population of bone marrow cells known as LSK cells (Lineage-Sca1+cKit+). Wilson and colleagues (2008) further divided the MPPs into four subgroups (MPP1-4) based on the absence or presence of CD135 and slam markers CD48 and CD150.
HSCs are responsible for the production of mature hematopoietic cells throughout the life of an organism; hence the ability to maintain their multipotency and self-renewal capacity is essential. Therefore, cell cycle activity in these cells is tightly regulated where oxygen levels in the local bone marrow (BM) environment play a crucial role in this regulation. Moreover, HSCs have been proposed to reside in regions of very low oxygen pressure compared to other tissues, which seems to be a requirement for their quiescence and reconstitution potential (Parmar et al., 2007; Kubota et al., 2008). Indeed, culturing of the most primitive HSCs under severe hypoxic conditions leads to a decrease in cell cycle activity and enhances their ability to reconstitute the stem cell compartment of lethally irradiated mice (Eliasson et al., 2010). The sedentary state and the positioning of HSCs in hypoxic zones are also reflected in their metabolic profile. For energy production, they mostly rely on the anaerobic process of glycolysis (Simsek et al., 2010).
Regulation of Hematopoietic Stem Cells through HIF
HIF1α plays a central role in hypoxia-mediated regulation of HSCs. Genetic deletion of ARNT (HIFβ), which abrogates the activity of HIF1α and HIF2α, results in defective embryonic hematopoiesis; a phenotype that was shown to be dependent on the interaction of HIF1α and ARNT. The multilineage hematopoietic progenitors could not proliferate in ARNT null embryos, leading to decreased numbers in the yolk sac (Adelman et al., 1999). In cultured adult HSCs, HIF1α mediates their hypoxia-induced quiescence and enhanced reconstitution ability. Hypoxia treatment of HSCs leads to their accumulation in the resting G0 phase, together with alterations in the cyclin-dependent kinase inhibitor p21, whereas p27 and p57 were not changed in LSK cells (Eliasson et al., 2010).
Recent reports have given new insights into the in vivo maintenance of the HSC pool and its dependency on HIF1α (Simsek et al., 2010; Takubo et al., 2010; Miharada et al., 2011). Conditional deletion of HIF1α in the HSCs leads to the loss of quiescence, accompanied by increased peripheral chimerism and subsequent exhaustion in serial BM transfers (Takubo et al., 2010). Enhanced proliferation is related to the senescence and final depletion of HSCs. Under conditions of severe stress, HIF1α-deficient HSCs display significant induction of p16Inka4/p19Arf, which is a hallmark for senescent stem cells (Naka et al., 2008). HIF1α null HSCs exposed to the stress of an ex vivo culture also show increased transcription of these cdk-inhibitors compared to their WT counterparts. Retroviral-induced expression of BMI-1, a transcriptional repressor of the Lnka4/Arf locus (Beã et al., 2001), in the HIF1α mutant HSCs rescued them from senescence and subsequent exhaustion in conditions of stress. In contrast, abolition of HIF1α in HSCs under steady state conditions did not alter hematopoiesis vastly, suggesting a pivotal role for HIF1α during conditions of severe stress (Takubo et al., 2010) (Fig. 1).
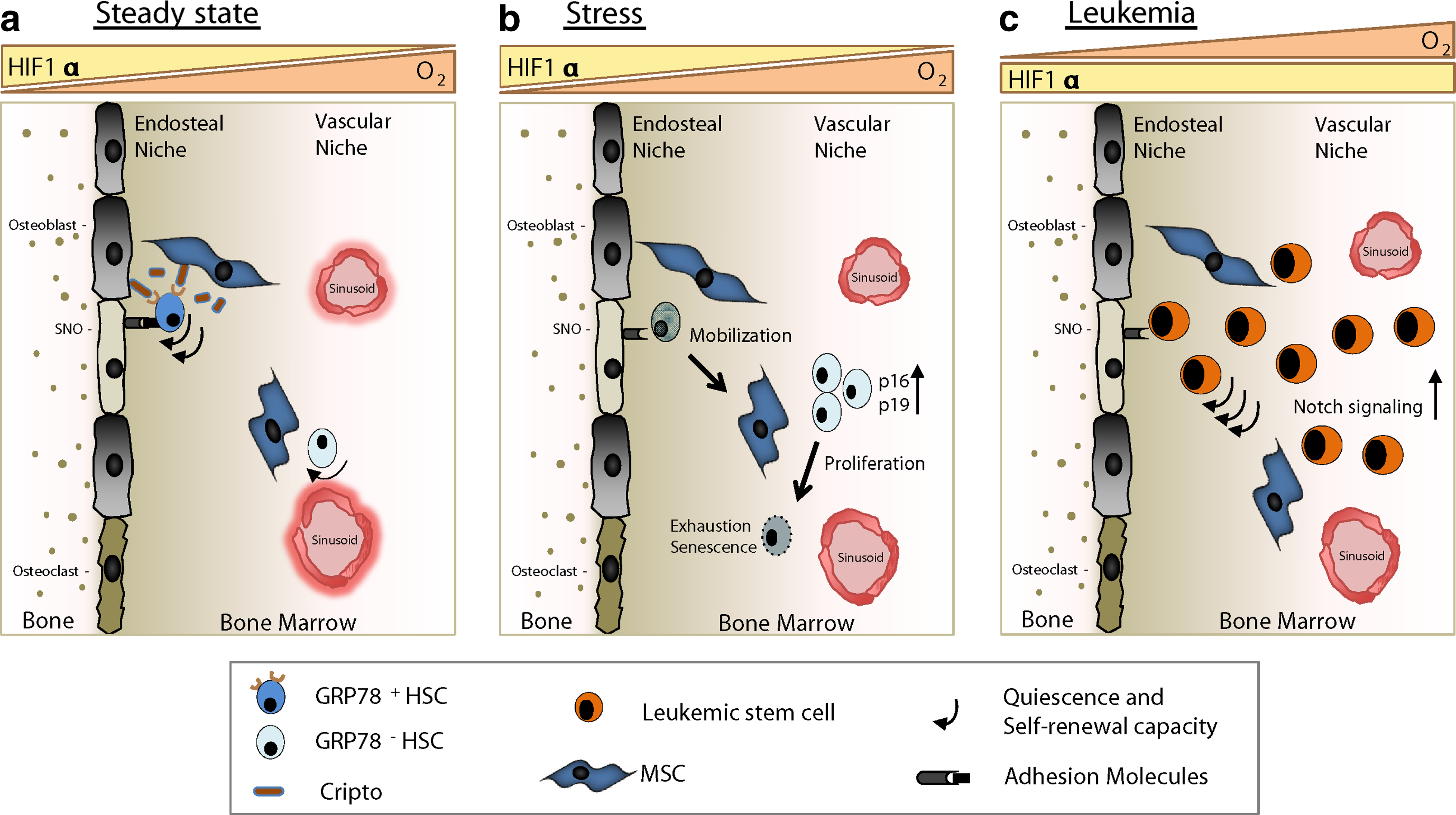
HIF1α-regulated steady state, stress, and malignant hematopoiesis.
It has also been shown that HIF1α should be stabilized specifically in HSCs to function normally. Overstabilization by removing VHL (pVHL) not only induced quiescence, reduced homing, and increased apoptosis in HSCs, but also in more mature hematopoietic progenitors (Takubo et al., 2010). As the proliferation of the latter populations is necessary for normal hematopoiesis, HIF1α should be selectively eliminated in them. The deletion of CITED2, another negative regulator of HIF1α, leads to the acute loss of HSCs via increased apoptosis and reduced quiescence (Kranc et al., 2009). HSCs lacking CITED2 have a poor reconstitution potential and their phenotype is very similar to pVHL-null HSCs. Transcriptional analysis of CITED2-null HSCs revealed that quiescence- and hypoxia-related genes like EGR1, p57, and Hes1 are affected (Du et al., 2012). Moreover, additional deletion of HIF1α in CITED2-deficient HSCs partially rescues the impaired HSC phenotype in a p57- and Hes1-dependent manner. Interestingly, deletion of CITED2 in mature lymphoid or myeloid cells did not alter their frequencies (Kranc et al., 2009). Taken together, this suggests that CITED2 is dispensable for mature hematopoietic cells, whereas the required deletion of HIF1α in late hematopoietic progenitors is executed by additional mechanisms. Maybe the HIF prolyl hydroxylases play a role in this regard. In that respect, it has been shown that short-term HSCs isolated from 10-week old mice have significantly higher transcripts of PHD2 compared to long-term HSCs (Takubo et al., 2010). Although rather unexplored, PHD-mediated regulation of HIF1α would certainly give more insights into the maintenance and functionality of HSCs and MPPs.
Since HSCs rarely divide and exist in a very hypoxic niche, they require very stringent metabolic adaptations. In this respect, it seems that HIF1α is a key player in mediating the switch from oxidative to glycolytic metabolism. Recently, it has been shown that a distinct population of cells in the BM characterized by low mitochondrial potential is highly enriched in long-term HSCs. These cells have low amounts of ATP and utilize the anaerobic glycolysis for energy production. Moreover, they have highly increased levels of HIF1α that is transcriptionally regulated by a stem cell associated transcription factor Meis1 (Simsek et al., 2010). Although it has been shown that HSCs have increased glycolytic activity accompanied by enhanced levels of HIF1α, it is still largely unknown how HIF1α promotes the glycolytic mode of metabolism.
Miharada and colleagues (2011) identified Cripto as a niche-related factor and reported that a subpopulation of HSCs express its receptor GRP78. The GRP78+ HSCs exhibit low cell cycle activity, slow BM reconstitution after transplantation, and low mitochondrial activity compared to GRP78- HSCs. GRP78+ HSCs were located in the endosteal region of bone and were found to be largely positive for pimonidazole, a hypoxic marker (Fig. 1). Additionally, they demonstrated that the interaction of Cripto and GRP78 leads to the activation of the PI3K/Akt pathway, whilst proteomics analysis revealed an increase and/or phosphorylation activation of the glycolysis enzymes, thus suggesting a role for Cripto in the metabolic regulation of HSCs. Moreover, the Cripto promoter region is known to have HREs to which the stabilized HIF1 complex binds (Bianco et al., 2009). The analysis of HIF1α conditional knock-out mice showed that mutant LSK cells had lower transcripts of Cripto. These findings suggest that, although HIF1α partially controls the expression of Cripto in HSCs, Cripto can also be regulated by other pathways (e.g., Wnt or Smad) (Bianco et al., 2009). Furthermore, HIF1α-deficient mice have decreased numbers of GRP78+ HSCs in the endosteal niche. Thus, Cripto acts as a docking molecule for HSCs keeping them in the hypoxic niche, which is directly regulated by HIF1α. Blocking antibodies against GRP78 led to the decrease of GRP78+ HSCs in the endosteal region reconfirming this notion (Fig. 1). Additionally, the PI3K/Akt pathway is involved in the protection of HIF1α from pVHL-independent degradation (Zhou et al., 2004) while Cripto-GRP78 signaling activates this pathway. Henceforth, it can be inferred that the Cripto-GRP78 pathway has a significant role in HSC maintenance during hypoxia through regulation of the HIF1α complex leading to glycolytic metabolism.
Hematopoietic Cancer Stem Cells and Hypoxia Inducible Factor
Since HIF1α plays a pivotal role in HSC biology, it's intriguing to know whether HIF signaling would be involved when hematopoiesis gets deregulated as in hematological malignancies. What has been shown earlier is that normal HSCs share a common phenotype and functionality with cancer stem cells (CSCs) (Wang and Dick, 2005; Essers and Trumpp, 2010). These CSCs are usually slow cycling and have the property of immortality. CSCs are not killed by conventional drug regimens as they target only highly proliferating cells, leading to drug resistance and tumor recurrence. The uncontrolled proliferation of tumor cells creates hypoxic zones in tumors, hence making hypoxia-related molecules likely targets for novel cancer therapies (Keith and Simon, 2007). Recently, Wang and colleagues (2011a) described the role of hypoxia and self-renewal capacity of CSCs, using a mouse lymphoma and a xenogeneic human acute myeloid leukemia (AML) model. Not only were they able to identify a CSC population in a mouse lymphoma positive for the cKit and Sca1 surface markers, but in addition their multipotency was confirmed by the immortality in long-term cultures and ability to initiate lymphomas after serial transplantations (Wang et al., 2011a). Moreover, these CSCs expressed high levels of HIF1α both at the mRNA and protein level along with HIF1α-target genes Glut1 and cMyc. VHL transcripts were virtually undetectable in CSCs, thus making physiological hypoxia irrelevant for preserving their ability to self-renew. Additionally, treatment with low doses of echinomycin, a HIF1α inhibitor, could selectively kill cKit+Sca1+ CSCs of mouse lymphoma. Abrogation of HIF1α via shRNA in lentiviruses led to the decreased self-renewal and tumor initiating ability of CSCs. Furthermore, the Notch pathway seems to be a critical target of HIF1α in this CSC context as it was found to work downstream of HIF (Gustafsson et al., 2005). In the cKit+Sca1+ CSCs, it was shown that HIF1α potentiates Notch signaling by blocking the negative feedback regulation of Hes1 (Wang et al., 2011a) (Fig. 1). Moreover, the precise extent to which HIF1α should be stabilized will define the physiological outcome and therefore should be kept in mind while designing future therapies.
Mesenchymal Stem Cells and Oxygen Tension
Mesenchymal stem or stromal cells (MSCs) are multipotent cells with an ability to differentiate into osteogenic, chondrogenic, myogenic, and adipogenic lineages (Pittenger et al., 1999; Kuhn and Tuan, 2010). Originally these cells were discovered in bone marrow and characterized as fibroblast stromal cells, which formed colonies that were adherent to plastic plates with an enormous osteogenic potential (Friedenstein et al., 1968; Friedenstein et al., 1987; Friedenstein et al., 1974). The term mesenchymal stem cell was first used by Caplan (1991). Later it was shown that these cells have the potential to differentiate into different cell lineages (Pittenger, 1999), thus making them an important tool for cell-based therapies. Until now, MSCs have been successfully isolated from bone marrow, adipose tissue, placenta, and umbilical cord (Chung et al., 2009; Lund et al., 2009; Kuhn and Tuan, 2010). These cells are essentially negative for CD45, the common antigen for leukocytes, but can be identified by their expression of surface markers like CD44, CD73, CD90, and CD105 (Dominici et al., 2006). Another feature of MSCs is their ability to migrate and translocate to sites of inflammation, creating a special cytokine milieu and thus altering the immune responses (Blank et al., 2008; Slavin et al., 2008; Chung et al., 2009; Hata et al., 2010). Moreover, Crisan and colleagues (2008) linked the adult human MSCs to perivascular cells or pericytes. Although described in the latter report next to the blood vessels, the regions where MSCs are found in the bone have very low oxygen concentrations (Harrison et al., 2002). In that respect, a Nestin+ MSC population has been recently described to express HSC maintenance genes like cKit-ligand, Ang-1 and Cxcl12 and spatially associate with HSCs and adrenergic nerve fibres (Méndez-Ferrer et al., 2010). Moreover, these cells are known to be predominantly located in very hypoxic niches, at the lowest end of an oxygen gradient (Parmar et al., 2007). Thus it can be inferred that hypoxia may very well play a significant role in keeping MSCs in an undifferentiated state and thereby prevent their exhaustion. Actually, it has been observed that cultivating human MSCs under hypoxic conditions prevents their differentiation and increases their proliferation potential at the same time (Fehrer et al., 2007). Moreover, this also induced the expression levels of HIF2α and Oct4, a protein associated with an undifferentiated phenotype and self-renewal of stem cells (Grayson et al., 2007).
HIF1α and two of its target genes, vascular endothelial growth factor (VEGF) and stromal cell derived factor 1α (SDF1α), have been shown to greatly influence the functionality of MSCs (Yun et al., 2009; Busletta et al., 2011; Liu et al., 2011; Zou et al., 2011). Treatment of human MSCs with a steroid hormone 17β-estradiol (E2) leads to the enhanced cell cycle activity along with activation of PKC and PI3K/AKT signaling pathways. The increased signaling of PKC and PI3K/AKT in turn leads to the upregulation of HIF1α, which was demonstrated via the use of chemical inhibitors for both pathways (Yun et al., 2009). Furthermore, E2-treated MSCs also exhibited HIF1α-dependent increased of VEGF levels, which mediated the proliferation of MSCs (Yun et al., 2009). HIF1α-mediated secretion of VEGF is also known to enhance the migration capacity of MSCs (Busletta et al., 2011), stimulate an increase in bone marrow sinusoidal vessels and thereby possibly facilitating the egress of MSCs (Liu et al., 2011). SDF-1α protein levels are increased in the peripheral blood of rats during MSC mobilization, whereas it promotes their migration under hypoxic conditions in vitro (Liu et al., 2011). Consequently, this accentuates a role for hypoxia and HIF1α in MSC mobilization that could be exploited for novel regenerative therapies. HIF1α in MSCs has also been implicated in coupling of angiogenesis with osteogenesis (Riddle et al., 2009). MSCs with constitutively stable HIF1α not only show increased VEGF levels, but also more ossification or new bone formation, most probably via the induction of BMP2 (Zou et al., 2011).
Neuronal Stem Cells Regulated by Hypoxia
It has been a long-held dogma that, unlike the hematopoietic system or the epithelial skin layer, neural cells cannot be regenerated. Therefore, the discovery of neural stem cells (NSCs) in the developing and adult brain was earth-shattering (reviewed in Gage, 2000). NSCs possess life-long self-renewal abilities and can differentiate into neurons, as well as in glial cells such as astrocytes and oligodendrocytes.
Adult neurogenesis takes place in the subgranular zone within the dental gyrus of the hippocampus, in the subventricular zone, and in the olfactory epithelium; areas where NSCs are also found. Several NSC subtypes have been described depending on the brain region and specific surface marker (Wang et al., 2011b). NSCs can be propagated in tissue culture where they form free-floating cell aggregates or neurospheres, which bear self-renewal potential but also the ability to differentiate into mature neural cells.
Compared to the rest of the mammalian body, the brain is a relatively hypoxic organ. Oxygen concentrations range from 8% in the meninges (pia mater) to 0.55% in the midbrain, which requires great adaptation potential of the different cell compartments (Mannello et al., 2011). Similar to other stem cell types, the ambient oxygen concentrations have been shown to regulate the proliferation and differentiation potential of NSCs. Experiments with immortalized human NSCs (ihNSC) revealed that mild hypoxia (2.5% oxygen) is optimal for NSC survival, proliferation and maintenance of multipotency (Santilli et al., 2010). Interestingly, culturing NSCs and neural progenitors in mild hypoxia conditions activates Oct3/4, the Wnt/beta-catenin, and Notch signaling pathways, which are involved in self-renewal and proliferation (Pistollato et al., 2010; Chavali et al., 2011; Cui et al., 2011). In contrast to mild hypoxic conditions, severe hypoxia (1% of oxygen) and nonphysiologically high oxygen (20%) decreased neurogenic abilities and increased the apoptotic rate. Interestingly and similar to other stem cells, the few cells surviving severe hypoxia remained in a state of quiescence (Santilli et al., 2010). Not just the survival but also the commitment of NSCs towards one of the three brain cell types (neurons, astrocytes, oligodendrocytes) highly depends on the oxygen concentration used in culture. Whereas 1% and 20% oxygen favor astrocyte differentiation, mild hypoxia allows for the augmented formation of neuronal cells and oligodendrocytes (Pistollato et al., 2007; Santilli et al., 2010). Furthermore, mild hypoxia has been shown to increase the expansion of rat and human neural precursors and the generation of tyrosine hydroxylase (TH) positive dopaminergic neurons (Studer et al., 2000; Storch et al., 2001). The transcription factor HIF1α is one of the main players inducing the cellular hypoxic response. Deletion of HIF1α in neural precursor cells (nestin+) disturbs the normal brain development and leads to an abnormal increase in cortical apoptosis (Tomita et al., 2003). Accordingly, these cells also showed impaired differentiation of TH positive neurons (Milosevic et al., 2007).
Since the adult CNS shows only limited regeneration potential, the hope to repair damaged brain tissue with NSCs, which produce fresh neural cells, is tempting. NSC cultures are subject to intensive investigation and one of the crucial factors that determine proliferation, differentiation, and survival of the NSCs is the environmental oxygen concentration.
Conclusions and Future Directions
Since the discovery of HIF1α two decades ago, intense research has led to the identification of several molecular targets associated with hypoxia during various physiological and pathological processes. Recent studies have also suggested that many different stem cells, including hematopoietic (Takubo et al., 2010), mesenchymal (Méndez-Ferrer et al., 2010), and neuronal stem cells (Mazumdar et al., 2010), reside in specialized niches defined by low oxygen tension. Moreover, HIF1α has been shown to be essential for the preservation of quiescence, self-renewal, and proliferation of these cells, as well as their reprogramming to glycolytic metabolism. For HSCs, these functions have recently been extended to hematological cancer stem cells (Wang et al., 2011a). Although it is increasingly clear that HSCs, MSCs, NSCs and other progenitors interact with each other within the different niches (Kuhn and Tuan, 2010; Méndez-Ferrer et al., 2010), many essential questions remain to be clarified. In that respect it is necessary to elucidate what levels of HIF1α are necessary to maintain quiescence in HSCs versus hematological CSCs. Is there a role for HIF2α in stem cells and is that role essentially different from that of HIF1α in maintaining ‘stemness’? Additionally, to what extend can the regulation of PHDs or other HIF regulators be beneficial in the development of new therapeutic strategies for tissue regeneration, as well as hematological malignancies.
Footnotes
Acknowledgments
We thank Soulafa Mamlouk for helpful discussions. We apologize to any colleagues whose work was not cited because of space limitations.
Author Disclosure Statement
This work was supported by the Emmy Noether program (the Deutsche Forschungsgemeinschaft - DFG, Germany) and the MeDDrive program (TU Dresden, Germany).
The authors declare no competing financial interests.