
Abstract
Abstract
Jain, Kanika, Geetha Suryakumar, Rajendra Prasad, and Lilly Ganju. Upregulation of cytoprotective defense mechanisms and hypoxia-responsive proteins imparts tolerance to acute hypobaric hypoxia. High Alt Med Biol 14:65–77, 2013.—Exposure to high altitude is a well-known environmental stress with physiological and metabolic consequences, with the major stressor being hypobaric hypoxia. The disruption in cellular homeostasis elicits several acute and chronic adaptations designed to diminish the stress imposed by the hypoxic insult. Highly conserved cellular machinery protects the myocardium from damage under reduced oxygen tension. In the present study, adult Sprague-Dawley rats were exposed to an altitude of 9754 m in a decompression chamber and screened on the basis of the time taken for onset of gasping. The animals were grouped as susceptible (<10 min), normal (10–25 min), and tolerant (>25 min). Histologically, susceptible animals showed increased myocardial inflammation and infiltration and greater CK-MB activity. These animals showed a three-fold increase in reactive oxygen species levels and subsequent oxidative damage to proteins and lipids as compared to control unexposed group. In tolerant animals, the damage was minimal. The resistance to damage in these animals was possibly due to enhanced myocardial antioxidant enzymes, catalase and superoxide dismutase. A significantly higher expression of HIF-1α and its responsive genes, including EPO, HO-1, and GLUT1, was seen in tolerant animals, although VEGF expression was enhanced in the susceptible group. Cytoprotective chaperones, HSP70 and HSP90, were elevated in the tolerant animals. The differential expression of these hypoxia-responsive molecules may thus act as potential biochemical markers for screening and identifying individuals susceptible to environmental stress.
Introduction
Mechanistically, low oxygen availability at high altitude decreases mitochondrial respiration, leading to the accumulation of reducing equivalents in the cellular electron transport chain, causing partial reduction of oxygen, which in turn generates superoxide radicals (Mohanraj et al., 1998). While reactive oxygen species (ROS) have been known to play a cytoprotective role in assisting in high altitude acclimatization (Boutilier, 2001), excessive ROS generation has a detrimental cellular effect (Yu, 1994). A considerable amount of literature is available highlighting an increased production of indicators of oxidative stress in breath, blood, urine, and tissue of laboratory rodents in response to hypoxia (Radak et al., 1997; Ilavazhagan et al., 2001).
The ability to respond to any kind of environmental stress differs from individual to individual. Along with the genetic adaptations likely playing a role in shaping the morphological and physiological differences (Moore, 2001), each individual has an inherent threshold of stress with which it can deal. Although every organism undergoes adaptation to high altitude environments, it has been well established that high altitude natives are better adapted to such conditions than lowlanders. Numerous studies have shown that the indigenous high altitude population have evolved mechanisms to more closely relate oxygen supply and metabolism and subsequent energy generation (Hoppeler and Vogt, 2001). Studies by Gelfi et al. (2004) have revealed that lowlanders show a much greater extent of oxidative muscular damage as compared to high altitude natives, indicating that high- and low-altitude populations adopt different metabolic strategies to ameliorate the consequences of oxidative stress. Interestingly, antioxidant enzymes have been found to be lower in the highlanders, indicating their adaptation during their long dwelling period at the high altitude; however sojourners to high altitude display an increase in activity of antioxidant defense systems (Sinha et al., 2009). It can be hypothesized that these same antioxidant defensive mechanisms may play a role in imparting tolerance to some lowlanders, thus increasing their ability to endure the hypobaric hypoxia conditions.
It has been proposed that an altered regulation of oxygen delivery under hypoxic conditions contributes to the successful high-altitude adaptation in Tibetan and Andean populations (Beall, 2007; Peng et al., 2010). Studies have also been reported highlighting an enhanced altered hypoxia sensing, rather than hypoxia response mechanism, as the reason for better adaptation in natives (Scheinfeldt et al., 2012). Mammalian cells detect the decrease in oxygen concentrations to activate a variety of responses allowing better adaptation. One such response is stabilization of the protein hypoxia-inducible factor (HIF-1 and −2α), which regulates the expression of genes mediating adaptive responses (Hochachka and Rupert, 2003). This includes a prominent role for more than 100 hypoxia-sensitive genes, such as inducible nitric oxide synthase (iNOS), glucose transporter (GLUT-1), erythropoietin (EPO), heme oxygenase-1 (HO-1), and others, regulated by the activation of these nuclear factors. Studies on populations of high altitude natives such as Sherpas and Tibetans have shown evidence of genetic variation in the hypoxia-inducible factor (HIF)-1α gene (Suzuki et al., 2003). Study of different strains of rodents, having a varying degree of tolerance to high altitude, has indeed attributed a significant role of altered HIF-1α expression in hypoxic tolerance (Engebretsen et al., 2007). Stimulation of these proteins through hypoxic preconditioning has been established to play a part in protecting from tissue damage (Sinha et al., 2010; Lin et al., 2011; Shukla et al., 2011).
High altitude acclimatization and its benefits for the lowlanders have long been considered and studied, but the identification of biochemical changes and protein markers within the myocardium remains incomplete. While tolerance to such environmental stressors differs from individual to individual, the recognition of such markers may provide a clue into the response of a person. The present study aims to evaluate the biochemical responses of the myocardium responsible for the resistance or susceptibility of animals subjected to acute hypobaric hypoxic stress.
Materials and Methods
Animals and experimental design
Animals
Male Sprague-Dawley rats (150±15 g) were used for all experiments. Animals were maintained under a 12-h light–dark cycle at temperature 24°C±2°C in the Institute's animal house facility. The study was approved by the Animal Ethical Committee of the institute in accordance with Committee for the Purpose of Control and Supervision on Experiments on Animals (CPCSEA) of the government of India.
Hypobaric hypoxia exposure
Hypoxic tolerance was determined by measuring the time taken for the onset of gasping (GT). Adult male Sprague-Dawley rats were exposed, one at a time, to simulated hypobaric hypoxia of 9250 m in an animal decompression chamber (Decibel Instruments) coupled to a mercury barometer at 32°C (Bhatia et al., 1966; Kumar et al., 1999; Shrivastava et al., 2008) The airflow in the chamber was 2 L/min, while the relative humidity was maintained at 40%–50%. The time taken for appearance of the first sign of gasping was recorded using an electronic stopwatch. Onset of gasping was identified as a hyperventilatory response, accompanied by a visible difficulty in breathing and enhanced breathing rate, beyond which time, if exposed, there occurred a cessation of breathing in the animal. The animals were given three consecutive exposures to identify their tolerance to hypoxic stress. Accordingly, the hypoxia exposed rats were divided into 3 groups (n=8): normal (10–25 min, N), tolerant (>25 min, T), and susceptible (<10 min, S). Unexposed control group rats (n=8) were maintained in the normoxic condition within the same laboratory conditions.
Histopathological studies
At the end of hypoxic exposure, the animals were sacrificed; hearts were excised and immediately fixed in 10% buffered neutral formalin solution. The fixed tissues were embedded in paraffin and serial sections (4 μm thick) were cut using microtome (Leica RM 2125, Germany). Each section was stained with hematoxylin and eosin (H and E). The sections were examined under the light microscope (Nikon, Tokyo, Japan) and photomicrographs were taken.
Oxidative stress and damage
After the hypoxic exposure, the animals were anesthetized under sodium pentobarbital and tissue collected. The hearts were perfused in saline and used fresh or snap frozen in liquid nitrogen and stored at −80°C for further use
ROS measurement
2′,7′-Dichlorofluorescein (DCFH) was used as a fluorescent probe to measure the rate of oxidant production in the fresh tissue homogenates according to LeBel et al., (1992) with some modifications (Cathcart et al., 1983, LeBel et al., 1992). DCFH-DA, a nonfluorescent lipophilic dye, passively diffuses through cellular membranes where it is cleaved into 2,7-dichlorofluorescein (DCF) by the action of intracellular esterases, and thus fluoresces in the presence of ROS. The production of free radicals was determined as described earlier. Briefly, 150 μL of heart homogenate was incubated with 10 μL of 100 μM DCFH-DA for 30 min in dark. Fluorescence was read using a fluorimeter (Perkin Elmer, UK) with excitation at 485 nm and emission at 535 nm. Readings were obtained in arbitrary flourometric units and results expressed as fold change in free radical generation.
Lipid peroxidation
Lipid peroxides are unstable indicators of oxidative stress in cells that decompose to form more complex and reactive compounds such as malondialdehyde (MDA). Lipid peroxidation was measured by direct estimation of MDA using the modified method of Dousset et al., (1983). Briefly, to 250 μL of myocardial tissue supernatant, 750 μL of trichloroacetic acid (20%) and 750 μL of thiobarbituric acid (0.67% in 0.05 M NaOH) was added. The samples were incubated in a water bath at 85°C for 45 min. The samples were then kept at room temperature followed by centrifugation at 400 g for 5 min. About 200 μL of the supernatant was removed and the absorbance was measured at 531 nm using UV-Vis spectrophotometer (BioRad, USA). Results were expressed as nanomoles of MDA formed per mg protein.
Marker for cardiac injury: Quantification of creatine kinase-MB
Creatine kinase MB is a known marker for assessing cardiac integrity. Its levels were quantified in the perfused rat heart tissue homogenate, using a commercially available kit, rat CK-MB ELISA (Cusabio, China) as per manufacturer's instructions. The concentrations were expressed as ng/mL.
Protein oxidation and damage
Advanced oxidation protein products
Advanced oxidation protein products (AOPPs), considered as markers to evaluate oxidant induced protein damage, are formed by the reaction of proteins with chlorinated oxidants such as chloramines or hypochlorous acid. The spectrophotometric detection of AOPP levels, first described in plasma by Witko-Sarsat et al. (1996), was modified for heart tissue. Briefly, heart tissue homogenates, prepared in 0.154 M KCl-EDTA, were diluted 1:5 with phosphate-buffered saline (PBS), pH 7.4. For standard curve, 200 μL of chloramin T (0–100 μmol/L) and 200 μL of PBS as blank were applied on a microtiter plate. Similarly 200 μL of diluted samples were applied. 10 μL of 1.16 M potassium iodide and 20 μL of acetic acid were added to each well and absorbance at 340 nm measured immediately. Concentration of AOPPs were obtained in chloramine units and expressed as μmol chloramine/mg protein.
Protein carbonyl content
Oxidative modifications of amino acid residues include derivatization of amino acid residues such as proline, arginine, and lysine to reactive carbonyl derivatives. Briefly, 2,4- dinitrophenylhydrazine reacts with protein carbonyl forming a Schiff base to produce the corresponding hydrazone, which can be analyzed spectrophotometrically (Levine et al., 1990). Heart tissue was homogenized in ice cold 50 mM phosphate buffer (pH 7.2) containing 1 mM EDTA and proteinase inhibitor cocktail. The homogenate was centrifuged at 10,000 g for 15 min, and the supernatant was checked for presence of nucleic acids by measuring the absorbance at 260 and 280 nm. A ratio of 280/260 nm more than 1 indicated that the sample was free of nucleic acid contamination. To 200 μL of sample, 600 μL 10 mM 2,4-dinitrophenylhydrazine (DNPH) in 2 N HCl was added; 600 μL of 2 N HCl was added as a blank control. The mixture was incubated for 1 h at room temperature. The protein was precipitated with an equal volume of 20% trichloroacetic acid and was washed three times with ethanol/ethyl acetate (1:1 v/v). The final precipitate was dissolved in 400 μL of 6 M guanidine hydrochloride (pH 2.3), and the insoluble debris was removed by centrifugation. The absorbance of the DNPH derivatives was measured at 360 nm. The concentration of carbonyl groups was calculated by using an absorbance coefficient 22 nM/cm and expressed as nanomole carbonyl per mg of protein.
Antioxidant enzyme activity
Superoxide dismutase activity
Superoxide dismutase (SOD; E.C. 1.15.1.1) activity was estimated in heart homogenates by the method of Marklund and Marklund (1974). The total SOD assay volume (3.0 mL) consisted of 1.5 mL of 50 mM Tris-cacodylate buffer pH 8.2, 0.3 mL of 1 mM nitro blue tetrazolium salt (NBT), 0.3 mL of Triton-X-100 (0.01%), 0.8 mL of water, 0.1 mL of sample, and 0.01 mL of 60 mM pyrogallol. A blank was run simultaneously consisting of 0.1 mL water instead of 0.1 mL sample. Enzyme kinetic activity was recorded at 540 nm for 3 min and change in OD per minute (Δ OD) was used to calculate % auto-oxidation inhibition to derive SOD units (U). One U of SOD was defined as 50% inhibition of the auto-oxidation caused by a certain value of enzyme. The results of SOD activity have been expressed as U (mg protein)−1.
Catalase activity
Catalase (CAT; E.C. 1.11.1.6) activity was measured in heart tissue homogenates by the method of Aebi (1984). The reaction mixture consisted of 2.9 mL buffer substrate (containing 0.1% H2O2 in 50 mM sodium potassium phosphate buffer, pH 7.0) and 0.1 mL sample in the final 3.0 mL assay volume. Change in absorbance was recorded for 150 s (every 15 s) at 240 nm. Catalase activity was calculated using an extinction coefficient of 0.041 cm2 (mmol)−1 and expressed as mmol H2O2 consumed per minute per mg of protein (mmol min−1 (mg protein)−1).
Reduced glutathione
Reduced glutathione (GSH) was measured in heart tissue homogenates by the method of Beutler et al. (1963). To 0.1 mL of sample was added 0.9 mL distilled water and 1.5 mL of precipitating reagent (3.34 g metaphosphoric acid, 0.4 g EDTA and 60.0 g NaCl). The mixture was allowed to stand for 5 min at room temperature and centrifuged for 15 min at 4000 rpm at 4°C. Supernatant (1.0 mL) was taken and to it 4.0 mL of 0.3 M phosphate solution and 0.5 mL 5-5′-dithiobis- (2-nitrobenzoic acid) (80 mg in 1% sodium citrate) were added. The developed yellow complex was read immediately at 412 nm on a spectrophotometer (Bio-Rad spectrophotometer, Smart Spec-3000). Reduced glutathione (Sigma) was used as a standard. Sample GSH concentrations were calculated and expressed as mmol of GSH per mg protein.
Vascular tone modulators
Nitrite levels
Nitrite, a biological metabolite of nitric oxide (NO), was measured in heart tissue homogenate using Griess reagent (Barrias et al., 2002). Briefly, 100 mg heart tissue was homogenized in 1 mL PBS and centrifuged at 2500 g for 10 min. Supernatant (100 μL) was mixed with an equal volume of Griess reagent (Sigma, USA) and incubated for 30 min in darkness. A chromophore azo-derivative thus formed was measured at 540 nm microplate reader. A standard curve was prepared by using different dilutions of sodium nitrite with each assay. The results were expressed in nmol/mL sodium nitrite.
Endothelin-1 content
Expression of endothelin-1, a potent vasoconstrictor released from the endothelium in hypoxic stress, was quantified in the perfused rat heart tissue homogenate, using a commercially available kit, rat endothelin-1 ELISA (Cusabio, China) as per manufacturer's instructions. The tissue homogenates were diluted 1: 20 using the sample diluent provided in the kit. The concentrations were expressed as pg/mL.
Hypoxia-responsive proteins
EPO content
EPO was quantified in the perfused rat heart tissue homogenate using commercially available kits, rat/Mouse Erythropoietin Immunoassay (R&D Systems) as per manufacturer's instructions. The concentrations were expressed as pg/mL.
Heme oxygenase-1 content
Heme oxygenase levels were determined in the perfused rat heart tissue homogenate using a commercially available kit, rat HO-1 ELISA (Enzo Life Sciences, USA) as per manufacturer's instructions. The concentrations were expressed as ng/mL.
VEGF content
VEGF content was measured by Rat VEGF ELISA (R&D Systems, Minneapolis, MN) according to the manufacturer's instructions. The intensity of the color reaction was read spectrophotometrically in a plate reader and the concentration expressed in pg/mL.
Immunoblotting
Preparation of nuclear and cytosolic extract
For nuclear and cytosolic fractionation, frozen heart tissue was homogenized in ice-cold buffer A containing 0.5 M sucrose, 10 mM HEPES, 10 mM KCl, 1.5 mM MgCl2, 10% glycerol, 1 mM EDTA, 1 mM dithiothreitol (DTT), 1 mM phenyl methyl sulfonyl fluoride (PMSF) fortified with protease inhibitors (Sigma, USA). Homogenates were kept on ice for 15 min and 0.6% Nonidet P-40 was added, and then centrifuged for 10 min at 2000 g at 4°C. The supernatant with cytosolic fraction was collected, stored, and the pellet was dissolved in cold buffer B (20 mM HEPES, 1.5 mM MgCl2, 0.3 mM NaCl, 0.2 mM EDTA, 20% glycerol, 0.5 mM DTT, 0.5 mM PMSF, and a cocktail of protease inhibitors) for the nuclear fraction. It was incubated for 30 min on ice followed by centrifugation at 20,000 g at 4°C for 15 min. The supernatant containing the nuclear fraction was aliquoted and stored at −80°C for further analysis. Total protein concentrations were determined using the Bradford protein assay (Bradford, 1976).
Western blot analysis
Protein expression of HSP70, HSP60, and HSP90, GLUT1, HO-1, and VEGF were quantified in cytosolic extract, whereas HIF-1α was analyzed in the nuclear extract by Western blot analysis. Primary antibodies against HO-1, GLUT1, and VEGF were obtained from Santa Cruz and those against HSP70, HSP60, HSP90, and HIF-1α were purchased from Sigma. Protein (50 μg) was separated by 10% SDS-polyacrylamide gel electrophoresis and transferred onto a nitrocellulose membrane (Hybond; Millipore, Billerica, MA). The membranes were blocked with 3% bovine serum albumin in PBS containing 0.1% Tween 20 (Sigma, USA), washed and probed with respective mouse/rabbit polyclonal antibodies (Sigma, USA and Santa Cruz). The membranes were then incubated with anti-mouse/rabbit-IgG HRP conjugate (Sigma). The membrane was washed and incubated with chemiluminescent substrate (Sigma, USA) and the bands were developed using X-ray films (Kodak, Rochester, NY). Quantification was performed by densitometry using ImageJ software.
Statistical analysis
All the experiments were performed on five different occasions, and data are presented as mean±SEM. One-way analysis of variance with post-hoc Bonferroni analysis was used to determine statistical significance between groups. All analyses were performed using SPSS 17 software. A p value of ≤0.05 was considered significant. Densitometric data for blots are expressed as a percentage of the control mean density after normalization to loading controls.
Results
Screening for differential tolerance
Taking the onset of gasping as an indicator for tolerance to acute sublethal hypoxic stress, a total of 115 animals were screened, and categorized into the following groups, with the value mentioned indicating the percentage of animals in each group:
• Normal range (10–25 min): 55% • Tolerant (>25 min): 27% • Susceptible (<10 min): 18%
Histopathological analysis and tissue damage
Figure 1a (i) shows a light micrograph of control rat heart showing normal architecture. In normal and tolerant animals [Fig. 1a (ii) and 1a (iii)], there was evidence of mild myocardial edema, with a presence of inflammatory cells in the myocardial sections from the normal group. Right ventricle sections from animals susceptible to sublethal hypoxic stress showed myocardial membrane damage and infiltration of inflammatory cells as compared to those in control unexposed group [Fig. 1a(iv)].
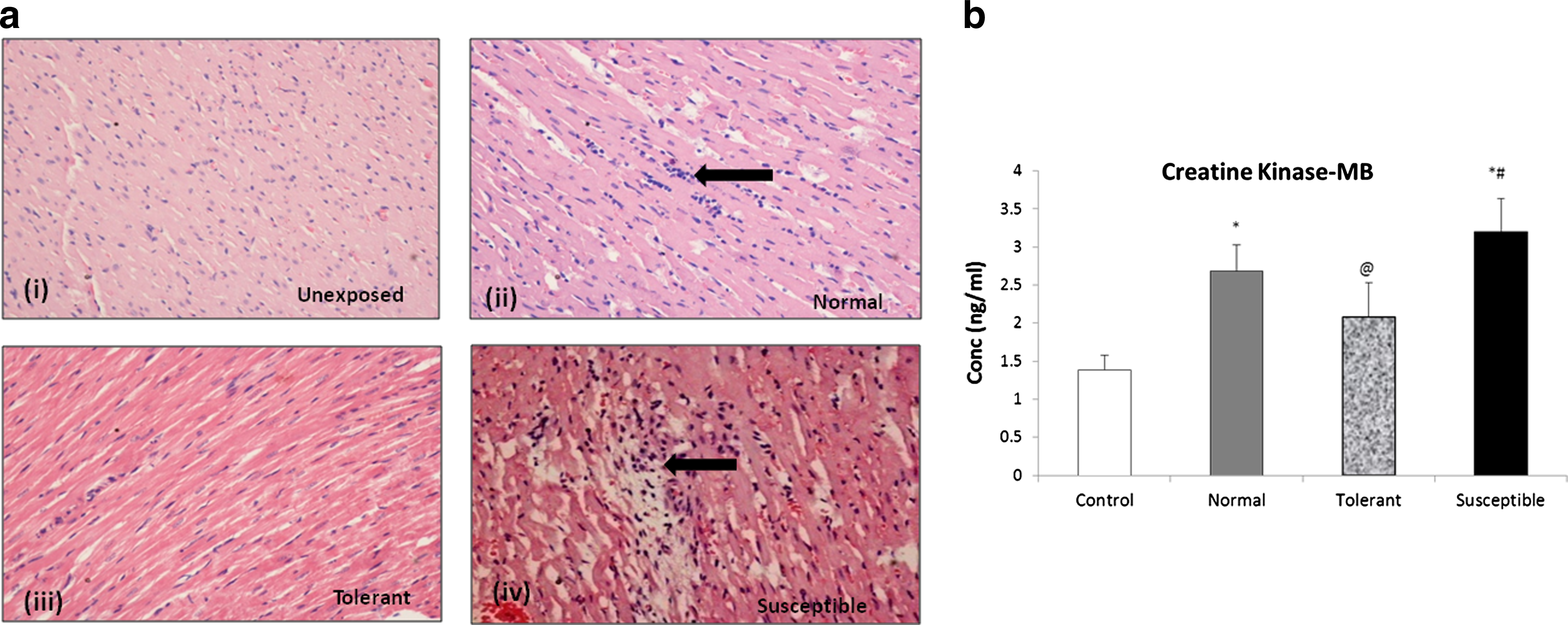
The integrity of the myocardial tissue under hypoxic stress was identified by the expression of CK-MB. This enzyme has been known to have an enhanced activity under oxidative stress and acts as marker for assessing cardiac damage. In our study, the expression of this protein was found to be significantly enhanced, more than 2-fold in the susceptible group, 3.20±0.43 ng/mL, p<0.001, with reference to the control unexposed, 1.38±0.19 ng/mL (Fig. 1b). The animals tolerant to hypoxic stress, showed a CK-MB level of 2.07±0.44 ng/mL, p<0.01, when compared to susceptible animals. Normal range animals also showed an elevated expression of CK-MB (2.69±0.34 ng/mL).
Hypobaric hypoxia-induced oxidative stress and damage
Reduced oxygen availability has long been known to cause an increase in generation of free radicals within the cell. In our study, we observed a 2.5-fold increase in the ROS generation within the myocardium of more susceptible animals, as compared to the control group (Fig. 2a). The increase in free radical generation in tolerant animals was less significant as compared to control. The damage to lipid and protein structure, as measured by lipid peroxidation and protein carbonyl derivative formation, was found to be greater in the animals with a lower gasping time. In these susceptible animals, malondialdehyde was found to be almost 8-fold higher (24.55±1.08 nmol/mg protein, p<0.001) as compared to unexposed animals (3.15±0.33 nmol/mg protein) as well as the tolerant animals (5.20±0.35 nmol/mg protein) (Fig. 2b). Carbonyl derivative formation was elevated in the susceptible animals (9.21±0.64 nmol/mg protein, p<0.001), with reference to the control (1.91±0.14 nmol/mg protein). The increase with respect to the tolerant (4.02±0.26 nmol/mg protein) and normal group (5.42±0.36 nmol/mg protein) was also significant (p<0.05, Fig. 2c). On measuring the levels of AOPPs in the myocardial tissue, a significantly higher level of oxidation was observed in the susceptible group (11.39±0.77 μmol/L, p<0.001) and in animals showing a normal range for the onset of gasping (9.59±0.38 μmol/L, p<0.05) as compared to control (4.83±0.23 μmol/L) (Fig 2d). In comparison, the formation of oxidized proteins was significantly lower in the animals tolerant to hypoxic stress (6.72±0.17 μmol/L, p<0.05).
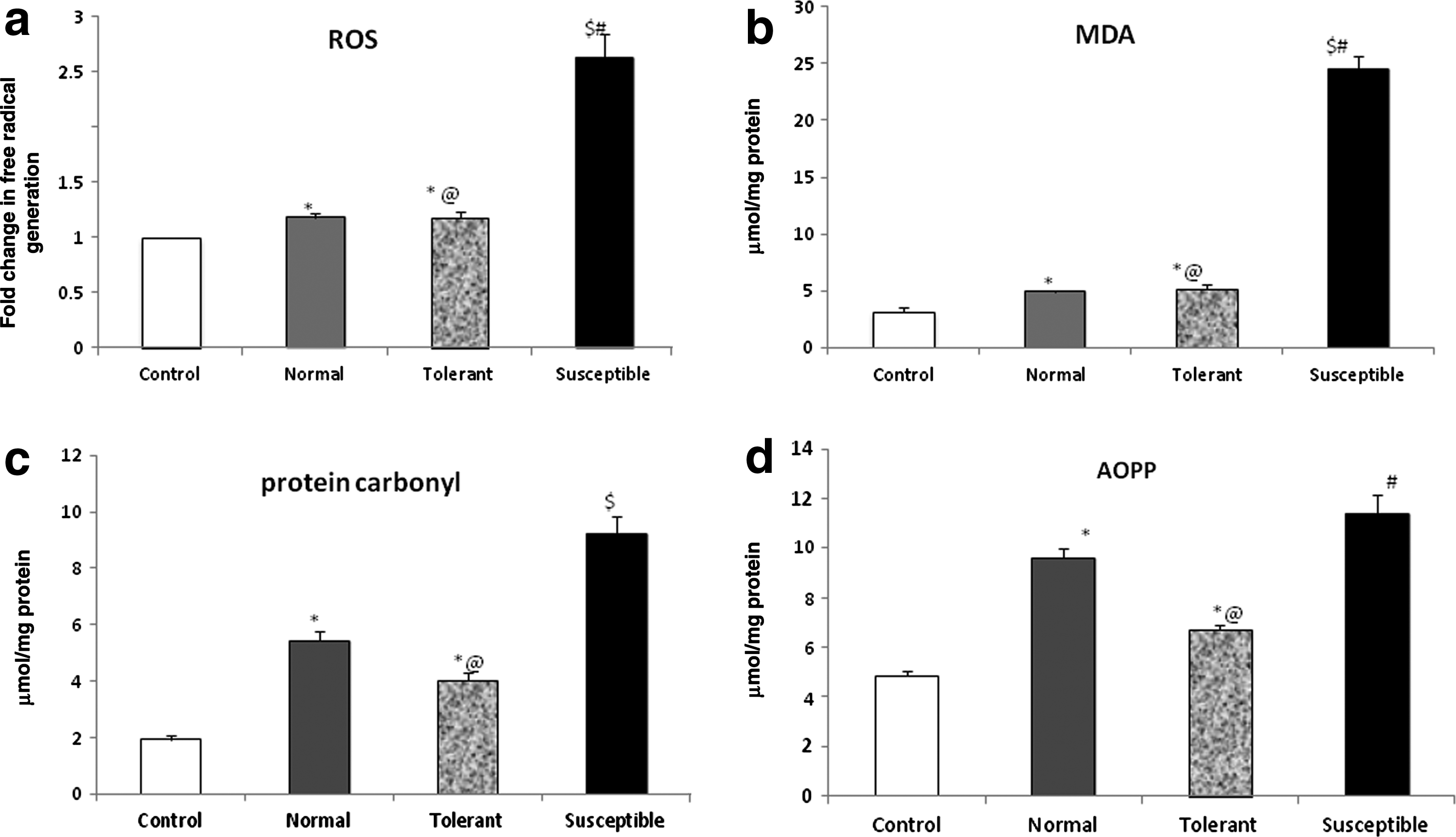
Hypoxia-induced oxidative damage.
Antioxidant levels
Antioxidant enzymes such as SOD and catalase protect the cells against oxidative stress-induced damage. SOD catalyzes the conversion of the superoxide anion (O2−), which is the first product of O2 radicals, to H2O2 and O2. In our study (Fig 3a), we found a higher level of the enzyme in the tolerant group of animals, 2.28±0.12 U (mg protein)−1, p<0.001, as compared to the control as well as the susceptible animals (1.33±0.11 and 1.51±0.05 U (mg protein)−1, respectively). Catalase, which catalyzes the breakdown of H2O2, was almost 3-fold elevated in the tolerant group (11.63±0.66 mmol/min/mg protein, p<0.001) as compared to the control unexposed animals (4.076±0.18 mmol/min/mg protein) (Fig.3b). While elevation in catalase activity was seen in the susceptible and normal group as well, the increase from control was less significant, (5.98±0.27 and 7.67±0.49 mmol/min/mg protein, p<0.05 respectively).

Antioxidant enzymes and cellular defense system.
Levels of reduced glutathione in the exposed myocardium from susceptible animals were not significantly altered with respect to its levels in the control group (Fig. 3C). The tolerant animals had relatively higher levels of this intracellular antioxidant, 202.31±17.74 nmol/mg protein as compared to control, 133.12±10.02 nmol/mg protein (p<0.001).
Modulators of vascular tone
Hypoxic vasodilation is a well-known response to reduced oxygen availability. Nitric oxide is a potent vasodilator and may contribute to increased hypoxic tolerance. In our study (Fig. 4a), we found a significant enhancement in levels of NO in animals tolerant to hypoxic stress, almost 2-fold, as compared to control (2.77±0.17 vs. 1.36±0.04 nmol/mL, p<0.001). The increase seen in the susceptible animals was relatively less (1.74±0.06 nmol/mL, p<0.01). Increased nitric oxide availability opposes endothelin-1(ET-1) mediated vasoconstriction, thereby modulating a balance between systemic vasodilation and pulmonary vasoconstriction under hypoxic conditions. In our study, we observed a highly significant 30-fold increase in the tissue ET-1 expression in the susceptible animals, 699.57±38.35 pg/mL, p<0.001 as compared to exposed tolerant and control unexposed animals,31.83±1.79 and 23.63±1.81, respectively (Fig. 4b).
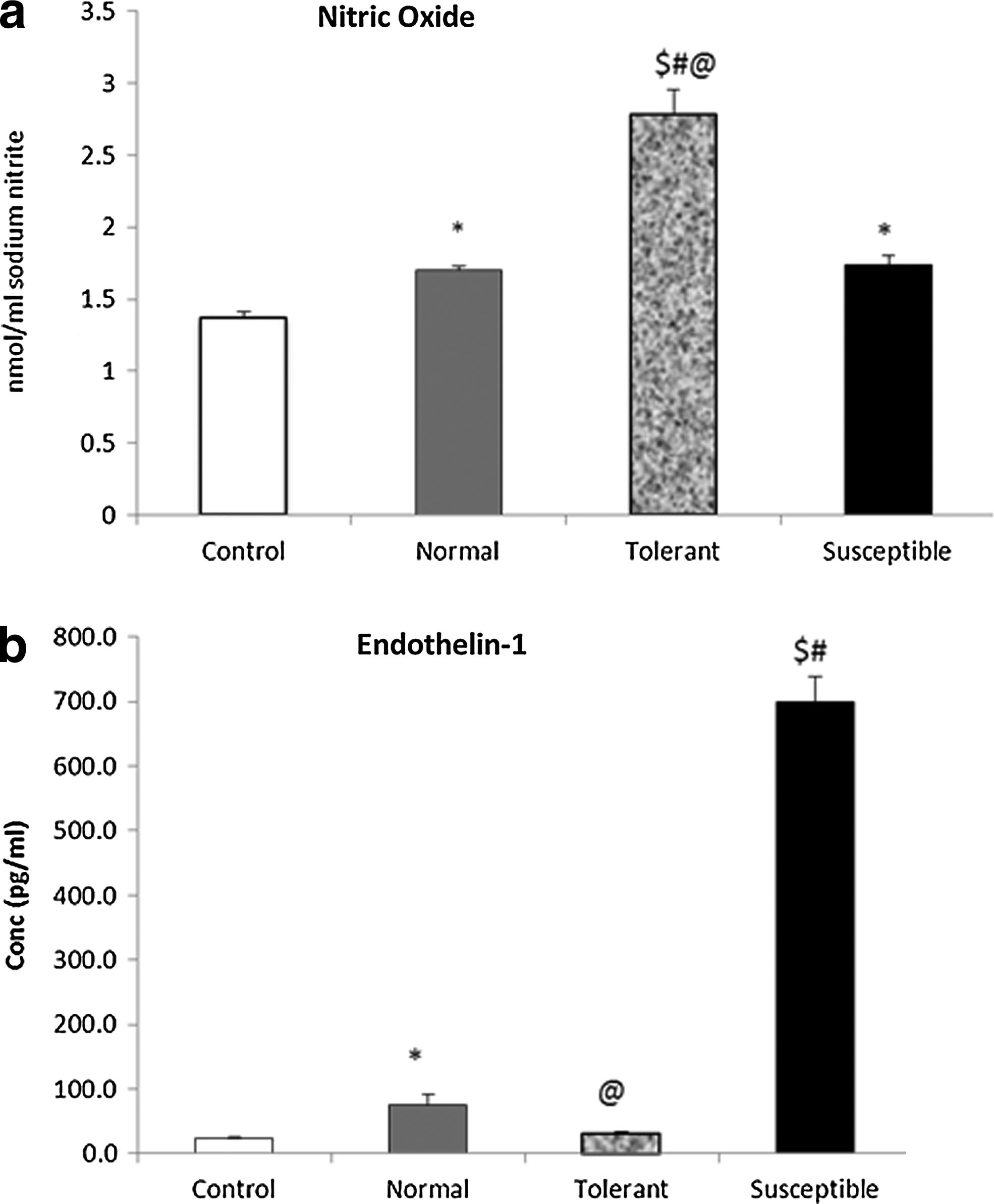
Modulators of vascular tone.
Hypoxia-inducible factor-1α and expression of hypoxia-responsive proteins
A key player in the cellular response to hypoxia is hypoxia-inducible factor-1α which mediates the physiological activation of a variety of responses to oxygen deprivation. In our study, we measured the levels of HIF-1α in the nuclear myocardial fractions. We observed a 2-fold increase in the expression levels in the tolerant group of animals, with reference to control, while the HIF1-α upregulation in the susceptible group was comparable to normal animals (Fig. 5a). Various proteins such as those involved in glucose metabolism and transport, angiogenesis as well as erythropoiesis, are regulated by the expression of HIF-1α. We studied the expression of glucose transporters, GLUT1 in the myocardial tissue, and observed a 1.5-fold increase in the tolerant group while the normal range and susceptible animals showed a similar level of elevation from control (Fig. 5b).
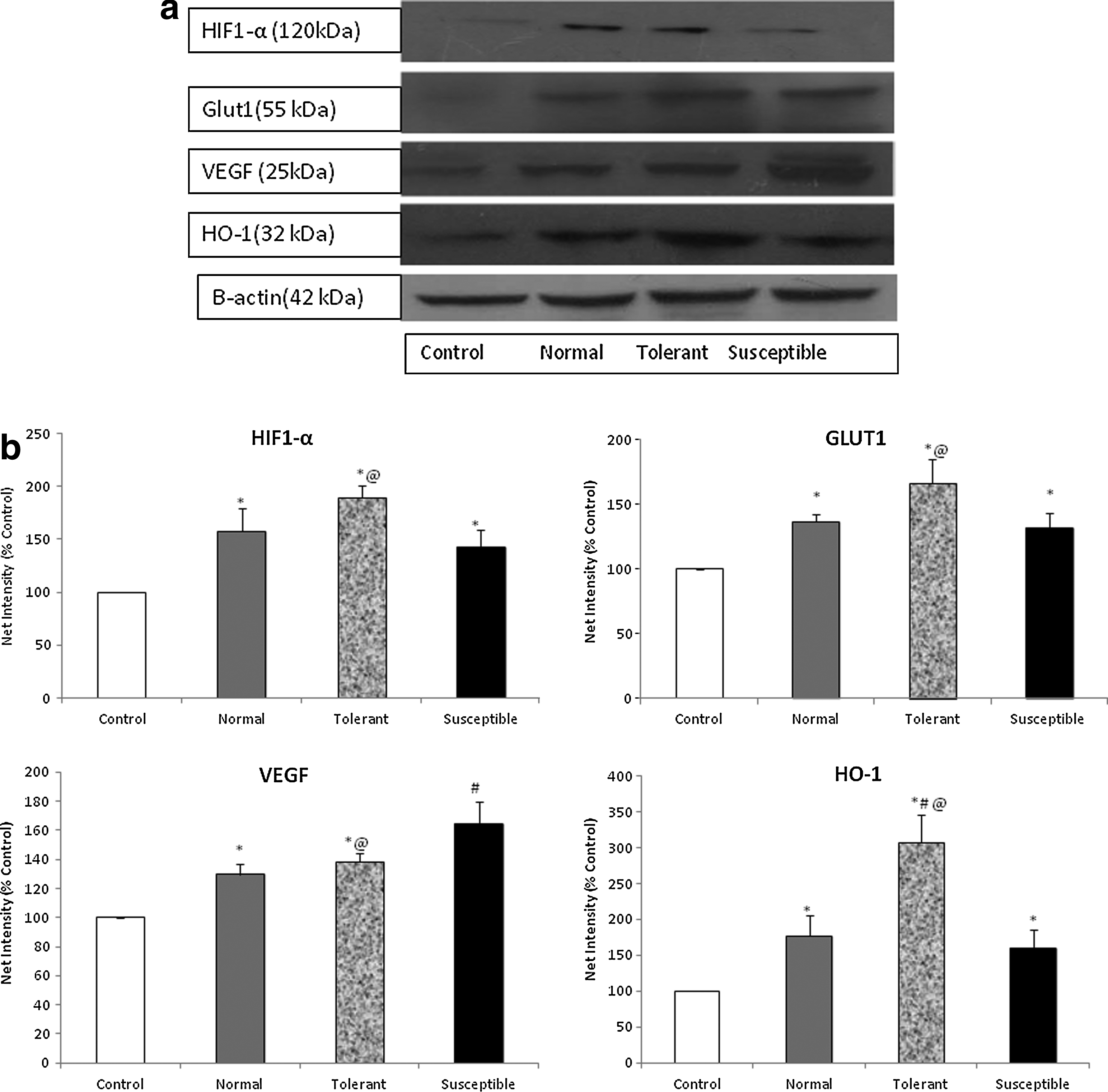
Differential expression of hypoxia-inducible factor-1α and its target proteins in response to acute sublethal hypoxic stress.
A diverse set of hypoxia responsive proteins, including those regulated by HIF-1α, collectively play a significant role in mediating systemic, local, or intracellular O2 homeostasis. Erythropoietin, regulated essentially by HIF-1α, augments the blood O2-carrying capacity by stimulating erythropoiesis. In our study we observed a slight increase in erythropoietin (EPO) levels in tolerant group of animals (470.53±25.44 pg/mL) as compared to control (382.35±13.3pg/mL) (Fig. 6a). Interestingly, the susceptible animals showed a fall in the myocardial erythropoietin levels (354.63±13.54 pg/mL, p<0.05). Heme oxygenase-1, a redox sensor and modulator of vascular tone, showed a 2-fold increase in its levels in the tolerant group, 4.29±0.19 ng/mL, p<0.005 vs. unexposed animals, 2.04±0.27 ng/mL (Fig. 6b). A significant increase was also seen in the myocardium from susceptible animals, 3.18±0.24 ng/mL, p<0.05. These results were substantiated by immunoblotting with anti-HO-1 antibodies, where a similar 2.5-fold increase was seen in the tolerant group. Measuring the levels of angiogenesis marker, vascular endothelial growth factor, VEGF, we found a 2-fold increase in the susceptible group of animals as compared to unexposed (399.57±33.49 vs. 177.40±9.0 pg/mL, p<0.005) (Fig. 6c). The increase in tolerant and normal range groups was not significant. A similar trend of results was observed on immunoblotting for these hypoxia responsive proteins.
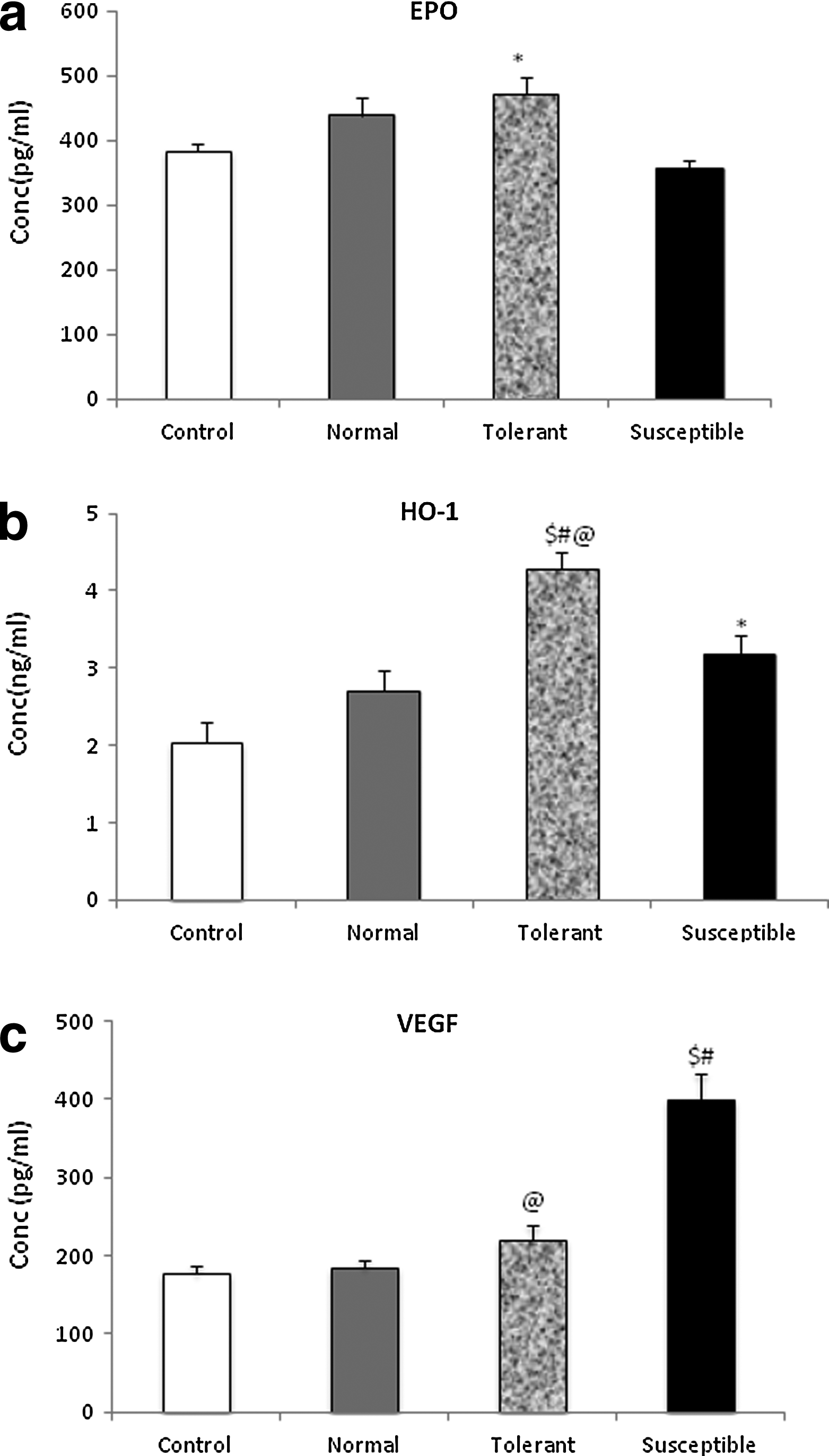
Quantitative determination of expression of hypoxia responsive proteins.
Expression of stress proteins
Heat shock proteins play a crucial cytoprotective role in response to environmental and metabolic stress. In our study we identified the expression of the major stress proteins in the myocardial tissue and we observed an increase in cytosolic expression of HSP60, HSP70, as well as HSP90 (Fig. 7). Tolerant animals showed higher expression of the major cytoprotective chaperone HSP70, the first chaperone to be activated under cellular stress, the increase as compared to control being almost 1.5-fold. While a higher expression of the chaperone was also seen in the normal and susceptible groups, as compared to tolerant, the increase was not significant. The expression of HSP60, measured in the cytosolic fraction, showed almost a 2.5-fold increase in the tolerant animals, whereas the increase in the normal and susceptible groups, from the unexposed animals, was comparatively less. HSP90, a major regulator of different transcription factors including HIF-1α, was also found to be expressed more significantly in the tolerant groups, as compared to the other two groups of exposed animals.
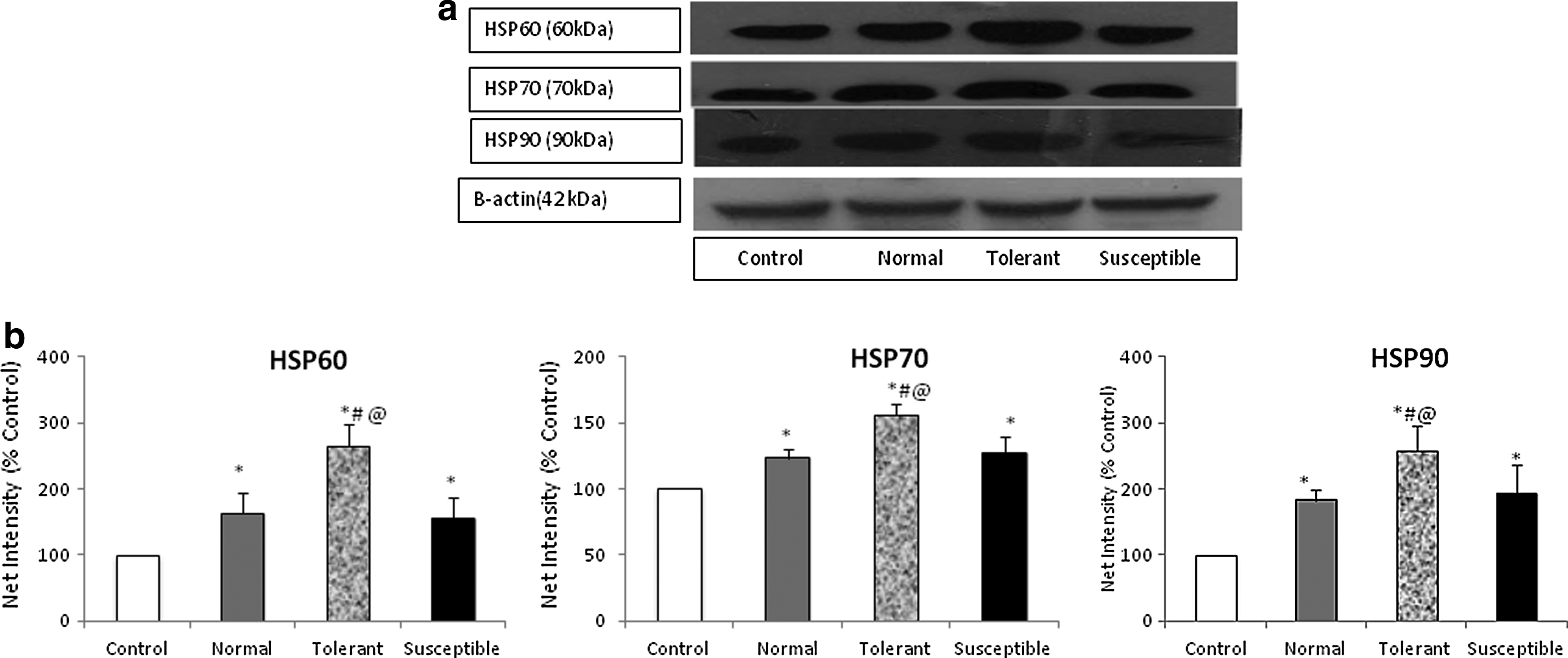
Acute hypoxic stress enhances the expression of cytoprotective heat shock proteins in the myocardium.
Discussion
The environment at high altitude is extreme, characterized by hypobaric hypoxia, yet people have inhabited it since ancient times. There is evidence of adaptive traits in highlanders (Moore et al., 1998), which makes their survival and performance easier under hypoxia; on the other hand, survival of sojourners is rendered difficult at HA. Identification of specific molecules that impart tolerance or susceptibility to such stressors would play a significant role in screening of individuals for survival in such challenging conditions. Our study highlights the importance of some such biochemical markers in the heart, which may be important for predicting the tolerance/susceptibility to hypoxic stress. Our observations indicate that an enhanced tolerance to stress can be contributed collectively to an inherently better cellular defense system, as well as a better ability to mount a cytoprotective response under hypoxia. We find that tolerant animals, capable of maintaining normal function for longer durations under acute stress, were relatively better armed, as evidenced by elevated antioxidant enzymes, stress proteins, as well as hypoxia responsive proteins, to combat the free radical generation and showed a lower degree of protein and lipid damage as compared to animals susceptible to acute hypoxic stress.
Hypoxia induces a number of physiological responses to compensate for the decrease in oxygen availability. A decrease in the arterial blood pO2, due to hypobaric hypoxia, can lead to increased pulmonary ventilation to facilitate an increase in oxygen uptake. The initial increase in ventilation has been found to be mediated by the carotid bodies, which are oxygen-sensitive cells that increase their activity in such conditions. Severe hypoxia has been known to directly enhance the excitability of neurons within the rostral medulla “gasping center” “pre-Botzinger complex” resulting in augmentation and then a decline in the rhythmic bursts, characteristic of gasping (St. John and Knuth, 1981; St. John, 1996). On exposure to acute hypoxia, equivalent to 32,000 ft or 9754 m, we observed a difference in the onset of gasping or hyperventilation in the animals. The animals were exposed to such extreme simulated altitudes, because smaller animals such as rodents have higher capillary density in tissues, making them more resistant to hypoxia than man. Previous studies from our laboratory have shown the effectiveness of this model to identify the effect of preconditioning with cobalt chloride and herbal supplementation on enhancing the hypoxic tolerance by increasing the time for onset of gasping, and hypoxic survival time (Kumar et al., 1999; Jayamurthy et al., 2008; Shrivastava et al., 2008). In our study, we used the onset of time of gasping as an efficient indicator for the ability of the animal to tolerate the stress. While most animals began gasping within 15–25 min (54%), few were able to survive for even longer than 30 min, indicating an increased tolerance to stress (27%). Some animals were very susceptible to hypoxic stress, in which the onset of gasping was observed in less than 10 min (18%).
On staining the myocardial sections with hematoxylin eosin stain to assess the degree of damage, neutrophil infiltration and edema were seen in the sensitive animals. While minimal damage was seen in the tolerant group, with normal myocyte architecture, the normal range animals showed mild infiltration. Thus while the exposure to acute stress might be shorter in duration in susceptible animals, they are highly vulnerable to damage. On the other hand, even under relatively longer exposure time, the tolerant animals are able to withstand the hypoxic stress and resist damage. Creatine kinase, present in variable isoforms in heart muscles, catalyzes the reversible transfer of a phosphate moiety between ATP and creatine and thereby acts efficient energy shuttles. A failing or damaged heart is unable to maintain its energetic reserves and shows altered levels of high energy phosphates. CK-MB isoform is a well-known marker for cardiac injury and damage and in our study was found to be significantly elevated in the susceptible group of animals. Previously, oxidative stress has shown the elevation in the levels of this enzyme; indeed a number of studies have highlighted the enzyme activity as a marker for myocardial infarction and ischemia reperfusion injury (Ingwall, 1993; Nascimben et al., 1996); an altered creatine kinase isoenzyme profile has also been observed in rats native to high altitude (Letout et al.,2005).
To identify the effect of hypobaric hypoxia on the redox status in myocytes, we measured the levels of ROS generation in the tissue and observed that while the normal and tolerant animals showed relatively less increase in the ROS levels, the animals susceptible to hypoxic stress showed a significantly greater production of such reactive species. It follows that the damage mediated by these highly reactive molecules would be maximal in the susceptible animals. In congruence with our hypothesis, we found that the damage to proteins, as evidenced by the almost 4-fold increase in carbonyl derivative formations and oxidized protein products in the same group. While an increase was also seen in the normal and tolerant animals, this was in concordance with the degree of increase in oxidative stress. A similar fold increase was observed in lipid peroxidation products in the susceptible group, a consequence of oxidative stress that was to a much greater extent than in the other two exposed groups. Previously, an increase in ROS formation under hypobaric hypoxia has been observed in a number of studies; Radak et al. (1997, 1998) have shown an increase in carbonyl derivative formation in skeletal muscle under high altitude stress.
To combat the excessive production of ROS, organisms have a cellular defense system consisting of antioxidants and antioxidant enzymes. The major antioxidant enzymes, catalase and superoxide dismutase, were elevated in the tolerant group. This point towards an intrinsically higher level of antioxidants in these animals, which may help in inhibiting hypoxia-induced oxidative stress. Superoxide dismutase, considered as the first line of defense in the free radical scavenging pathway, has been shown to be higher in high altitude sojourners as compared to lowlanders. A similar upregulation in catalase and glutathione peroxidise activity has also been seen in the sojourners as compared to natives who are exposed to high levels of free radicals throughout their lives and have developed adaptive mechanisms (Sinha et al.,2009). Several studies have shown that natives of HA areas are well adapted to their hypoxic surroundings (Beall, 2007; Wu and Kayser, 2006; Xing et al., 2008). Most studies have highlighted the different strategy adopted by lowlanders and mountain populations to cope with oxidative stress, which is the most harmful consequence of chronic hypoxia (Gelfi et al., 2004). A better antioxidant defense system such as seen in the tolerant organisms may be capable of ameliorating the high altitude-induced oxidative stress.
Nitric oxide (NO) and endothelin-1 (ET-1) are endothelium-derived mediators that play important roles in vascular homeostasis. NO, because of its competition with O2, acts as an inhibitor of cellular respiration to a large extent at low O2 tensions and may thus protect cellular functions under hypoxia by extending O2 availability while modulating the pulmonary vascular tone (Hagen et al., 2003). In our study we have observed a significant 2-fold increase in the nitric oxide levels in tolerant animals, while the increase in the susceptible and normal range animals is negligible. The increased nitric oxide levels may be a contributory factor in enhancing the ability to sustain normal respiratory ability for a longer duration even under the impact of acute sub-lethal hypoxia. On the other hand, endothelin, which regulates the pulmonary vascular tone under hypoxic stress, was almost 30 times higher in the susceptible animal tissues. Within the myocardium, ET-1 has been shown to be an inducer of cardiac hypertrophy and is associated with the development of cardiac dysfuntion (Yamazaki et al., 1996; Bupha-Intr et al., 2012). Interestingly, higher NO synthesis attenuates oxidative stress, thereby facilitating hypoxic pulmonary vasoconstriction and inhibits the production of endothelin from the vasculature. In humans, high altitude has been shown to induce the hormone and increased ET-1 synthesis may contribute to exaggerated hypoxic pulmonary hypertension in HAPE-prone subjects (Kourembanas et al., 1993; Goerre et al., 1995).
Hypoxia elicits a cascade of functional and structural modifications in the physiological systems that govern the supply and demand of oxygen in organisms. A major physiological response to oxygen deprivation in mammals is mediated by the hypoxia responsive nuclear factor, HIF-1α. While degraded under normoxic conditions, HIF is recognized as a key modulator of the cellular transcriptional response to hypoxic stress (Wang and Semenza, 2003). Its stabilization under low oxygen results in the activation of a transcriptional program with effects on metabolism, redox homeostasis, vascular remodeling, inflammation, and other processes (Salceda and Caro, 1997; Chavez et al., 2000). Results of our study show that tolerant animals show a greater stabilization of HIF-1α as compared to susceptible and normal animals. Ventilatory acclimatization to hypoxia, a critical physiological adaptation following HIF-1 α stabilization, is augmented in animals and humans displaying a higher expression of this nuclear factor (Pialoux et al., 2009). Identifying a prominent role for HIF-1α in the hypoxic ventilatory response, studies on heterozygous HIF-1α knockout mice have shown a significantly diminished O2-sensitivity in the carotid bodies of such mice (Kline et al., 2002).
Under acute hypoxic stress, HIF-1α induces several hematological modifications that enhance oxygen transport in the blood and facilitate the unloading of oxygen at the tissues. Erythropoietin, responsible for increasing the oxygen carrying capacity, was found to be lower in the susceptible animals. The hormone, known to modulate the hypoxic ventilatory response (Soliz et al., 2005), was increased in tolerant animals. Heme oxygenase-1, an effector molecule of HIF-1α, involved in maintenance of the redox homeostasis, was 2-fold higher in the myocardium of tolerant animals. The enzyme, a member of the heat shock protein family (HSP32), is also induced directly by oxidative stress, nitric oxide, and hypoxia (Tyrell, 1999; Kalmar and Greensmith, 2009). It can be postulated that while the tolerant animals had a limited intracellular ROS production, presence of elevated HO-1 and antioxidant levels allowed the animals to mount a rapid response to decreased oxygen availability. HIF-1α stabilization also regulates VEGF, higher expression of which increases capillary density and angiogenesis. VEGF levels have been shown to be augmented in lungs and plasma in pulmonary edema and acute mountain sickness (Maloney et al., 2009); ischemia and chronic hypoxia upregulate its expression in the myocardium (Banai et al., 1994; Hashimoto et al., 1998). Results of our study show a significantly higher VEGF expression in animals with higher susceptibility to hypoxic stress, substantiating a link between ROS–HIF-1–VEGF pathways, as hypothesized earlier by Irwin et al., 2009.
Oxygen delivery under hypoxic conditions, along with vasodilatory and ventilatory responses, is further aided by a shift in fuel preferences, such as an increased reliance on glucose oxidation. In our study, we detected an enhanced expression of glucose transporter, GLUT1 in tolerant animals, capable of coping with acute hypoxic stress, as compared to other hypoxia-exposed animals. A large number of previous studies, (Behrooz and Ismail-Beigi, 1997; Wenger, 2002; Bateman et al., 2007), have identified the HIF-1α mediated regulation of glucose transporters. GLUT1itself has been established as an indicator for the effective shift from aerobic to anaerobic metabolism in hypoxia exposed myocardium (Sivitz et al., 1992).
Molecular chaperones play a prominent role in physiological responses to environmental stress. In our study, we found an elevated expression of the major members of the heat shock protein family, the increase being more pronounced in the tolerant animals. This observation suggests that while hypoxia-induced oxidative stress is capable of activating a heat shock response, better chaperone activity, preventing protein misfolding and impaired function, aided their ability to coping with stress. Studies by Dillmann and Mestril (1995) have proposed a positive correlation between stress protein expression, especially HSP70, and protection against myocardial damage. HSP70, along with HO-1, forms the first line of defence against oxidative injury, acting as efficient redox sensors (Calabrese et al., 2004). Indeed, preconditioning-induced HSP70 expression has been shown to not only attenuate acute lung injury under hypobaric hypoxia (Lin et al., 2011), but also to play a cardioprotective role, preventing ischemic injury and myocardial damage (Suzuki et al., 2000).Downstream upregulation of other members of chaperone family, such as HSP60 and HSP90, occurs through HSP70-mediated action of heat shock factor-1. While HSP60 has previously been shown to play an antiapoptotic role in myocardial injury (Kirchhoff et al., 2002), HSP90 was recognized as a regulator of HIF-1α stability (Minet et al., 1999; Isaacs et al., 2002). Earlier studies on various species have shown the link between the activation of HSP90, HIF-1α, and eNOS in the cellular decision to survive or succumb to stress (Chen and Meyrick, 2004). Collectively, these responses mediate tolerance to the effects of hypoxia induced oxidative stress in the myocardium. These cellular responses play a central role in maintaining cardiac function under reduced oxygen tensions, while protecting it from free radical induced damage.
Conclusion
Although the molecular mechanisms of hypoxia and high altitude acclimatization are being identified and primarily include information about hypoxia-induced factors (HIFs) and their role in the high altitude response (Clerici and Planes, 2009), the signaling pathways and molecular functions that mediate such responses remain largely unknown. In conclusion, while the precise basis for each organism's distinct ability to cope with stress has not been elucidated, our study provides knowledge on biochemical markers for such differences. We present evidence that a differential induction and stabilization of the HIF-1α activates a downstream pathway responsible for the regulation of a variety of genes. Such genes including those involved in increased oxygen transport and delivery (EPO, and HO-1), vascular tone modulators and angiogenesis (ET-1 and VEGF), and glucose utilization (GLUT1) regulate the degree of tolerance an organism has to acute hypoxic stress. A key role is also played by the intracellular antioxidant defense machinery, as well as the molecular chaperone system, which contribute to the reduced vulnerability to stress and damage in the tolerant VEGF and HO-1 animals. While the present study identifies these biochemical and protein responses in the heart, further more extensive studies are required to evaluate these molecules as markers for screening of individuals for resistance or susceptibility to environmental stress (Fig. 8).
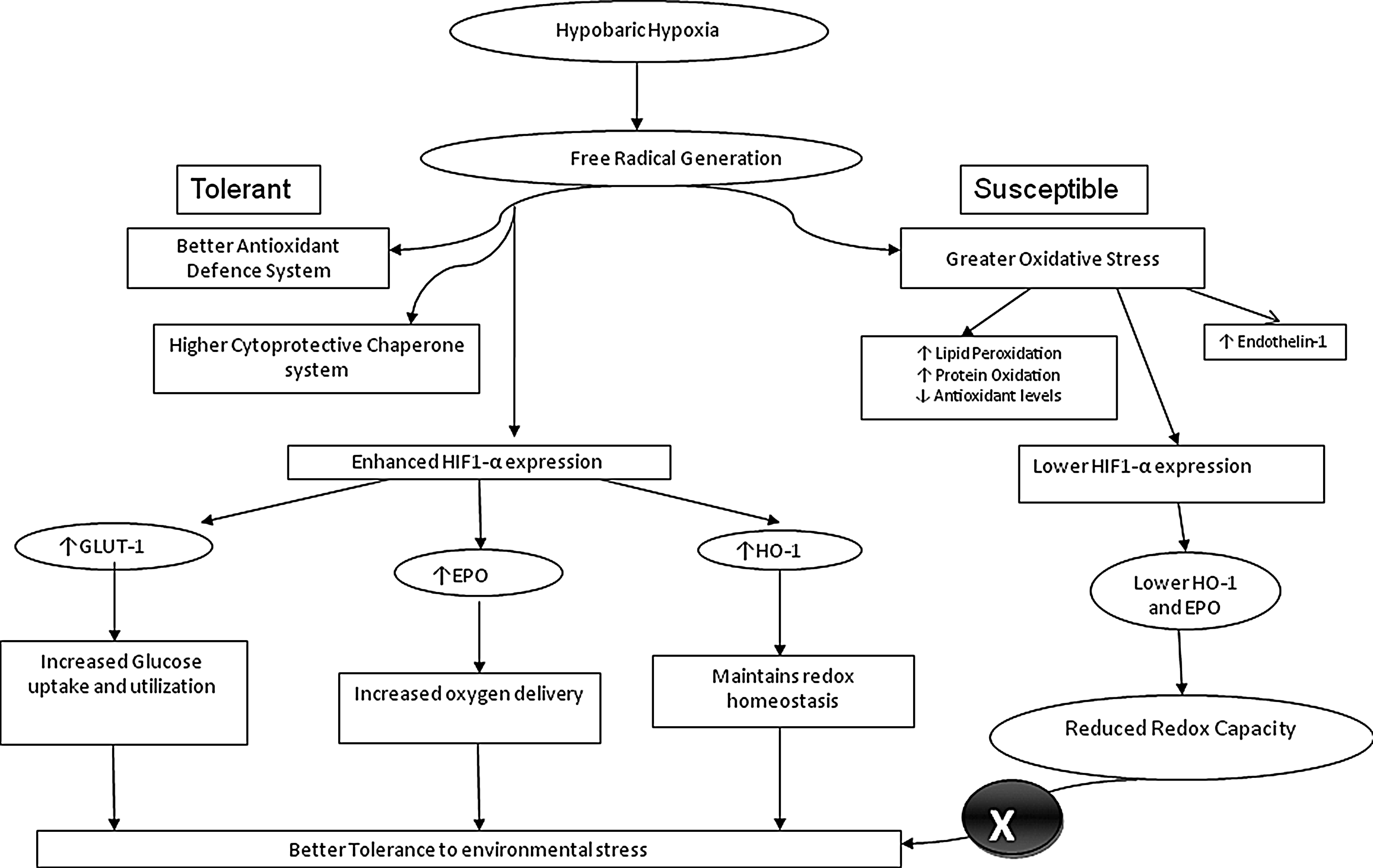
Differential myocardial response for enhanced tolerance to acute hypoxic stress.
Footnotes
Acknowledgement
The study was supported by Defence Research and Development Organisation, Ministry of Defence, Government of India. The first author is a Council of Scientific and Industrial Research Senior Research Fellow.
Author Disclosure Statement
The authors state that there is no personal or financial conflict of interest in the present study.