
Abstract
Abstract
Serebrovskaya, Tatiana V., Valentyna I. Nosar, Larissa V. Bratus, Bronislav L. Gavenauskas, and Iryna M. Mankowska. Tissue oxygenation and mitochondrial respiration under different modes of intermittent hypoxia. High Alt Med Biol 14,280–288, 2013.—We compared the results of five modes of intermittent hypoxia training (IHT) on gastrocnemius muscle P
Introduction
On the other hand, the intermittent hypoxia research in Western Europe and North America was primarily focused on the detrimental effects of chronic intermittent hypoxia associated with sleep-disorder breathing. For instance, Prabhakar and Semenza (2012) consider that intermittent hypoxia almost always represents a pathological stimulus that evokes maladaptive responses. However, during the past decade, such a gap of division between East and West is progressively shrinking, and mutual understanding on what “intermittent hypoxia” means, becomes clearer (Semenza, 2012; Serebrovskaya and Xie, 2009).
The questions that arise are, what are the key mechanisms determining the adaptive versus maladaptive nature of different paradigms of intermittent hypoxia, and, what molecular pathways are mediating the observed pathological or physiological response? It is appropriate to mention here the ancient wisdom well expressed by Paracelsus in the XYIth century: “Sola dosis facit venenum” (only the dose makes the poison).
Until now, there is no exact evidence about the precise mechanism for switching adaptive or maladaptive responses to hypoxic impact. Several attempts were undertaken to analyze this question (Lukyanova, 2005; Lukyanova et al., 2009, 2012; Serebrovskaya et al., 2008; Yin et al., 2012). The most significant contribution to this question was made by Prabhakar and Semenza (2012), who described transcriptional regulation of gene expression mediated by hypoxia-inducible factors 1 and 2 (HIF-1 and HIF-2). These factors constitute important components of the genetic makeup in the body, influencing hypoxic sensing by regulating intracellular redox state via transcriptional regulation of pro-and antioxidant enzymes. The large number of known feedback loops underscores the critical importance of precisely regulating O2 delivery and utilization. The basic message of the authors is that HIF signaling is not a linear pathway, but a complex web with perhaps hundreds of input stimuli and thousands of potential output responses, with each representing a different target gene. The discovery of the HIF family has led to novel insights into the molecular basis of adaptive and maladaptive cellular and systemic responses to continuous and intermittent hypoxia.
While these fundamental molecular studies on cell culture models provided important insights into mechanisms of HIF activation by hypoxia, they cannot answer as yet, the practical question of what dose and regimen of hypoxic impact could be mostly beneficial for animals and humans. In practice, hypoxic regimens that are used for the study of hypoxic adaptations vary broadly from 3–12 short hypoxic sessions (2–10 min) with 2–20 min normoxic breaks during 7–30 days to hypoxic influences lasting from 1–12 hours during 2–90 days (Fagan, 2001; Joyeux-Eaure et al., 2005; Lin et al., 2002; Manukhina et al., 2011; McGuire and Bradford, 2001; Naryzhnaia et al., 2009; Nattie et al., 1978; Neckár et al, 2002; Rozova et al., 2012, Vavilova et al., 2005; Zhu et al, 2004; Vavilova et al., 2005).
This wide spectrum of protocols for IHT is represented now in literature showing both beneficial and detrimental effects. In addition, simply the mode of hypoxic influence (depth, duration, and intermittence) appears to be critical for the determination of healing or harmful result. Therefore, special purposeful investigations are needed to elucidate basic mechanisms of different IHT effects depending on the modality of hypoxic stimuli and to elaborate the most effective and safe regimen for the introduction in human practice.
The two most informative parameters of hypoxic influence on an organism are tissue oxygenation and tissue respiration. At the tissue level, hypoxia activates the transcription of multiple genes encoding angiogenic growth factors and cytokines. The result is an increase in the capillary density and tissue perfusion that increases in tissue oxygenation (Faiss et al., 2013; Mankovska et al., 2005; Panisello et al., 2008; Xu et al., 2011). At the cellular level, hypoxia leads to reprogramming of mitochondrial metabolism that ensures adequate ATP generation and prevents adverse consequences of excess mitochondrial ROS generation. These metabolic adaptations are due to HIF-1-mediated transcriptional regulation of glycolytic enzymes, mitochondrial electron transport chain components, and other metabolic enzymes (Prabhakar and Semenza, 2012). Many studies confirm the close relations between physical performance and mitochondrial respiration during adaptation to intermittent hypoxia (Mankovska et al., 2005; Saxena et al., 2012; Zoll et al., 2002).
Based on the above summary of current knowledge, the aim of this study was to compare the effects of the five most widespread modes of hypoxic training on rat muscle P
Materials and Methods
Animals
All procedures followed the criteria, technical standards, and rights applied to animal research. All trials were followed in accordance with the statements of the European Union regarding handling of experimental animals. This investigation conforms to the law and local ethical committee guidelines for animal research. Experiments were conducted on adult male Wistar rats (weight range 200 to 220 gram). The animals were housed (4 per cage) in a room with 12:12-h light-dark cycle at 22°C and were provided with standard rat chow with water ad libitum.
Experimental design
Two series of experiments were performed. In Series I, muscle oxygen partial pressure (Pm
In Series II, the investigation of liver and myocardium mitochondrial respiration was completed after a course of the two most beneficial IHT modes. In total, 91 animals were used in the study.
Series I: Measurements of muscle Po2
Each of the five modes was studied in a separate group of animals consisting of 7 rats. Each rat was exposed six times to the chosen mode during one test day (Table 1). Animals of Gr. I-1 were furthermore subjected to 3-week IHT in Mode 1, and Pm
Protocol
All manipulations were made under light etherization. Animals were fixed in supine position. Rats breathed through a two-valve respiratory mask that could feed gas mixtures directly to the nostrils, thus minimizing the dead space. A mask covered the animals' eyes protecting them from outside irritants. Investigations included Pm
To determine Pm
Series II: Study of mitochondrial respiration
Animal groups
Eight groups of rats, each group with 7 animals, participated in this series. Among them, group II-1 was the control group, exposed to a sham IHT, and five groups (Gr.II: 2–6) were exposed to IHT courses of different durations, Mode 1:
Gr.II-1: control group;
Gr.II-2: IHT during 7 days;
Gr.II-3: IHT during 14 days;
Gr.II-4: IHT during 21 days;
Gr.II-5: 2 month residence in vivarium after 3-week IHT;
Gr.II-6: 3 month residence in vivarium after 3-week IHT.
Gr.II-7 had sham treatment with IHT, Mode 2 (control group), and Gr.II-8 underwent to 3-week IHT in Mode 2.
IHT protocol
Rats underwent the IHT sessions in two gaseous chambers: hypoxic (ventilated by hypoxic gaseous mixture with 12% O2) and normoxic (ventilated by ambient air). Animals were placed inside a chamber in a metal cage that could be quickly moved from the air chamber and placed into the hypoxic chamber (there and back) every 5 or 15 min depending on the IHT mode. This procedure was repeated five times daily during 7, 14, or 21 days, depending on group number. The level of O2 fraction in the hypoxic chamber was monitored by a gas analyzer.
Mitochondrial respiration protocol
On the next day after the last hypoxic exposure, rats were anesthetized with 1–2% isoflurane, decapitated, and their livers and hearts were extracted for further analysis of mitochondrial respiration. Mitochondria were isolated by differential centrifugation according to Kondrashova et al. (2001). Briefly, rat hearts and livers were quickly excised, washed in ice-cold buffer, weighed, and homogenized in a glass Potter-Elvehejm homogenizer with a motor-drive Teflon pestle. The homogenization medium for heart contained 120 mM KCl, 10 mM HEPES, 10 mM EDTA, and 0.5% bovine serum albumin; pH 7.2; for liver: 120 mM KCl, 2 mM K2CO3, 10 mM Tris HCl, 1 mM EGTA; pH 7.2. Homogenate was centrifuged for 7 min at 700 g at 4°C, then supernatant was centrifuged for 15 min at 11,000 g at 4°C. Mitochondrial suspension (3–4 mg protein/mL) was slurred in a medium without EDTA and kept on ice at 4°C.
Mitochondrial respiratory function was measured in a water-jacketed chamber using a Clark O2 electrode by the polarographic method of Chance and Williams (1956). Mitochondria were added to the thermostated and magnetically stirred respiration chamber containing a total volume of 1.0 mL of respiration medium. Medium composition: for heart—30 mM tris-HCl, 125 mM KCl, 10 mM NaCl, 5 mM KH2PO4, 1.5 mM MgCl2, and 3 mM EGTA; for liver—120 mM KCl, 2 mM KH2PO4, 2 mM K2CO3, 10 mM TrisHCl, pH 7.2 at 25°C. Following oxidative substrates were used: 1 mM succinate in the presence of 2 μM of rotenone (S+R), or 3 mM glutamate+2.5 mM of malate (G+M). 0.2 mM ADP (phosphate acceptor) was administered.
The following parameters were measured and calculated: V4s (oxygen consumption before ADP addition), V3 (oxygen consumption stimulated by ADP), V4 ATP (oxygen consumption after cessation of ADP phosphorylation), respiratory control ratio by Chance (V3/V4 ATP ), and efficiency of phosphorylation (ADP/O, ADP-to-oxygen-ratio) (Estabrook, 1967). Oxygen consumption was recorded as ng atoms oxygen per minute per mg mitochondrial protein, determined by the Lowry assay (Lowry et al., 1951).
Statistical analysis
Results were statistically analyzed with Student's t-test and ANOVA and presented as Mean±SD. Statistical differences were considered significant if the p value was<0.05.
Results
Figure 1 represents the results of Pm
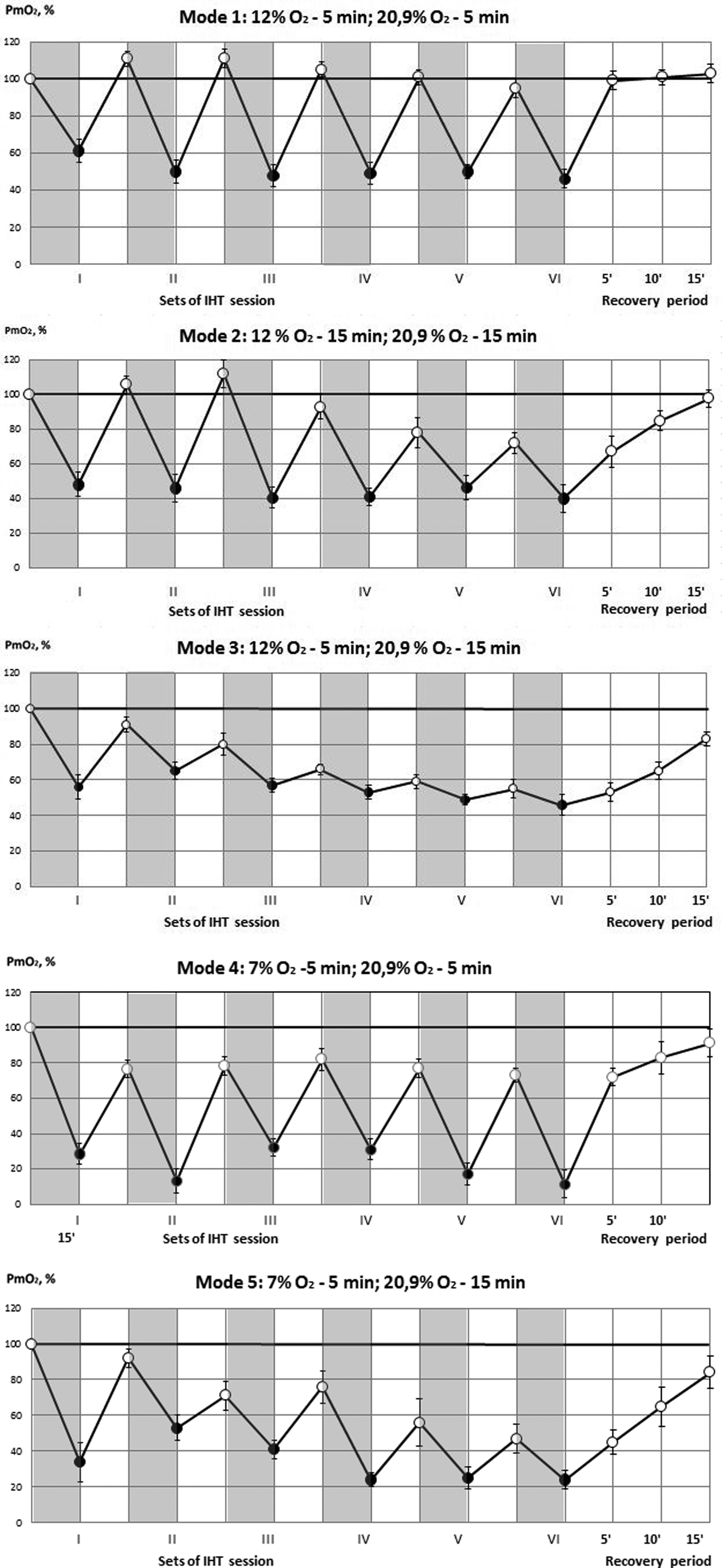
Oxygen tension (Pm
The increase of both hypoxic and normoxic exposures up to 15 min (Gr. I-2), showed similar results as in Gr. I-1 during the first set but provoked a decrease in Pm
The 5-min hypoxic load with 12% O2, but with prolonged breaks of up to 15 min (Gr. I-3), did not lead to the full restoration of muscle oxygen supply after repeated sessions: Pm
More severe hypoxic exposure (7% O2, 5 min) led to more severe drop in Pm
Thus, the most advantageous mode, which provides the least drop in Pm
Since fifth and sixth sets of hypoxia load did not differ significantly in all groups (Fig.1), we used in the sequel five hypoxic sets per day for IHT implementation.
To prove the beneficial effects of IHT with Mode 1, we further subjected animals of Gr.I to a 21-day IHT cycle using this mode. For more evidence, we provided Pm
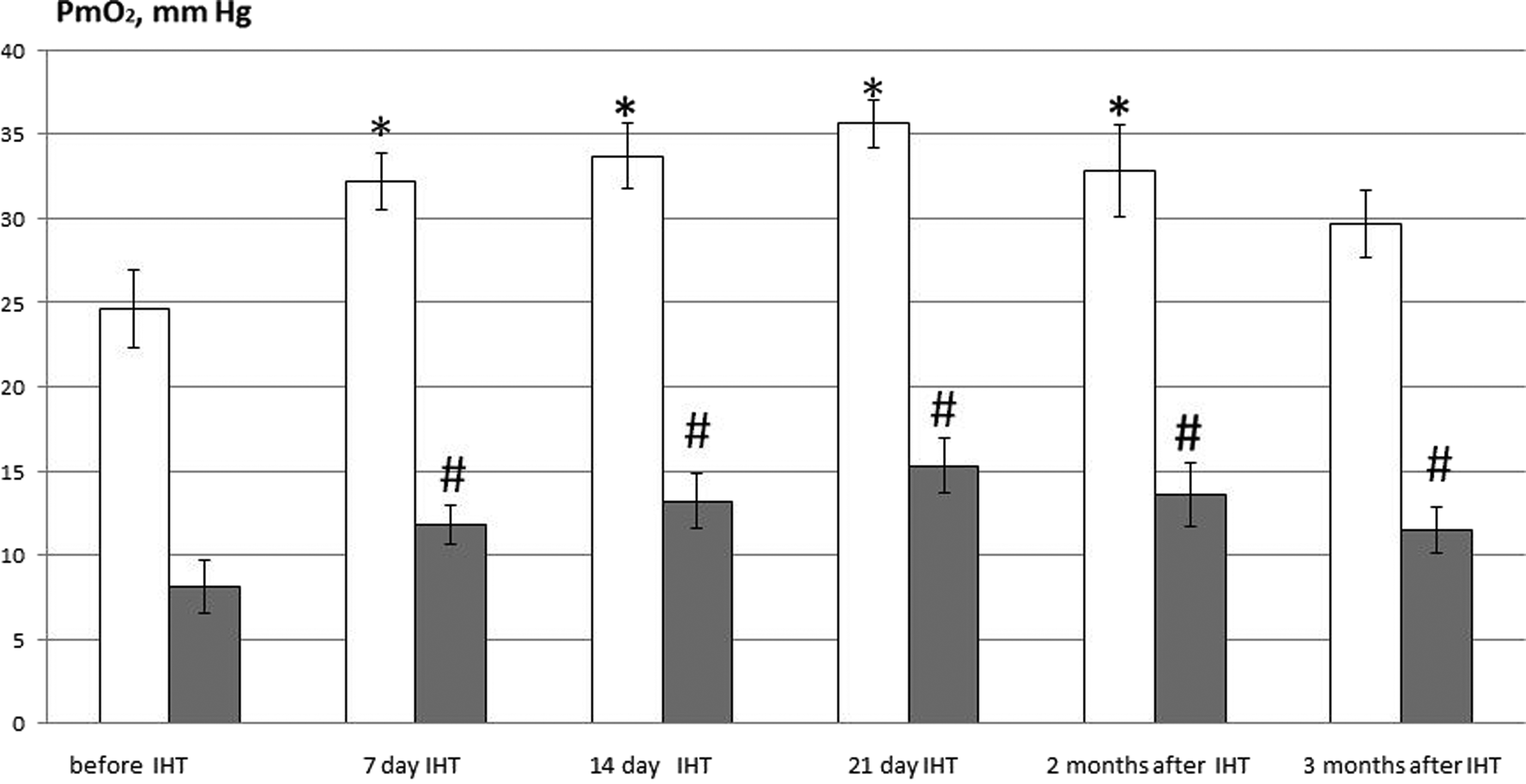
Oxygen tension (Pm
On this basis, we further investigated the liver and heart mitochondrial respiration during a 3-week course of IHT (Mode 1) and its consequences for 2 and 3 months. For comparison, we also tested IHT in Mode 2 during 21 days.
When using Mode 1, it was shown that adaptation to IHT caused the reorganization of mitochondria energy metabolism. Table 2 shows mitochondrial oxygen consumption data using FADH-(succinate) and NADH-dependent substrates (glutamate+malate). Seven days of IHT treatment (Gr.II-2) induced a decrease in liver mitochondrial O2 consumption (V3) at succinate oxidation and an increase in respiratory control ratio (V3/V4 ATP ) indicating the enhancement of respiration and phosphorylation coupling. Meanwhile, the effectiveness of O2 consumption (ADP/O ratio) did not change (Fig. 3). This means that the significance of succinate oxidation pathway (mitochondrial enzyme complex II, MEC II), which is effective at acute hypoxia impacts, becomes insignificant under IHT treatment. Mitochondria switch the energy production to a more effective NADH-dependent pathway (mitochondrial enzyme complex I, MEC I). This is well illustrated by mitochondrial oxygen consumption under the MEC I substrates oxidation (Table 2): an increase in V3/V4 ATP and V3 by 23% and 20%, respectively. ADP/O ratio increased by 13% (Fig. 3). The defined IHT effects for liver mitochondria were also registered after 14- day IHT (Gr. II-3), diminished when using IHT during 21 days (Gr. II-4), and mostly preserved in 2 (Gr. II-5) and 3 (Gr. II-6) months after IHT finished (Table 2, Fig. 3).
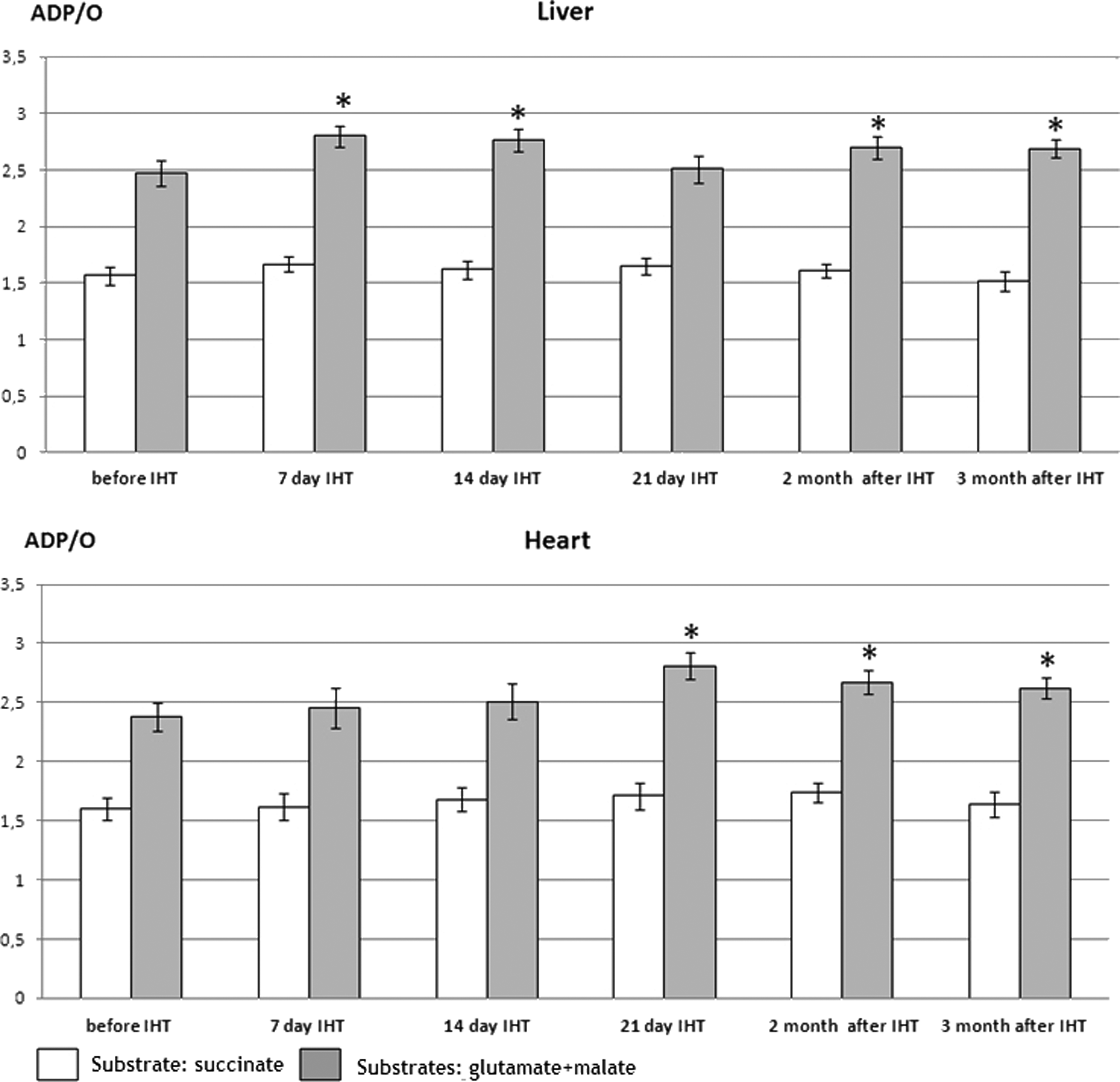
Effectiveness of O2 consumption by liver and heart mitochondria under IHT. White columns characterize ADP/O ratio when using the MEC II substrate succinate (in the presence of rotenone) for oxidation; gray columns, ADP/O ratio when using the MEC I substrates (glutamate+malate) for oxidation; *p<0.05, significantly different from initial value (before IHT).
Values are means±SD.
V3, oxygen consumption stimulated by ADP; V4 ATP , oxygen consumption after cessation of ADP phosphorylation; V3/V4 ATP , respiratory control ratio.
At the same time, the significant adaptive changes in the heart mitochondrial respiration developed later than in the liver, viz only after 3 weeks of training of training, showing a decrease in V3 by 11%, an increase in V3/V4 ATP by 20% when using MEC II substrate, and an increase in V3 and V3/V4 ATP by 16% and 26%, respectively, when using MEC I substrate (Gr. II-4). Beneficial effects continued during 2 and 3 months after IHT finishing.
When using Mode 2 (Table 3), the positive changes in mitochondrial respiration were also registered, but they were less pronounced. For example, after 21-day IHT the V3/V4 ATP for heart mitochondria increased by 18% (Gr.II-8) as opposed to an increase in this parameter by 26% under Mode 1 (Table 2). For liver mitochondria, the respiratory control ratio changed insignificantly.
Values are means±SD.
V3, oxygen consumption stimulated by ADP; V4 ATP , oxygen consumption after cessation of ADP phosphorylation; V3/V4 ATP , respiratory control ratio.
Discussion
In this study we compared for the first time the effects of five widespread modes of IHT on rat gastrocnemius muscle P
To determine the optimal mode of IHT in sports and military activities, many authors use such key parameters as increased ventilation, arterial oxygen saturation, and hemoglobin or hematocrit values, (Katayama et al, 2009; Millet et al., 2010; Musa, 2007; Wilber et al., 2007). In our experiments on animals, we took the most informative integral parameter of muscle oxygen supply—Pm
Animal investigations have also used a variety of exposure paradigms to study intermittent hypoxia. Foster et al. (2005) using two regimens—(1) 5 min of 12% O2/5 min of air, 1 h, 12 day, and (2) 30 min of 12% O2, 12 days—showed that both short and long hypoxic durations had similar effects on the ventilatory and cardiovascular response to acute progressive hypoxia. Similar results were obtained on humans (Ainslie et al., 2007) who compared short-term intermittent hypoxia (IH: 5-min hypoxia to 5-min normoxia, 90 min/day, 12 days) with continuous hypoxia (CH: ascending to 1560 m altitude for 12 days) and showed that IH and mild CH can equally enhance the HVR, which facilitates alterations in blood pressure and middle cerebral artery blood flow velocity.
But most authors consider that the shorter periods of hypoxic and normoxic exposures have more beneficial effects on an organism. Currently, there is practical necessity to decrease training time for achieving long-term adaptation, while maintaining its effectiveness. As Lukyanova (2005) suggested, short-term hypoxia does not cause tissue damage and permits changes of mitochondrial enzymes to occur during normoxic intervals.
Peng and Prabhakar (2004) exposed rats to either short-duration intermittent hypoxia (SDIH, 15 s of 5% O2 at 5 min intervals, 8 h day−1) or long-duration hypobaric intermittent hypoxia (LDIH, 4 h day−1, 0.4 atm for 10 days) and showed that exposure to SDIH enhanced carotid body sensory response to hypoxia, but exposure to LDIH did not. Satriotomo et al. (2012) believe that acute intermittent hypoxia (5-min episodes of 10.5% O2 with 5-min normoxic intervals) initiates plasticity in respiratory motor control. Hu et al. (2010), using rat model's hypoxic preconditioning, concluded that 15%–10% oxygen concentration range with short breaks could be regarded as a helpful effective area.
One of the key mechanisms of cell damage during hypoxia and reoxygenation is an excessive production of reactive oxygen and nitrogen species in mitochondria. These considerations may partly explain why the prolongation of reoxygenation periods in our studies from 5 to 15 minutes (Modes 2, 3, and 5) led to the deterioration of muscle oxygen supply. Probably, longer normoxia provokes greater free radicals production, preventing normal oxygen delivery to tissues. On the other hand, there is evidence that by using 30% O2 during recovery, adaptation to IHT could be achieved earlier and provide the upregulation of adaptive ROS signals compared to classical intermittent hypoxic training (Gonchar and Mankovska, 2012; Zhukova et al., 2005). Studies support the viewpoint that brief hypoxic generation of free radicals induces antioxidant enzyme protein synthesis that may be an important trigger for specific adaptations. Unfortunately, fundamental molecular studies on cell culture models (Chua et al., 2010; Kim et al, 2006; Mansfield et al., 2005), which provided important insights into mechanisms of HIF activation by hypoxia, can not explain exactly why the prolongation of reoxygenation periods during IHT treatment provoked less positive effects on the Pm
Additionally, reactive oxygen species (ROS) also function as secondary messengers in a variety of physiological processes increasing antioxidant defense. It was shown that ROS participate in the signaling pathways involved in the activation of multiple transcription factors such as HIF-1, NF-κβ, c-fos, c-Jun under IHT (Huang et al., 2005; Prabhakar and Semenza, 2012; Sasaki et al., 2001). In our experiments, the extension of reoxygenating periods both after moderate (12% O2) and severe (7% O2) hypoxia resulted in the deterioration of muscle oxygen supply. In addition, a decrease in oxygen content per se from 12% to 7% also led to worsening of tissue P
Our investigations also have shown that adaptation to IHT in Mode I caused reorganization of liver and myocardium mitochondrial energy metabolism favoring NADH-dependent oxidation. Since mitochondrial superoxide production is inversely related to MEC I activity, the activation of the latter by IHT could limit the oxidative stress development under acute hypoxia (Koopman et al., 2005; Verkaart et al., 2005). So, increasing oxidation of NAD-generated substrates may preserve the mitochondrial oxygen consumption capacity during stress conditions. One can also assume that adaptation to IHT decreased the vulnerability of heart and liver mitochondria to ROS as demonstrated by elevated respiratory control ratio, ADP/O ration, and maximal rate of oxidative phosphorylation.
When using Mode 2, positive changes in mitochondrial respiration were also registered, but they were less pronounced.
Our findings are in agreement with previous reports demonstrating the activation of mitochondrial respiration by IHT (Kurhaliuk et al., 2002; 2013; Prabhakar and Kumar, 2004;). As Lukyanova et al. (2009) suggest, the suppressed function of MEC I and the alternative activation of MEC II comprise a signaling regulatory mechanism that contributes to the formation of tissue-specific and general resistance of the body to different types of hypoxia.
Recent studies demonstrated the mechanisms by which intermittent hypoxia reversibly inactivates MEC I. The primordial function of HIF-1 appears to involve finding the optimal balance between oxidative and glycolytic metabolism for any given cell as a function of the local O2 concentration (Prabhakar and Semenza, 2012). Based on cell culture experiments with very short (30 sec) intermittent hypoxic exposures, Khan et al. (2011) identified NADPH oxidases and mitochondrial electron transport chain at MEC I as major cellular ROS sources mediating systemic and cellular responses to intermittent hypoxia and demonstrated a functional cross-talk between NADPH oxidases and MEC I activity.
According to our results, we consider that the Mode 1 of IHT (5 min 12% O2 with 5-min normoxic intervals) is more effective and less dangerous for the studied organs and tissues and may be recommended for sports, as well as for prophylaxis and treatment of cardiac and lung pathology. However, we must take into account that all these beneficial results were obtained on a rat model. In actual human practice (Musa, 2007), the regimen of IHT (the degree of hypoxia, exposure duration, and number of sessions) could be related to the mission requirements, such as the operational target altitude, the risk of developing acute mountain sickness, or the anticipated physical activity levels.
Conclusion
Our experimental data indicate that among five tested modes of IHT, the optimal hypoxic dose for muscle oxygen supply is 5-min breathing with 12% O2 gas mixture and 5-min breaks (Mode 1), 5–6 times a day during 2 or 3 weeks. Under such mode, Pm
Using Mode 1, it was shown that adaptation to IHT caused the substrate-dependent reorganization of liver and heart mitochondrial energy metabolism favoring NADH-dependent oxidation and improving the efficiency of oxidative phosphorylation. Mitochondrial adaptation developed after 7 days of IHT in liver tissue, but after 21 days in myocardium and was preserved for 3 months after IHT termination. When using Mode 2 (15-min 12% O2 with 15-min breaks), positive changes in mitochondrial respiration were also registered, but were less pronounced.
Footnotes
Author Disclosure Statement
No competing financial interests exist.