
Abstract
Abstract
Espinoza, Jose R., Giancarlo Alvarez, Fabiola León-Velarde, Hugo F. Ju Preciado, Jose-Luis Macarlupu, Maria Rivera-Ch, Jorge Rodriguez, Judith Favier, Anne-Paule Gimenez-Roqueplo, and Jean-Paul Richalet. Vascular endothelial growth factor-A is associated with chronic mountain sickness in Andean population. High Alt Med Biol. 15:146–154, 2014.—A study of chronic mountain sickness (CMS) with a candidate gene—vascular endothelial growth factor A (VEGFA)—was carried out in a Peruvian population living at high altitude in Cerro de Pasco (4380 m). The study was performed by genotyping of 11 tag SNPs encompassing 2.2 kb of region of VEGFA gene in patients with a diagnosis of CMS (n=131; 49.1±12.7 years old) and unrelated healthy controls (n=84; 47.2±13.4 years old). The VEGFA tag SNP rs3025033 was found associated with CMS (p<0.05), individuals with AG genotype have 2.5 more risk of CMS compared to those with GG genotype (p<0.02; OR, 2.54; 95% CI: 1.10–5.88). Pairwise Fst and Nei's distance indicate genetic differentiation between Cerro de Pasco population and HapMap3 population (Fst>0.36, p<0.01), suggesting selection is operating on the VEGF gene. Our results suggest that VEGFA is associated with CMS in long-term residents at high altitude in the Peruvian Andes.
Introduction
C
Vascular endothelial growth factor (VEGF) is one of the most potent angiogenic factors (Leung et al., 1989). VEGFA gene has eight exons that produce several VEGF isoforms by alternative splicing (Tischer et al., 1991). It stimulates vascular endothelial cells and other tissues (Ferrara et al., 2003), and appears as the main mediator of hypoxia-induced-neovascularization (Robinson and Stringer, 2001), vascular permeability, and other functions (Senger, 2010). Both VEGF and erythropoietin derive from the hypoxia-inducible factor 1 (HIF1) regulatory pathway and many interactions have been described between the angiogenic and erythropoietic pathways. They both exert neurotrophic and neuroprotective effects in the central nervous system (Dale et al., 2013). In patients with von Hippel Lindau disease, the administration of anti-VEGF drug induced a polycythemia (Richard et al. 2002), suggesting the hypothesis that, in patients with CMS, an increase in VEGF production might be a compensatory mechanism to polycythemia.
In addition to its angiogenic function, VEGF may also activate pathways associated with NO synthesis and thus induce vasodilation and improve blood supply to cardiac cells (Schoch et al., 2002). Acute hypoxia or Epo-deficiency result in an overexpression of HIF-1α, VEGF, Epo, and Epo receptor in the heart and the brain of mice, while chronic hypoxia depresses the HIF-1α/VEGF pathway in Epo-deficient mice (El Hasnaoui-Saadani et al., 2009; 2013).
Angiogenesis and vasculogenesis are part of the physiological response to hypoxia and therefore might have a role in adaptation to high altitude in long-term residents, especially in subjects with excessive erythropoiesis in order to compensate a plausible alteration in microcirculation (Appenzeller et al., 2003; Ge et al., 2011; Buroker et al., 2012).
Therefore, a change in the expression of a gene related to VEGF might be considered as a marker of adaptation to the disease, and not to a contributor of polycythemia. The present work was undertaken to assess the contribution of the candidate gene VEGFA in the genetics of CMS by a controlled study.
Materials and Methods
Cases and controls
The study population was composed of 215 residents of Cerro de Pasco (4380 m, Peru), studied from February 2010 to May 2011: 131 were cases affected by CMS (49.1±12.7 years old) and 84 were healthy controls (47.2±13.4 years old). Volunteers were clinically assessed by a physician to establish their condition in reference to CMS by applying the Qinghai score that included Hb ≥21g/dL cut-off and clinical symptoms (Leon–Velarde et al., 2005). Only male individuals with residence longer than 6 months in Cerro de Pasco were recruited in the study. Individuals were not enrolled in the study if they have any other chronic disease or smoking habit (≥1 cigarette per day). This research was conducted at the Molecular Biotechnology Unit of the Laboratories of Research and Development from Universidad Peruana Cayetano Heredia (Peru) and in the Paris-Cardiovascular Research Center at the Georges-Pompidou European Hospital (INSERM, UMR970) in Paris, France.
The study was approved by the ethics committee of Universidad Peruana Cayetano Heredia. All participants were enrolled in the study after signing an informed consent.
DNA extraction
A blood sample was drawn from all participants. Frozen blood tubes were delivered to the laboratory. Leukocytes DNA was extracted by salting out procedure (Sambrook et al., 1989). DNA was quantified and qualified with NanoDrop™ 2000 (260/280 ≥2.0; 260/230 ≥1.8) and stored at −20° C until use.
Selection of tag SNPs
Informative SNPs (tag SNPs) were selected using a multi-population algorithm applied to twelve HapMap3 populations (http://hapmap.ncbi.nlm.nih.gov/) and the following parameters: combined samples with common variants, cut-off value for allele frequency was 1%, no monomorphic sites were allowed, and minimum value threshold r2=0.8.
PCR amplification of tagSNPs
Seven pair of primers (available on request) were designed to amplify by PCR the VEGFA region containing the selected tag SNPs with Primer 3 online program hosted by SourceForge. The default parameters were set to GC content, to minimize mispriming and complementary or self-complementarity annealing. PCR amplification was carried out using Taq polymerase Mix Red’y’Gold (Eurogentec, Seraing, Belgium), primers, 200 μm dNTP, 5 mM MgCl2 1, 2X buffer and red loading dye buffer. PCR Reaction Mastercycler nexus thermocycler (Eppendorf AG, Hamburg, Germany) was used and was calibrated on a temperature gradient between 50°C and 60°C. Finally melting temperatures selected were 60°C, 58°C, 60°C, 56°C, and 54°C for each of the seven pairs of primers.
Genotyping
We amplified the VEGFA gene fragments with each corresponding pair of primers from 215 DNA samples. PCR conditions were: 95°C for 10 min; 24 cycles composed by 45 sec at 95°C, followed by 45 sec with the annealing temperature according to the set of primers and then 45 sec at 72°C; a final extension at 72°C for 7 min, and a storage at 4°C. Samples were run on an agarose gel for verification and purified with Exonuclease I and Antarctic Phosphatase (New England Biolabs, Inc). Finally the purified PCR products were carried out through a sequencing reaction by using BigDye Terminator with forward primers only. The samples were run in a 3730xl DNA Analyzer (Applied Biosystems,) after a step of purification of the PCR products with silica powder. Sequences were analyzed with the software Sequencher version 5.1 (Gene Codes Corporation, Ann Arbor, MI, USA).
Statistical analysis
Student t-test was used to compare physiological variables between cases and controls. Hardy-Weinberg equilibrium and linkage disequilibrium were estimated by using Genepop 4.1. All statistical analyses were conducted using the statistical software STATA: Release 11 (StataCorp LP, College Station, TX). Genotype and allele frequencies of each SNP were compared in cases and controls with a Pearson's chi-squared test (χ2). For association analyses, a value of p<0.05 was considered significant, for comparison of genotype frequencies with HapMap3 a value of p≤0.001 was used for statistical significance. Odds ratio (OR) was calculated in case of significance. Haploview program was used for haplotype association analysis and to define haplotype blocks. Data analysis was done with Epi Info 2002 program (http://wwwn.cdc.gov/epiinfo/). Haplotypes frequency and linkage disequilibrium were determined by EH program (v. 1.1; Jurg Ott, Rockefeller University, New York). Linkage disequilibrium (LD) between pairs of markers in VEGFA was estimated by using Lewontin's D’ parameter (Zhao, 2004).
Results
The CMS cases (n=131) and heathly controls (n=84) were significantly different for hematocrit (p<0.001), O2 saturation (p<0.001), heart rate (p<0.01), body max index (p<0.001), and weight (p<0.05). No significant difference was observed for age, blood pressure, and height (Table 1).
Mean±SD. p: significance with Student's t-test between cases and controls.
Eleven tag SNPs were genotyped in cases and controls. Sporadic sequencing failures explain slight variations of the total number of genotype and allele frequencies (Table 2). None of these SNPs deviated from Hardy-Weinberg equilibrium and no significant difference between the expected and observed heterozygosity was observed (Table 3). A significant association was found with the VEGFA tag SNP rs3025033 (p=0.044) as shown in Table 4. Individuals with AG genotype at this locus have a 2.5 greater risk of CMS compared to those with GG genotype (p=0.017; OR=2.5; CI 95%, 2.29–2.95)
There was no difference between the expected and observed heterozygosities. The Hardy-Weinberg law was tested by a χ2 test with one degree of freedom and although the markers rs833068, rs833069, rs3024997, and rs3025000 showed a value of p<0.05, these were found to be not significant after Yates and Bonferroni corrections. Finally, PIC values for markers rs25648, rs10434, and rs3025035 were smaller than 0.25, which indicates that they are poor informative markers but this could be due to the shortage of heterozygotes for these markers.
p, significance with Pearson Test.
A significant association was found for marker rs3025033. Given this statistical significance, we proceeded to perform an analysis of odds ratio: subjects with the AG genotype have a 2.5 greater risk of developing the disease compared with those with the GG genotype (p=0.017) with confidence intervals at 95% from 2.29 to 2.95.
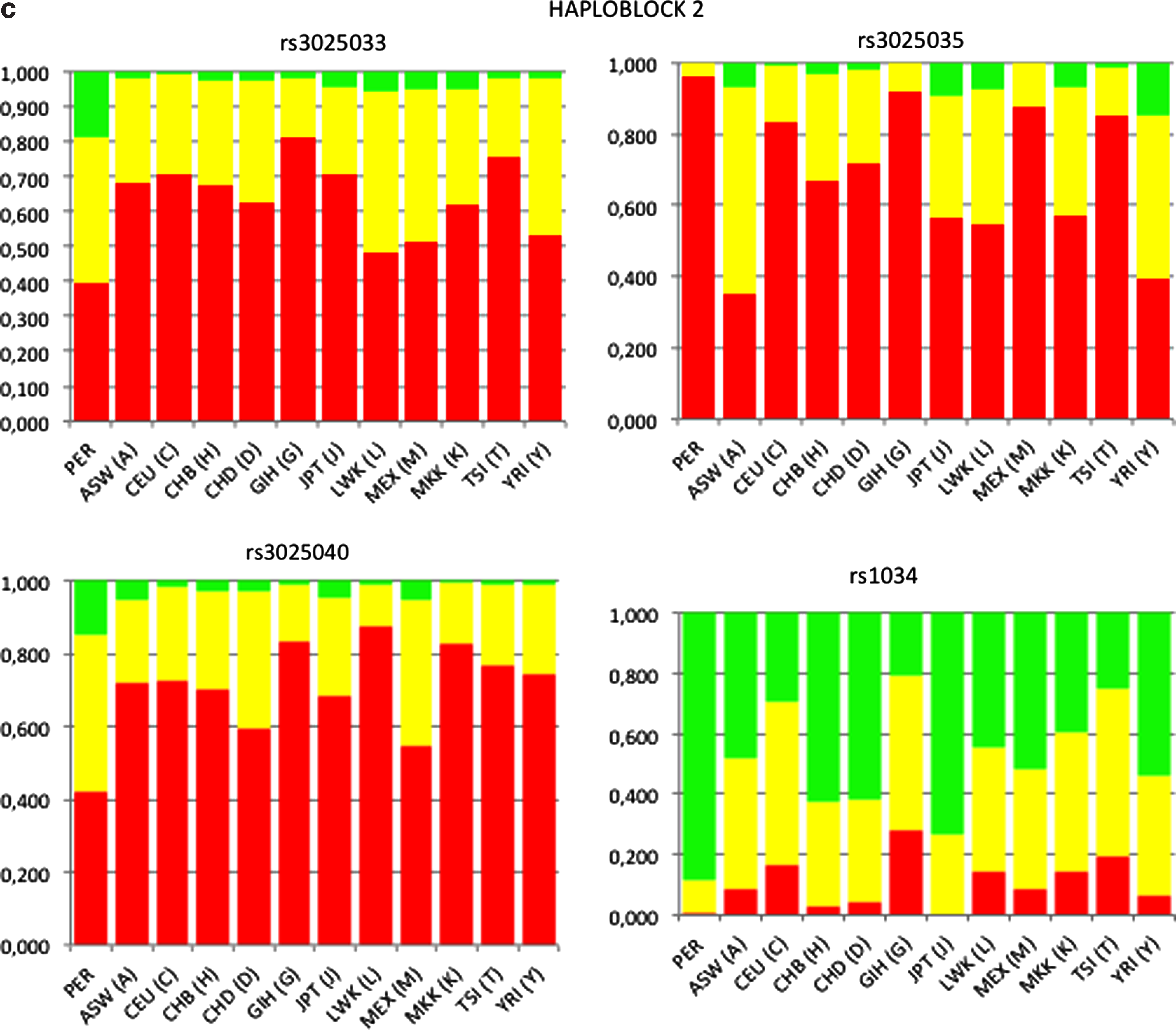
VEGFA is organized in two haploblocks based on LD values between the tag SNPs. The haploblock 1 (∼8 kb, LD ≥0.89) extended over rs25648, rs833068, rs833069, rs833070, rs3024997, rs3025000, rs3025010 and haploblock 2 (∼2 kb; LD ≥0.95) included rs3025033, rs3025035, rs3025040, rs1034 (Fig. 1).Haplotypes did not show any significant association with CMS (Table 5).

Haplotype blocks in the VEGF-A locus in Peruvian Andeans. Two haplotype blocks based on LD values separated by recombination between rs3025010 and rs3025033 SNP markers.
There was no significantly associated haploblock. The haplotypes were tested for a frequency greater than 1%. The haplotypes of the block 1 TGTTGCC and CGTTGCC are over-represented in comparison to the controls, while haplotypes CACCATT, CGTTGCT, and CGTCGCT are under-represented. On the other hand, the block 2 haplotypes that are over-represented are ACCG and ATCG, while haplotypes GCTG, ACCA, and CGCG are under-represented in comparison to healthy controls. The haplotypes with higher frequency are CACCATT with 35.5% for block 1 and ACCG with 52.6% for block 2.
A pairwise comparison of the genotype and allele frequencies of the VEGFA tag SNPs in residents of Cerro de Pasco, including CMS and controls, was performed with 11 HapMap3 reference populations (Fig. 2).A significant difference in genotype and allele frequencies was observed in SNPs of haploblock 2 of the Cerro de Pasco residents compared all other populations (p<0.001) but the difference was not as extended as in haploblock 1 (Fig. 2).
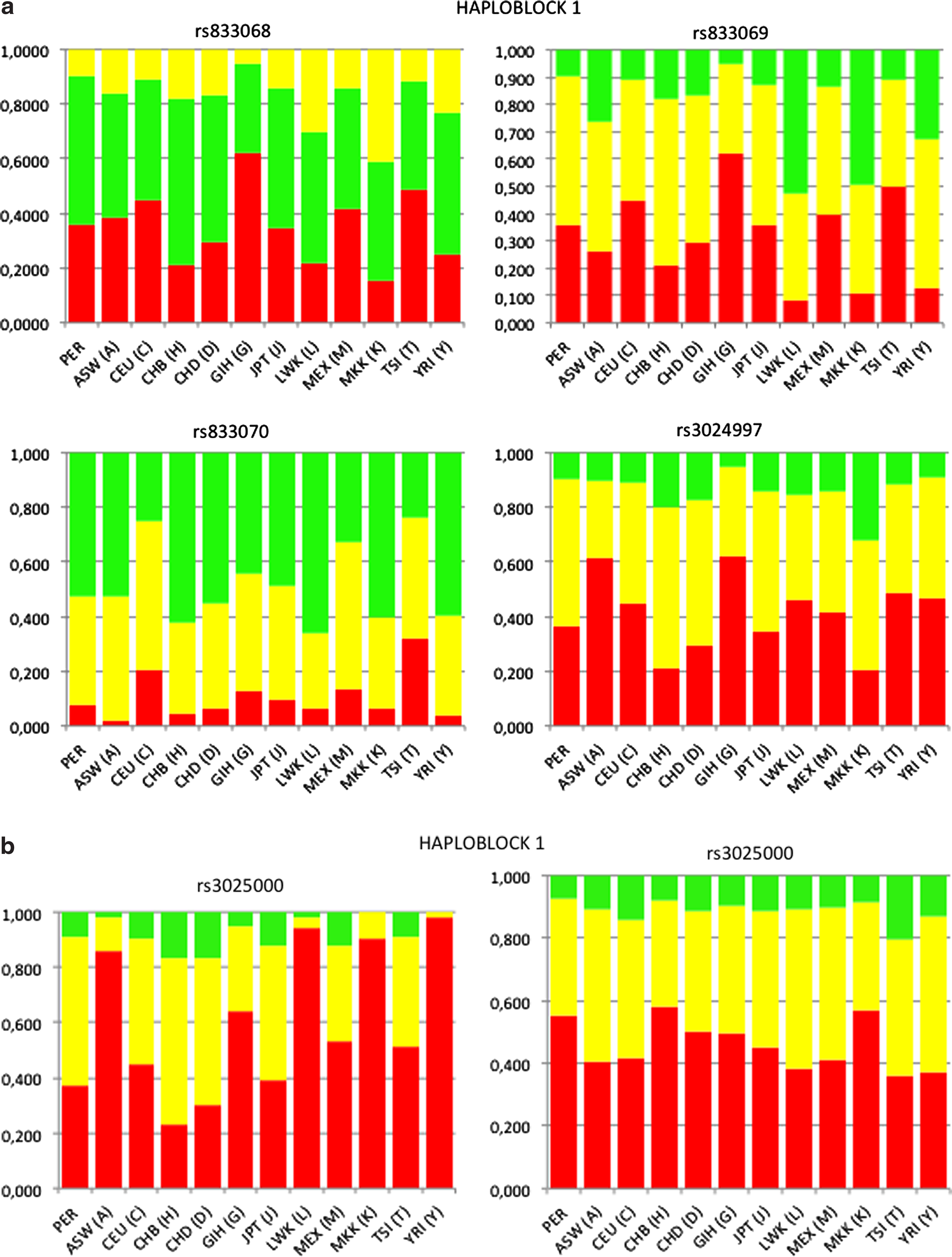
Genotype frequency of tagSNPs in haploblocks 1 and 2 of VEGFA locus in Peruvian and HapMap3 populations.
Pairwise Fst was calculated between CMS and non-CMS Peruvian and HapMap3 populations with tag SNPs from haploblocks 1 and 2 of the VEGFA gene (Table 6). The pairwise Fst for haplogroup 1 (0.3631–0.4238; p<0.01) and haplogroup 2 (0.105–0.1595; p<0.01) indicated moderate but significant genetic differentiation between Peruvian high altitude residents and HapMap3 populations, but no differentiation was observed between CMS and non-CMS (Table 6). Nei's genetic distance showed the divergence between high altitude CMS and non-CMS Peruvians from HapMap3 population. Altogether, these data suggest a genetic differentiation of Cerro de Pasco residents based on VEGFA tag SNPs (Fig. 3).
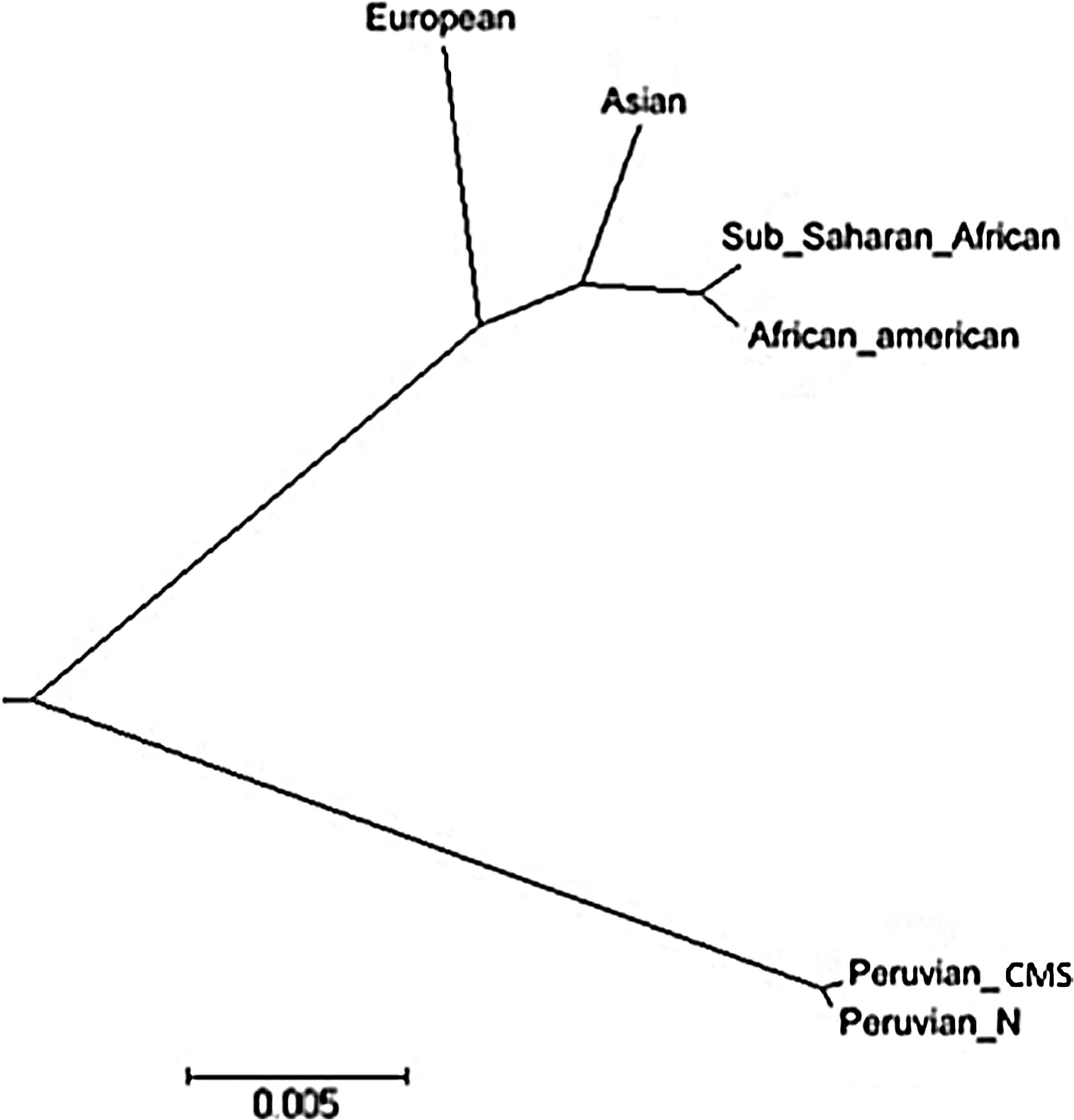
Nei's genetic distance of CMS, non-CMS Peruvians, and HapMap3 populations.
p<0.01
Discussion
In a study involving 215 subjects living at 4380 m, we found that the VEGFA tag SNP rs3025033 was associated with CMS, individuals with AG genotype having 2.5 more risk of CMS compared to those with GG genotype. The genetic basis of Monge's disease has been difficult to tackle due to the late clinical manifestation of the disease, as well as to the lack of animal models (Sime et al., 1975), making it difficult to ascertain the mode of inheritance of the disease and to map causative genes by linkage analysis. The first approaches were done by gene candidate studies, but until now no gene has been clearly involved in the development of Monge's disease (Appenzeller et al., 2003; Mejia et al., 2005; Leon-Velarde and Mejia, 2008, Xing et al., 2008). Recent studies have proposed some genetic characteristics of high altitude natives (Tibetan) to be advantageous for life at high altitude, but no attempt was made to compare them to subjects suffering from high altitude-induced diseases (Beall et al., 2010; Simonson et al., 2010; Yi et al., 2010). The recent study by Zhou and co-workers gives an interesting insight in the comprehension of the mechanisms involving the regulation of erythropoiesis in CMS (Zhou et al., 2013). However, the very small number of patients included (10) necessitates replication studies.
We cannot formally exclude that the association we found between VEGFA tag SNP rs3025033 and CMS was by chance, since we did not perform a Bonferroni correction for multiple comparison. However, the lower limit of the OR was very good and far from 1 (2.29), suggesting that the risk of predicting a spurious association is low. Anyway, this study aims to be replicated in similar populations living at high altitude in the Altiplano and suffering from CMS.
High altitude poses a selection pressure to human populations living in this environment (Stortz, 2010). CMS appears as a lack/loss of adaption of humans to the stress of high altitude, showing an exacerbated production of circulating red blood cells, high levels of hemoglobin and other physiological traits (Leon Velarde et al., 2005). Pulmonary hypertension might develop, due to hypoxic pulmonary vasoconstriction and remodeling, as well as high-viscosity induced increase in pulmonary vascular resistance (Richalet et al., 2008; Pichon et al., 2012; Naeije and Vanderpool, 2013). Whole genome scanning using SNPs chips has suggested the role of several genes in adaption to high altitude in Andeans, such as EGLN1 (PHD2), NOS2A, PRKAA1 (Bigham et al., 2009; 2010), in which VEGF and nine other genes regulated by HIF appear as candidates for positive selection by two statistical tests (Bigham et al., 2009). Therefore, these genes appeared as candidates for association with CMS.
In the present study, we did not measure circulating VEGF in order to correlate the observed SNP polymorphism with changes in protein concentration or turnover. However, the circulating VEGF level has been shown to increase in individuals who ascend to high altitude (Walter et al., 2001, Tissot van Patot et al., 2004) and in patients with CMS (Ge et al., 2011). In both cases, VEGF levels decrease when individuals are moved to sea level. Whether this response influences the outcome of the disease in long residents is still unknown. However, VEGF expression in peripheral blood mononuclear cells of CMS patients appears different compared to high altitude natives living at high altitude or sea level (Appenzeller et al., 2003). Interestingly, the risk of developing acute mountain sickness has also been associated with a polymorphism of VEGFA (Ding et al., 2012).
Our results suggest that VEGFA is associated with CMS in Peruvians, as also reported with CMS in Tibetans: in a study performed in Tibet, CMS (n=48) was found to be significantly associated with single nucleotide polymorphisms involving VEGFA (rs1570360, rs28357093, rs13207351) compared to Tibetan unaffected subjects (n=31) (Buroker et al., 2012). Moreover, in the same study, these nucleotide changes were associated with physiological variables such as arterial O2 saturation and red blood cell count. Recently, the same group confirmed their previous results, showing that SNPs in the promoter region of VEGFA are associated with the development of mountain sickness (Buroker et al., 2013).
Even though the genetics of high altitude adaptation in Tibetans is probably different from Andeans (Beall, 2000; Moore, 2001; Wu and Keyser, 2006), VEGFA might play a role in the development of CMS in both populations. The allele and genotype frequencies in SNPs of the haploblock 2 are different from the HapMap3 populations of the International HapMap 3 Consortium (2010). A significant genetic differentiation between Cerro de Pasco population and HapMap3 populations (Fst >0.36, p<0.01) suggests that high altitude poses a selection pressure on VEGF locus.
Footnotes
Acknowledgments
This work was partially funded by the Franco-Peruvian Red Raul Porras Barrenechea, by the Agence Nationale de la Recherche (ANR 08 GENOPAT 029 MitOxy) and by the Pulmonary Vascular Research Institute.
Author Disclosure Statement
None of the authors declare any conflict of interest.