
Abstract
Abstract
Ainslie, Philip N., and Andrew W. Subudhi. Invited Review: Cerebral blood flow at high altitude. High Alt Med Biol. 15:133–140, 2014.—This brief review traces the last 50 years of research related to cerebral blood flow (CBF) in humans exposed to high altitude. The increase in CBF within the first 12 hours at high altitude and its return to near sea level values after 3–5 days of acclimatization was first documented with use of the Kety-Schmidt technique in 1964. The degree of change in CBF at high altitude is influenced by many variables, including arterial oxygen and carbon dioxide tensions, oxygen content, cerebral spinal fluid pH, and hematocrit, but can be collectively summarized in terms of the relative strengths of four key integrated reflexes: 1) hypoxic cerebral vasodilatation; 2) hypocapnic cerebral vasoconstriction; 3) hypoxic ventilatory response; and 4) hypercapnic ventilatory response. Understanding the mechanisms underlying these reflexes and their interactions with one another is critical to advance our understanding of global and regional CBF regulation. Whether high altitude populations exhibit cerebrovascular adaptations to chronic levels of hypoxia or if changes in CBF are related to the development of acute mountain sickness are currently unknown; yet overall, the integrated CBF response to high altitude appears to be sufficient to meet the brain's large and consistent demand for oxygen. This short review is organized as follows: An historical overview of the earliest CBF measurements collected at high altitude introduces a summary of reported CBF changes at altitude over the last 50 years in both lowlanders and high-altitude natives. The most tenable candidate mechanism(s) regulating CBF at altitude are summarized with a focus on available data in humans, and a role for these mechanisms in the pathophysiology of AMS is considered. Finally, suggestions for future directions are provided.
Introduction
R
Historical Overview—The First Measurement of CBF at Altitude
Angelo Mosso (1846–1910) is undoubtedly the forefather of high altitude cerebrovascular physiology. In order to pursue his principal curiosity of the physiological effects of hypobaria, Mosso built barometric chambers and was reported to expose himself pressures as low as 192 mmHg (equivalent to >10,000 m). He was also responsible for the building of the Capanna Margherita laboratory on Monta Rosa at 4559 m. In both settings, Mosso utilized his hydrosphygmomanometer to measure changes in “brain pulsations” in patients that had suffered removal of skull sections, due to illness or trauma (Mosso, 1880). Indicative of changes in CBF, these recordings preceded the next estimates of CBF in humans by some 50 years.
At sea level, Kety and Schmidt (Kety, 1945) were the first to quantify human CBF using an inert tracer (nitrous oxide, N2O) combined with arterial and jugular venous sampling. This method for the measurement of global CBF is based on the Fick principle, whereby the integrated difference of multiple arterial and venous blood samples during the first 10 or more minutes after the sudden introduction into the lung of a soluble gas tracer is inversely proportional to cerebral blood flow (Kety and Schmidt, 1948a). In 1948, they showed that breathing 10% oxygen increased CBF by 35% (Kety and Schmidt, 1948b); however, it was not until 1964 that the first measurements of CBF were made in humans at high altitude (Severinghaus et al., 1966). The motivation for these high-altitude experiments was stimulated, in part, from the earlier discovery of the brain's ventral medullary cerebrospinal fluid (CSF) pH sensors in animals (Mitchell et al., 1963). Following the location of these central chemoreceptors, Severinghaus and colleagues examined in humans the role of CSF pH and bicarbonate in acclimatization to high altitude (3810 m) at the White Mountain (California, USA) laboratories (Severinghaus et al., 1963). A year later, at the same location, John Severinghaus performed his seminal study of CBF at high altitude. He was joined by Tom Hornbein—shortly after his first ascent of Everest by the West Ridge—who was part of the research team and also volunteered for the study (Fig. 1a). The results (Fig. 1b) showed clear time-dependent changes in CBF during acclimatization to high altitude (HA).
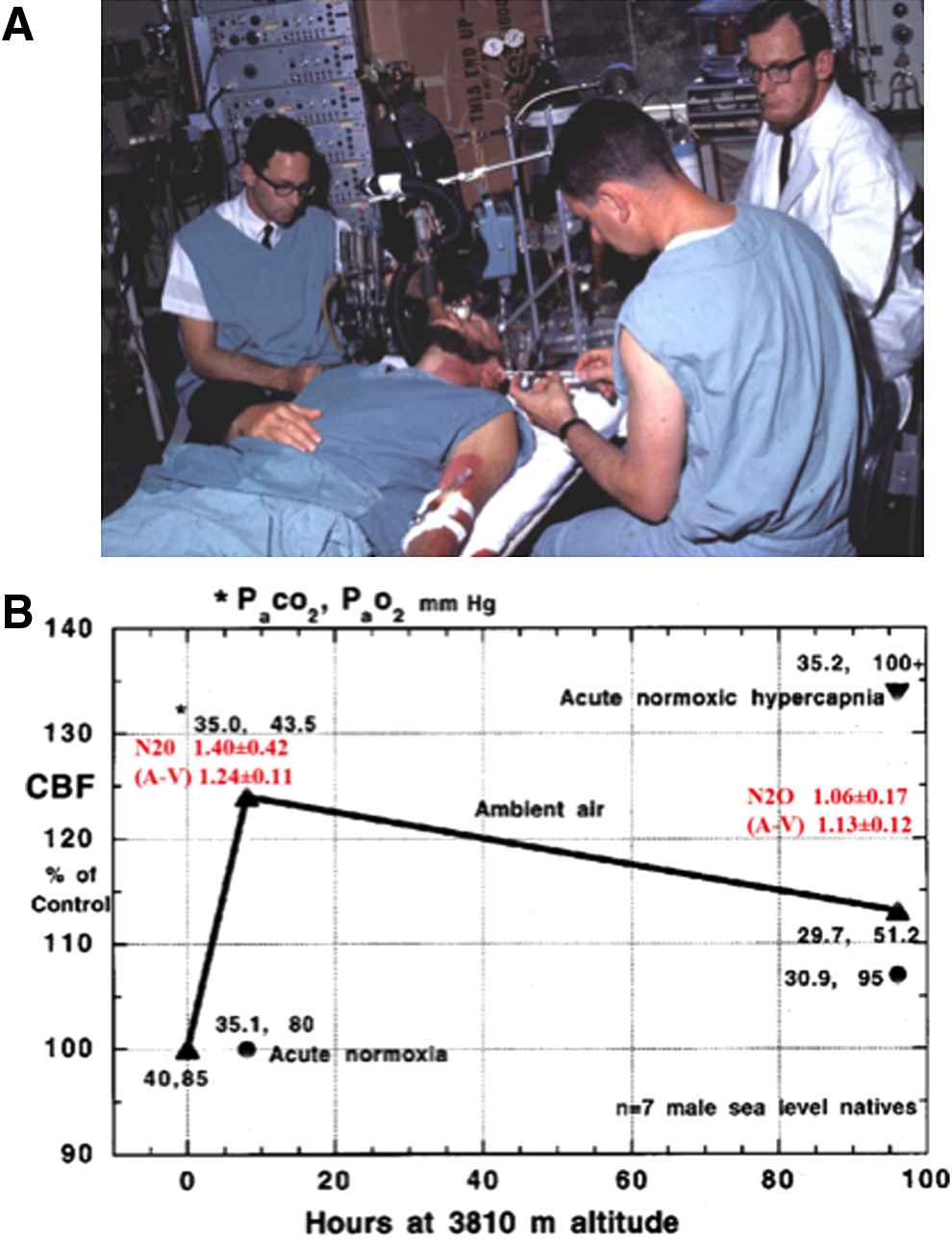
CBF in Lowlanders at High Altitude during Acclimatization
Severinghaus et al. (1963) first reported a ∼24% CBF increase within the first 6–12 hours of arrival at 3810 m before decreasing to 13% above sea level values by day 3–5; these data have been largely confirmed in several later studies (Huang et al., 1987; Jensen et al., 1990; Baumgartner et al., 1994; Lucas et al., 2011; Subudhi et al., 2013; Willie et al., 2013) conducted using a range of different methods to obtain estimates of CBF over extended periods at high altitude (Fig. 2). For example, Jensen et al. (1990) used radio-labeled xenon in 12 individuals who ascended from 150 m to 3475 m over 1–2 hours, and found a similar increase in CBF of 24%. Ascent from 3200 m to between 4785 and 5430 m caused a further increase in CBF to 53% above the readings at sea level. Four other studies measured CBF following ascent to >5000 m and showed a 40%–70% increase (Jensen et al., 1990; Lucas et al., 2011; Subudhi et al., 2013; Willie et al., 2013). The apparent dose dependency in the CBF response to high altitude has been confirmed in two recent studies (Wilson et al., 2011; Willie et al., 2013), yet it is unclear if the rate of ascent affects the CBF response. One study showed a peak CBF increase of 60% following slow (8 day ascent with Diamox supplementation to speed up acclimatization) ascent to 5050 m (Willie et al., 2013), while rapid ascent to 5260 m (on oxygen), resulted in a peak CBF increase of 70% (Subudhi et al., 2013). Regardless, all studies have shown that the initial increase in CBF dissipates with acclimatization. In essence, despite marked variation in methods to estimate CBF, ascent rate, and elevation, studies collectively show that CBF reaches a peak within 2–3 days at HA and then returns towards sea level values within 1–3 weeks (see Fig. 2). What is noteworthy from Figure 2 is that the increase in CBF is closely matched to the reduction in arterial oxygen content (Ca
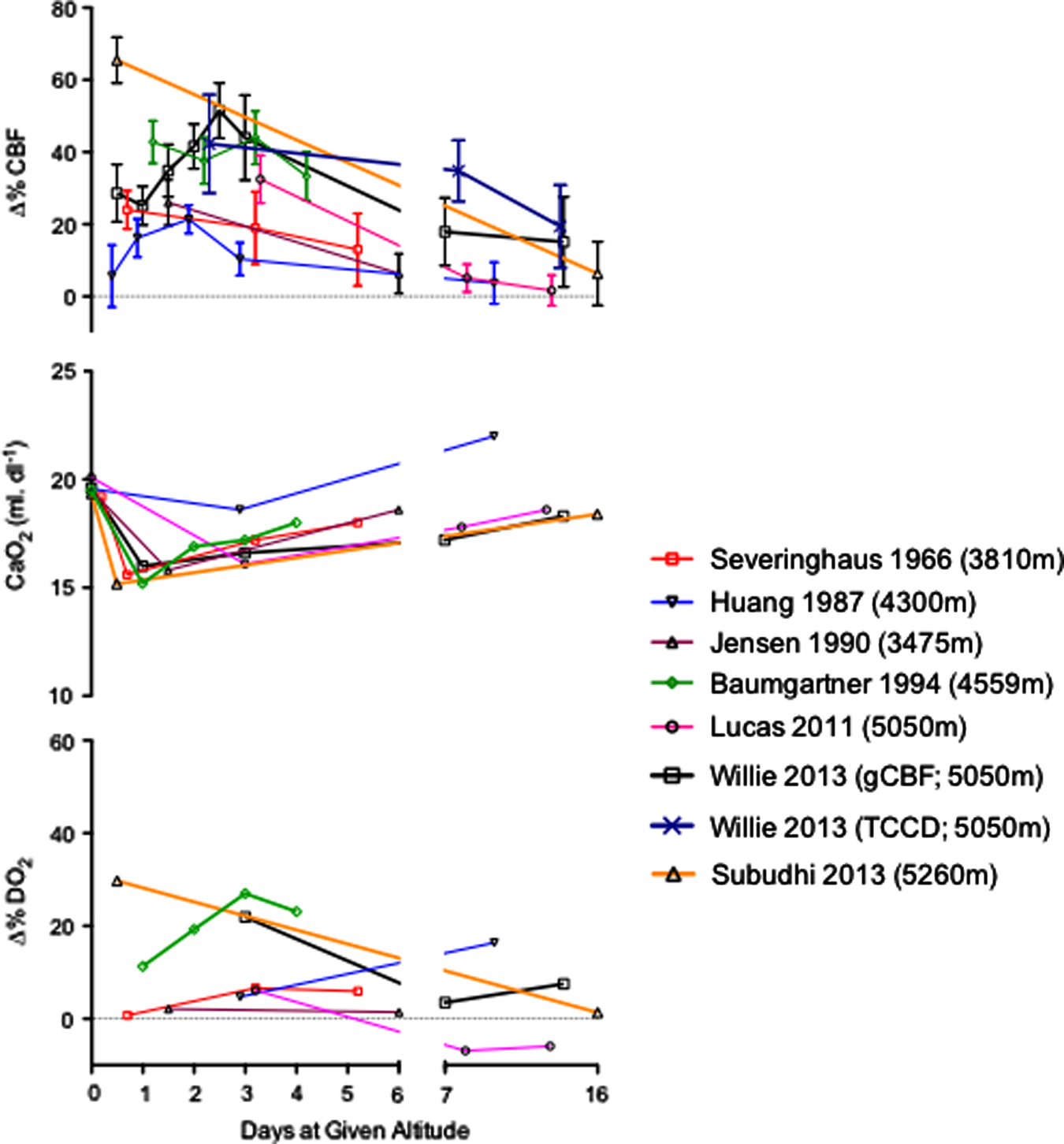
Percent changes in cerebral blood flow (Δ%CBF, graph A), arterial oxygen content (Ca
CBF in Natives to High Altitude
Native Tibetan (or Himalayan) and Andean populations arrived approximately 25,000 and 11,000 years ago, suggesting that these populations either carried traits that allowed them to thrive at high altitude or were able to adapt to the environment. The physiological and genetic traits associated with native high-altitude populations have been elegantly reviewed (Beall, 2007; Erzurum et al., 2007; Frisancho, 2013). As such, this topic is briefly summarized here with the focus on CBF at altitude in context of Andean and Tibetan high-altitude residents.
Andean high-altitude residents
In general, native Andeans have lower CBF values compared to sea level natives. The first evidence suggesting lower flow was reported in 8 Peruvian natives living at 4300 m altitude in Cerro de Pasco (Milledge and Sørensen, 1972). The authors found the mean arterial–venous oxygen content difference across the brain was 7.9±1 vol%, about 20% higher than the published sea level mean of 6.5 vol%. They suggested that CBF probably was proportionately about 20% below sea level normal values, assuming that brain metabolic rate was normal, and postulated that the mechanism might be high blood viscosity given the high hematocrit (58±6%) in these subjects. However, since the cerebral metabolic rate for oxygen (CMRO2) is constant even in severe hypoxia (Kety and Schmidt 1948b; Ainslie et al. 2013), the inverse linear relationship between CBF and arterial–venous oxygen content differences could also explain the reduction in CBF, as less flow would be needed to match the oxygen demand of the brain when arterial content is elevated. A similar study (Sørensen et al., 1974), using arterio-venous differences combined (in a subgroup) with a modified version of Kety–Schmidt method (krypton instead of N2O,) conducted in high-altitude residents in La Paz in Bolivia at 3800 m, also reported a 15%–20% reduction in CBF (with a reported average hematocrit of 50%) compared to a sea level control group.
More recently, a number of studies using transcranial Doppler (TCD) ultrasound have been performed in high-altitude residents in Cerro de Pasco at 4330 m (Appenzeller et al., 2004; Claydon et al., 2005; Appenzeller et al., 2006). Although sea level controls were not included, the findings reveal about 20% lower middle cerebral arterial blood flow velocity (MCAv) than would be expected in age-matched controls at sea level (Willie et al., 2011). The major factor underpinning the ∼20% reduction in CBF of high-altitude residents is the reported elevation in hematocrit and consequently increased Ca
Tibetan high-altitude residents
Unfortunately, information on CBF in Tibetan high-altitude residents is sparse and largely based on blood velocity indices of CBF in the internal carotid artery [(ICA) Huang et al., 1992 (3658 m)] or MCA [(Jansen et al., 1999; 2007 [4243 m]). A recent analysis (Jansen and Basnyat, 2011) contended that velocities in the ICA and MCA were 11.7% and 3.4% (mean 6.2%) higher as compared with normal values at sea level, yet still within the range of expected variation at sea level reported by others (Scheel et al., 2000a; 2000b). Additionally, it has recently been reported that Tibetan (Sherpa) high-altitude residents have comparable velocities in the MCA and PCA, as well as comparable flow and diameter in the MCA, with well-acclimatized low-landers (Smirl et al., 2013). Also, hematocrit and Ca
Lessons from CBF in Lowlanders at High Altitude During Progressive Ascent
Only two studies have measured serial changes in CBF during progressive ascent to high altitude, but the findings may help explain small discrepancies between studies. In 2011, Wilson et al. (2011) measured diameter and velocity in the MCA (using transcranial color-coded Duplex-ultrasound, TCCD) following partial acclimation to 5300 m (n=24), 6400 m (n=14), and 7950 m (n=5). Remarkable elevations (200%) in flow in the MCA occurred at 7950 m. Notably, the authors estimated ∼24% dilation of the MCA occurred at 6400 m. Dilation of the MCA further increased to ∼90% at 7950 m (Fig. 3) and was rapidly reversed with oxygen supplementation (Fig. 3). Cerebral oxygen delivery and oxygenation were maintained by commensurate elevations of CBF even at these extreme altitudes. In another recent study, CBF and MCA diameter were measured at 1338 m, 3440 m, 4371 m, and over time at 5050 m (Willie et al., 2013). Dilation of the MCA was observed upon arrival at 5050 m with subsequent normalization of CBF and MCA diameter by days 10–12. Such findings are consistent with unchanged diameter following 17 days at 5400 m (Wilson et al., 2011). It is important to note that according to Poiseuille's Law, flow is proportional to radius raised to the fourth power. Therefore, consistent with previous concerns about TCD (Giller, 2003), that the MCA dilates at such levels of hypoxemia indicates that previous studies using TCD at altitude (Baumgartner et al., 1994; Lucas et al., 2011) may have underestimated flow (see Fig. 2) and thus may explain differences between studies. These findings are particularly important because they suggest regional regulation of CBF occurs in both large and small cerebral arteries.

Changes in blood flow in the middle cerebral artery (MCA) upon progressive ascent to 7950 m. Data were collected following partial acclimation to 5300 m (n=24), at 6400 m (n=14), and at 7950 m (n=5). Remarkable elevations (200%) in flow in the MCA occurred at 7950 m following removal of breathing supplementary oxygen and breathing air for 20 min. Dilation (∼24%) of the MCA occurred at 6400 m, which was further increased to 90% at 7950 m. Oxygen supplementation at this highest altitude rapidly reversed the observed MCA vessel dilation (denoted by blue triangle). Elevations in CBF via cerebral vasodilation were adequate to maintain oxygen delivery, even at these extreme altitudes. Modified from Wilson et al. (2011).
Regional CBF Changes at High Altitude
Recent reports at sea level indicate greater reactivity to hypoxia in the brainstem than cortex (Willie et al., 2012; Ogoh et al., 2013). In addition, during isocapnic hypoxia, PET scan data reveal that cortical blood flow is less responsive to hypoxia than phylogenetically older areas of the brain (Binks et al., 2008). During a slow hiking ascent to 5050 m, a recent study (Willie et al., 2013) reported no regional difference in ICA or vertebral artery (VA), whereas upon rapid ascent to 5260 m, Subudhi et al., (2013) reported a greater elevation in VA flow compared to the ICA. Thus, it seems upon sudden exposure to hypoxia there is a preferential maintenance of brainstem blood flow. Although this response is of potential teleological relevance in order to maintain oxygen delivery to the cardiorespiratory control centers, the mechanisms underlying this response are unknown. Finally, it should be noted that in children (aged 6–13 years) a preliminary report indicates that although elevations in velocity in the middle and anterior cerebral arteries occurred following ascent to 3500 m from sea level, velocity in the posterior and basilar arteries were otherwise unchanged (Gavlak et al., 2013).
Regulation of CBF at High Altitude
The small cerebral arterioles are anatomically located on the brain's surface tethered within the subarachnoid space, and surrounded by CSF. Thus, the tone of the pial vessels is a function of arterial blood gases, arterial pH, local CSF, and the metabolic status of the downstream cortical tissue. There may also be some independent effects of Ca
The initial factors that determine the magnitude of change in CBF depend upon the relative strengths of four reflex mechanisms (Severinghaus, 2001): 1) hypoxic ventilatory response (and hence the degree of respiratory alkalosis); 2) hypercapnic ventilatory response; 3) hypoxic cerebral vasodilatation, and 4) hypocapnic cerebral vasoconstriction (Fig. 4). The well-known within- and between-individual variability in these factors likely highlights the variability in changes in CBF upon initial arrival at altitude and difficultly in isolating independent effects of the respective mechanisms. The subsequent onset of ventilatory acclimatization coincides with the fall in CBF due to an increase in Pa
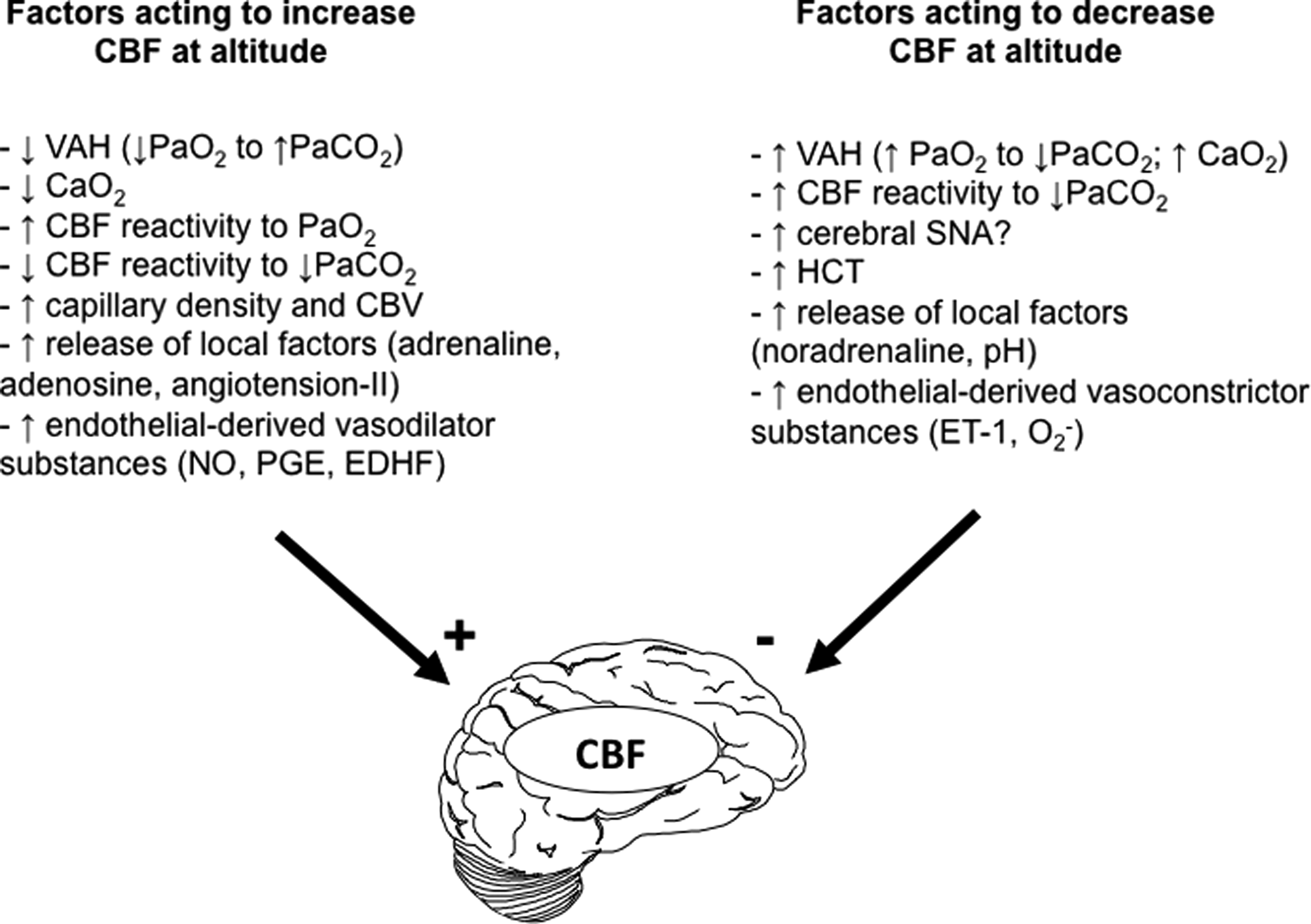
Summary of the major factors acting to increase (plus) and decrease (minus) CBF during exposure to hypoxia. Ca
In addition to these reflex responses, which most likely adjust during the acclimatization process, CBF is also influenced by a myriad of other hypoxic-induced changes [e.g., capillary density (angiogenesis), adenosine, nitric oxide, blood viscosity/hematocrit, hypoxia-inducible factor, vascular endothelial growth factor, and free radicals] (Ainslie et al., 2014). However, it should be noted that little information is available on the influence of these factors in relation to changes in CBF at altitude in humans. For example, although studies at sea level during acute hypoxia have indicated a contributing role of NO (Van Mil et al., 2002) and, to a lesser extent, adenosine (Bowton et al., 1988; Nishimura et al., 1993) in mediating cerebral vasodilation, these experiments have not been conducted at altitude. Much further work is needed to unravel the complex interactions that are responsible for the changes seen in CBF over time at high altitude.
Acute Mountain Sickness and CBF
There has been a long-standing debate concerning the role that increased CBF may play in the pathogenesis of AMS (e.g., Jensen et al., 1990; Baumgartner et al., 1994; 1999; Roach and Hackett, 2001). Although some early findings indicated that increased CBF (as assessed via TCD) is higher in those with AMS than those without (Baumgartner et al., 1994), this has not been confirmed in later studies (Jensen et al., 1990; Baumgartner et al., 1999; Ainslie et al., 2008; Lucas et al., 2011; Smirl et al., 2013; Subudhi et al., 2013; Imray et al., 2014). Figure 5 combines data from four different field studies (Ainslie et al., 2008; Lucas et al., 2011; Smirl et al., 2013; Subudhi et al., 2014) and, as shown, there is no relationship between changes in CBF (or velocity) with AMS. Nevertheless, such data should be interpreted with caution since it is possible that: 1) increased CBF may act or contribute to the development of AMS in the face of other risk factors (e.g., metabolic/genetic profile, sleep, exercise); 2) changes in a global CBF may not reflect localized dilation of cerebral vessels in discrete areas of the brain contributing to AMS symptomology; and 3) that the symptomology of AMS is similar to myriad ailments commonly experienced during field studies to high altitude

Lack of relationship between changes in CBF with symptoms of AMS. Data combined from field studies (Ainslie et al., 2008; Lucas et al., 2011; Smirl et al., 2013; Subudhi et al., 2013). Each point represents an individual measurement of CBF and AMS symptoms. Red data points are from Subudhi et al., (2013).
Future Directions
It is clear that many aspects of CBF regulation and brain function at high altitude warrant further investigation. Indeed, several questions remain. For example, over the period of ventilatory acclimatization (weeks to months), how do interactions between the hypoxic ventilatory response, hypercapnic ventilatoy response, hypoxic cerebral vasodilatation, and hypocapnic cerebral vasoconstriction interact to alter CBF? Furthermore, what is the role of NO and/or adenosine in mediating cerebral vasodilation at high altitude? And last, what is the time-course of recovery in CBF following descent to sea level?
Our understanding of CBF regulation in Tibetan and Andean high altitude residents is also limited. Continuing fieldwork is therefore urgent since modernization and migration are changing the traditional ways of life and patterns of exposure to the environment among highlanders everywhere. Some future questions include: How is CBF and its regulation altered following treatment of chronic mountain sickness and related lowering of hematocrit or following Diamox administration? Although it is clear that the brain is highly sensitive to environmental hypoxia, little is known about the effects of high altitude residence in the developing brain. It is known that the immature brain undergoes a protracted period of development into the second decade of life (Niermeyer et al., 2009). However, although a recent report highlights a differential change in regional cerebral velocities in children upon ascent to 3500 m (Gavlak et al., 2013), it is uncertain how longer term hypoxic stress affects CBF in children. Addressing some of these intriguing questions will not only provide new information by which CBF is regulated at high altitude, but will also provide insight into the understanding of cerebral hypoxia in the clinical setting (e.g., with chronic lung and cardiovascular diseases).
Footnotes
Acknowledgments
This brief review was inspired by the Cerebral Blood Flow in Hypoxia: From Early Human Experiments to New Discoveries session at the 2013 International Hypoxia Symposia in Lake Louise, Alberta, Canada. The authors appreciate editorial assistance of John Severinghaus and thank Anthony Bain and Chris Willie for their feedback on this manuscript. PNA is support by a Canada Research Chair in Cerebrovascular Physiology.
Author Disclosure Statement
No competing financial interests exist.