
Abstract
Abstract
Rubenstein, David A, and Wei Yin. Hypergravity and hypobaric hypoxic conditions promote endothelial cell and platelet activation. High Alt Med Biol 15:396–405, 2014.—Cardiovascular disease risk is heightened during exposure to altered gravity and/or altered barometric conditions. Previous work has suggested that this heightened cardiovascular risk is due to enhancements of endothelial cell inflammatory and/or thrombogenic responses. In recent work, the role of platelets on instigating or inhibiting endothelial cell responses associated with cardiovascular disease has been found to be dependent on both biochemical and biophysical factors. In this work, we aimed to determine how two biophysical forces, gravity and atmospheric pressure, alter endothelial cell and platelet functions and their interactions to instigate or inhibit cardiovascular disease responses. To address this aim, endothelial cells and platelets were subjected to a force 8 times greater than the normal gravitational force, for up to 30 minutes. In separate experiments, endothelial cells and platelets were subjected to 50% of normal atmospheric pressure. Endothelial cell and platelet responses, associated with cardiovascular diseases, were measured as a time course during exposure. In general, the exposure of endothelial cells to either hypergravity or hypobaric conditions enhanced cardiovascular disease responses. However, the presence of platelets generally inhibited endothelial cell responses. Platelet activation was, however, somewhat enhanced under both hypergravity and hypobaric conditions. Our data suggest that altered biophysical forces can modulate endothelial cell and platelet responses that are salient for cardiovascular disease progression. However, the interaction of these two cells tends to restrain the progression of the pro-cardiovascular disease responses.
Introduction
C
Gravitational forces (or g-force) are felt by any object under the influence of gravity that is not under free-fall. For instance, the g-force that is felt by an object residing on the Earth's surface at sea level and approximately 45 degrees latitude is defined as 1-g. During space flight, astronauts experience both hypergravity (e.g., take-off) and hypogravity (e.g., orbiting) conditions. Standard atmospheric pressure is defined as 760 mmHg at sea level. Atmospheric pressure varies as a function of altitude above the Earth's surface. During extravehicular activities, astronauts can be subjected to hypobaric conditions. When these biophysical forces deviate significantly from the standard values, they may induce adaptation or pathophysiological responses in the host system (Oyama and Platt, 1965; Pitts et al., 1983; Bozzini et al., 2005). It is our goal to determine how changes in these forces alter cardiovascular disease responses and the role of loading duration on cardiovascular responses.
There has been significant effort in the past to determine the role of altered gravity on cardiovascular functions. Altered gravitational loading is common for astronauts, high-speed jet pilots, and amusement park thrill seekers. Oshima et al. (2005) have shown that a mild exposure to hypergravity, 2-g or 3-g for 4 hours, increases the expression of cyclooxygenase-2, hypoxia-inducible factor-1α, inducible nitric oxide synthase, vascular endothelial growth factor, and heme oxygenase-1, in the mouse cardiac circulation. All of these factors have been implicated in inflammatory cardiovascular disease progression. Others have shown that dendritic cells and dermal fibroblasts reorganize their cytoskeleton and interact with vascular cells in response to hypergravity (10-g or 15-g) (Croute et al., 1995; Bellik et al., 2008), which can lead to enhanced cardiovascular disease processes. Endothelial cells have also showed an impaired angiogenic response and changes to their cytoskeletal organization in response to hypergravity (3-g or 10-g, respectively) for longer exposure durations (up to 48 hours) (Spisni et al., 2002; Monici et al., 2006). Interestingly, over this long exposure duration, there were no changes to endothelial cell metabolism, viability, or other standard culture parameters (Spisni and others, 2002).
There are two central issues regarding the exposure to hypobaric conditions; are the biological responses observed, a) due to the changes in atmospheric pressure or b) due to changes in oxygen concentration. Previous work has compared cellular responses to normobaric hypoxia, hypobaric hypoxia, and hypobaric normoxia, showing that neither hypoxia nor hypobaric conditions by itself typically elicit a significant cardiovascular response, whereas the combination of hypobaric and hypoxic conditions is typically more detrimental and elicits cardiovascular responses that are associated with cardiovascular disease progression (Levine et al., 1992; Savourey et al., 2003). For instance, it has been shown that hypobaric hypoxia can induce tachycardia, a reduction in coronary blood flow, and a decrease in cardiac output, which are all associated with cardiovascular disease progression (reviewed in (Hultgren and Grover, 1968)). Endothelial cells have been shown to have an enhanced expression of intercellular adhesion molecule-1, E-selectin, and major histocompatibility complex class II after exposure to hypobaric hypoxic conditions (Dore-Duffy et al., 1999). Thus, there is evidence supporting the role of hypobaric hypoxia inducing endothelial cell responses related to cardiovascular disease progression.
Platelets have an enhanced aggregation, adhesion to von Willebrand factor, and an enhanced expression of glycoprotein Ibα and integrin αIIβIII, after exposure to 8-g for 15 minutes (Dai et al., 2009; Li et al., 2009). Interestingly, the tail bleeding time for mice exposed to short duration hypergravity was significantly decreased (Dai et al., 2009). Combined, these studies suggest that short duration exposures to hypergravity can increase the thrombotic responses of platelets. The majority of reports that investigated the role of hypoxia or hypobaric hypoxia on platelet functions/coagulopathies, have shown that these conditions elicit no significant effects on coagulopathy that can lead to pathological events (Maher et al., 1976; Bartsch et al., 1989; Crosby et al., 2003; Schobersberger et al., 2007). This has been confirmed by population-based studies of high altitude residents, who show no differences in the incidence of coronary arterial thrombosis, coagulability, and deep vein thrombosis (Hurtado, 1955; 1960), as compared to lower altitude residents. However, it appears that platelets and other thrombotic responses can adapt to hypobaric hypoxic conditions, because there is evidence of coagulopathies after short-term exposure (8 hours) to hypobaric hypoxia (Bendz et al., 2000). Additionally, some investigations find specific changes to coagulation processes under hypobaric hypoxic conditions, such as a shortened activated partial thromboplastin time (aPTT), but these groups were unable to link this to any pathological change in coagulation protein concentration, including fibrin (O'Brodovich et al., 1984). The role of endothelial cells and the mechanisms behind these platelet changes have yet to be elucidated.
Exposure to hypergravity or hypobaric hypoxic conditions has been shown to induce responses in endothelial cells and platelets that have been linked to cardiovascular disease progression. However, the interaction of these cell types during hypergravity or hypobaric hypoxic conditions have not been investigated. Here, we aimed to determine the role of exposure duration and combined incubation of endothelial cell and platelets on cellular responses that are common to cardiovascular diseases.
Methods and Materials
Endothelial cells and platelets
Human umbilical vein endothelial cells (HUVECs) were obtained from ScienCell Research Laboratories (Carlsbad, CA, USA) as passage 1 cells and were used between passages 2 and 7 for all experiments. Cells were maintained in endothelial cell media (ECM) supplemented with 5% fetal bovine serum, 1X endothelial cell growth supplement, 10 U/mL penicillin and 10 μg/mL streptomycin (all from ScienCell). Passage at confluence was by trypsin digestion for no more than 3 min. Platelet rich plasma (PRP), anti-coagulated with 0.32% sodium citrate, was purchased from Oklahoma Blood Institute (Oklahoma City, OK). PRP was centrifuged at 1100 g to obtain washed platelets, which were maintained in 1X modified HEPES-Tyrode's buffer supplemented with 10% bovine serum albumin (pH 7.4). For all experiments, platelet concentration was adjusted to 100,000/μL with HEPES-Tyrode's buffer. For experiments when endothelial cells and platelets were incubated together, the total volume of ECM and HEPES-Tyrode's buffer was held constant (either 2 mL or 500 μL, depending on the response measurement), but the ratio of buffers was set based on the post-wash platelet count, which averaged 522,000±27,000/μL (mean±SEM, n=57).
Exposure to hypergravity/hypobaric hypoxia
Cells were exposed to what we termed “hypergravity via centrifugation” (Sorvall Legend T plus, Thermo Scientific) at 300 rpm at room temperature for various time durations. This generated a g-force equal to 8-g on the cells. Paired normogravity controls were keep on a laboratory bench at room temperature for the same time duration. Cells were exposed to hypobaric hypoxia (termed as hypobaric conditions) in a standard electronic vacuum chamber, which maintained an atmospheric pressure of 380 mmHg at 37°C for various time durations. Paired controls for these conditions were maintained at local atmospheric pressure (which averages to about 735 mmHg over 2012) at 37°C for the same time durations.
Endothelial cell response variables
A standard MTT assay was used to determine the metabolic activity of endothelial cells after exposure to hypergravity or hypobaric hypoxic conditions. All details of the MTT assay have previously been reported (Rubenstein et al., 2011). To confirm metabolic activity changes, a live/dead cell cytotoxicity assay, composed of calcein and ethidium, was used. All details of the live/dead cell cytotoxicity assay have been reported (Rubenstein and others, 2011). Briefly, cell viability is defined as the total number of live cells divided by the total number of cells. Cell density is normalized to the cell seeding density, which was set at ∼1000 endothelial cells/cm2 and provides a measure of proliferation.
Immunohistochemistry was used to determine the cytoskeleton reorganization in response to hypergravity or hypobaric hypoxia. Briefly, after exposure, cells were tri-stained with 5 U/mL phalloidin (which fluoresces when associated with F-actin), 5 μg/mL wheat germ agglutinin (which fluoresces when associated with the cell membrane) and 300 nM DAPI. Cells were first fixed with 1.5% glutaraldehyde, then nonspecific binding was blocked with 100 mM glycine+0.1% albumin and finally permeabilized with 0.2% Triton-X. The exact protocol has been previously reported (Maria et al., 2011). Briefly, the percent alignment quantifies the percent of the cytoskeleton that aligns with the major loading force direction (hypergravity) or the alignment direction (hypobaric). Percent structure quantifies the amount of fibrillar actin to globular actin. The expression of intercellular adhesion molecule-1 and tissue factor was quantified to monitor the extent of cardiovascular disease marker activation. A standard enzyme linked immunosorbent assay was used to monitor ICAM-1 and tissue factor expression. Briefly, cells were fixed (0.5% glutaraldehyde) and blocked (100 mM glycine+0.1% BSA) as above. Primary antibodies were incubated with cells for 60 min, after which appropriate secondary antibodies were incubated the cells for 60 min. All details have been previously reported (Maria and others, 2011).
Finally, the relative expression of ICAM-1 and tissue factor mRNA, was quantified with RT-PCR. Briefly, after the exposure conditions, cells were treated with 4°C Trizol (Invitrogen, Grand Island, NY) for 5 min, after which cells were collected and ice cold chloroform was added to individual samples. Samples were centrifuged for 15 min at 11,300 rpm (4°C), after which the aqueous phase was transferred into isopropanol and high precipitation salt solution (0.8 M sodium citrate and 1.2 M NaCl, Molecular Research Center, Cincinnati, OH) for 10 min on a rotator. Samples were centrifuged for 10 min at 11,300 rpm (4°C) and the supernatant was removed. The RNA pellet was resuspended in ice cold 75% ethanol and then centrifuged for 5 min at 8900 rpm (4°C). After centrifugation, the ethanol was removed and the pellet was dried and resuspended in pure water, after which the samples were incubated on a heating block for 10 min at 58°C. Potential DNA contamination was removed using a DNA free kit (Invitrogen) following the manufacturer's instructions. Once pure RNA was obtained, the RNA was reverse transcribed with qScript cDNA supermix (Quanta Biosciences, Gaithersburg, MD) following the manufacturer's directions. After cDNA generation, SYBR green PCR Master Mix (Invitrogen) was used. The forward and reverse primers used are shown in Table 1.
Platelet response variables
After exposure to hypergravity or hypobaric conditions, timed platelet samples were assayed for the surface expression of GPIIb (CD41) and P-selectin (CD62P) and for adhesion to endothelial cells. Surface expression was quantified with flow cytometry as described previously (Rubenstein and others, 2011). Briefly, fluorphore-conjugated antibodies against human CD41 or CD62P were incubated with platelets for 30 min at room temperature. After incubation, surface expression was quantified immediately using an Accuri C6 flow cytometer (Ann Arbor, MI). Adhesion was quantified via the difference in platelet concentration before and after exposure to hypergravity or hypobaric conditions (as quantified by a Z2 Coulter Counter). Platelet counts were averaged from three samples of the supernatant to obtain a relative measure of adhesion.
Statistics
All hypergravity endothelial cell or platelet experiments were paired with a normogravity experiment for the same experimental duration (as a control). All hypobaric endothelial cell or platelet experiments were paired with a normobaric experiment for the same experimental duration (as a control). The only exception to this was the platelet adhesion quantification as described above. All experimental data points were normalized to the aggregate data from the paired control. Data were then pooled from a minimum of 4–5 independent experiments and reported. A multi-way ANOVA was conducted using SAS (v9.3) with α set as 0.05 and β set as 0.8. The possible factors for the multi-way ANOVA, with a least-squares means post-hoc test, were gravity or atmospheric pressure, duration, and presence of platelets.
Results
Endothelial cell metabolic activity
The metabolic activity of endothelial cells as a function of exposure duration to altered gravity or altered atmospheric pressure was investigated (Fig. 1). The presence of platelets did not alter the metabolic activity of endothelial cells under these conditions, and thus the pooled data are plotted for ease of viewing. After exposure to hypergravity or hypobaric conditions, there was no difference in endothelial cell metabolic activity as compared with normogravity or normobaric conditions. Although there was a marginal increasing trend of endothelial cell metabolic activity after exposure to hypergravity and a decreasing trend for normogravity conditions, as a function of time, this was only significantly different at the 20 minute time point. Thus, endothelial cell metabolic activity was not a function of altered gravity, altered atmospheric pressure, exposure duration or presence of platelets, as investigated here.
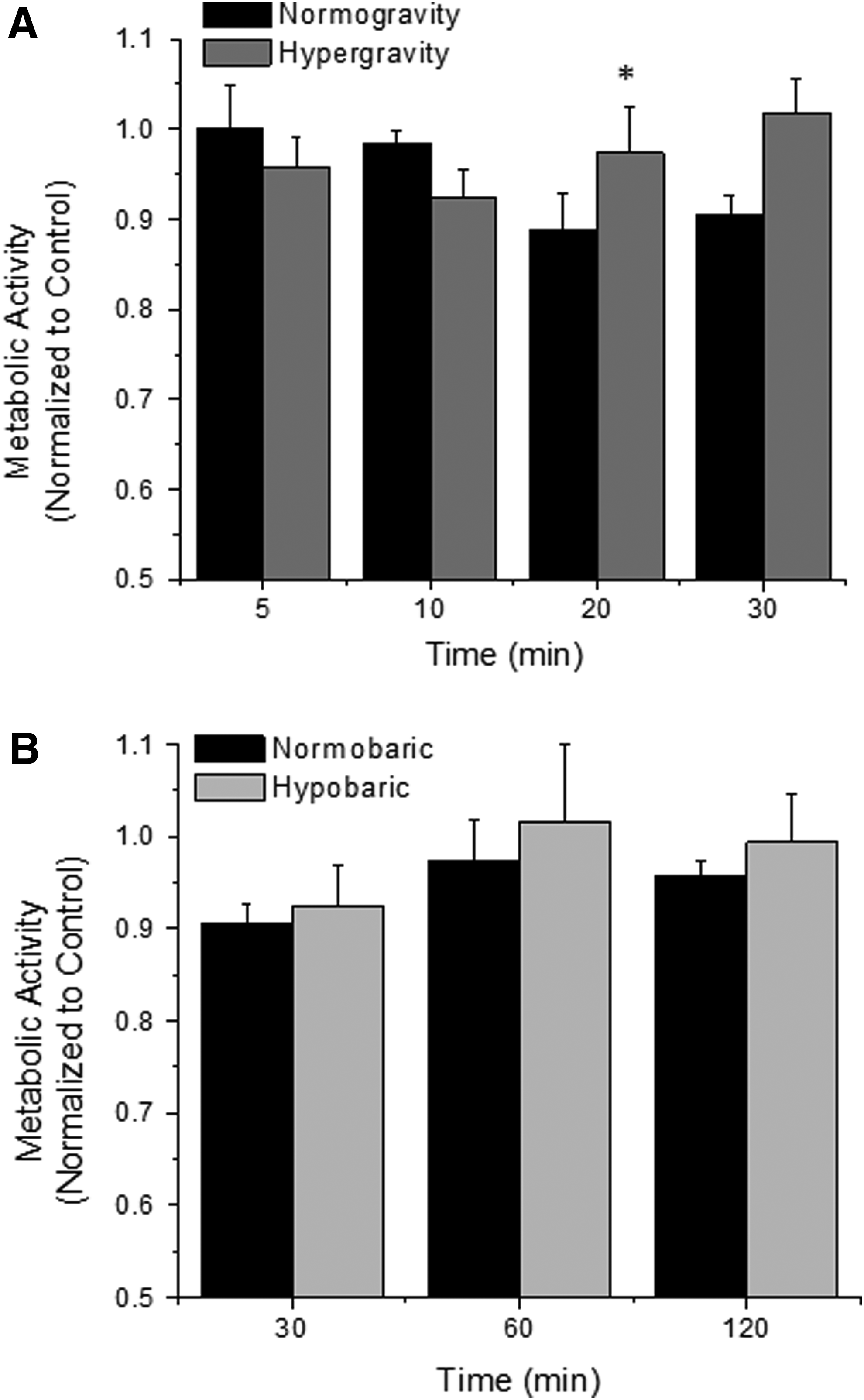
Metabolic activity of endothelial cells exposed to hypergravity
Endothelial cell viability and density
The viability of endothelial cells was measured after exposure to hypergravity or hypobaric conditions to confirm metabolic activity data (Fig. 2 A,C).In parallel, the density of endothelial cells was also quantified (Fig. 2 B,D). Similarly to the metabolic activity data, the presence of platelets did not significantly alter endothelial cell viability or density and thus the data presented have been pooled for with or without platelets. The effects of hypergravity and hypobaric hypoxia on endothelial cell density was not a function of time, although there were nominal decreases and increases, respectively, during the experimental time course (Fig. 2B,D). Likewise, altered gravity did not significantly change the endothelial cell viability (Fig. 2A). However, there was a significant trend for an increasing viability after exposure to hypobaric conditions that was somewhat paralleled in the normobaric conditions (Fig. 2C). Overall, the effects of altered gravity or altered atmospheric pressure on viability and density, as compared with normogravity or normobaric conditions, were not as drastic as exposure duration; although at some time points, both hypergravity and hypobaric conditions increased viability. Thus, our data suggest that exposure duration to hypergravity or hypobaric conditions have the most significant effect on endothelial cell viability and density and not the exposure conditions themselves.

Cell viability
Cytoskeletal reorganization
The cytoskeletal reorganization of endothelial cells in response to hypergravity or hypobaric conditions was quantified (Fig. 3).Cytoskeletal dynamics were not a function of presence of platelets and in general, exposure duration to hypergravity or hypobaric conditions did not alter the reorganization of the actin cytoskeleton. The platelet factor has been pooled for plotting. Cytoskeletal organization is a measure of how the actin cytoskeleton can adapt to different loading conditions.

Actin cytoskeleton adaptation of endothelial cells exposed to hypergravity
Surface and mRNA expression of ICAM-1 and tissue factor
The relative surface expression of intercellular adhesion molecule-1 and tissue factor was quantified after exposure to hypergravity or hypobaric conditions to determine whether these conditions promote pro-inflammatory and pro-coagulant reactions in endothelial cells (Fig. 4, platelet factor is pooled). With a longer duration exposure to hypergravity or hypobaric conditions, there was an increased expression of both ICAM-1 and tissue factor as compared with the paired normogravity or normobaric conditions. Under standard gravity and standard pressure, there was no change in these response measurements as a function of time. For the most part, the protein expression data was correlated with the mRNA expression data (Fig. 5, platelet factor is pooled). With a longer duration exposure to hypergravity or hypobaric conditions, the expression of ICAM-1 and tissue factor mRNA increased and decreased, respectively, as compared with the normogravity and normobaric conditions. Interestingly, there was an early increase in tissue factor mRNA after exposure to hypobaric conditions and this appears to be recovered after longer exposures. Thus, our data suggest that hypergravity and hypobaric conditions can enhance pro-inflammatory and pro-coagulant responses.

ICAM-1
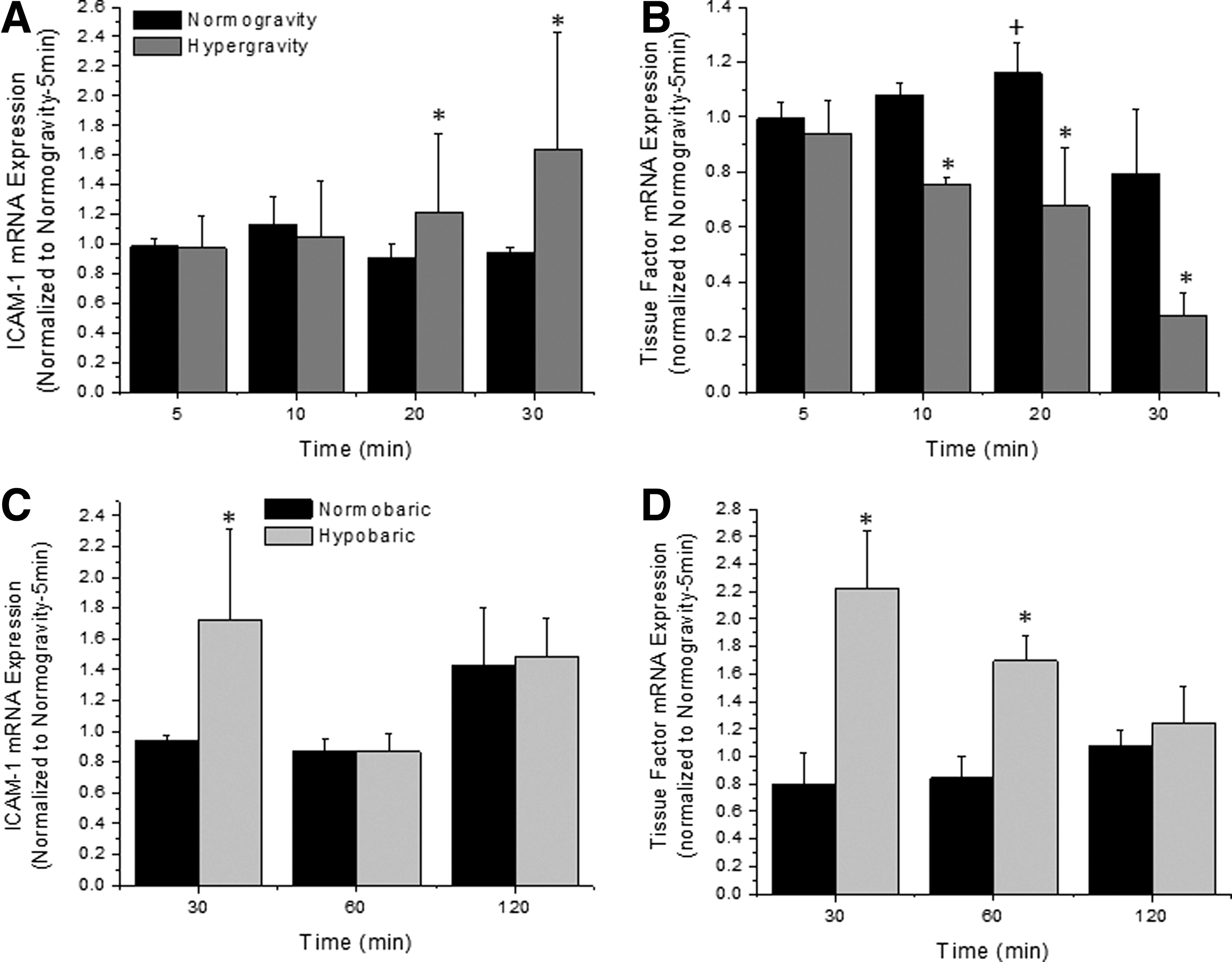
ICAM-1
Platelet activation and adhesion
The activation and adhesion potential for platelets exposed to hypergravity and hypobaric conditions was quantified via flow cytometry against the activation marker P-selectin and the adhesion marker GPIIb (Fig. 6 A,B).In parallel, platelet counts were obtained after exposure to hypergravity or hypobaric conditions to quantify adhesion to endothelial cells (Figure 6 C,D). Exposure to both hypergravity and hypobaric conditions induced a significant increase in CD62P and CD41 expression. With longer duration exposures, platelets subjected to hypergravity showed an overall increase in marker expression, while platelets subjected to hypobaric conditions showed an early increase that was somewhat recovered over the exposure duration. Platelet adhesion to endothelial cells followed a similar trend, with an overall increase as a function of time and exposure to hypergravity. Platelets that were exposed to hypobaric conditions did not differ significantly from the paired normobaric conditions. Thus, our data suggest that platelets are both sensitive to altered gravity, altered atmospheric pressure, and the exposure duration to these conditions.

Platelet expression of CD41 and CD62P
Discussion
Metabolic activity/viability/density
We quantified the metabolic activity (Fig. 1) via the activity of mitochondrial dehydrogenase, and the viability and density (Fig. 2) of endothelial cells after exposure to hypergravity or hypobaric conditions as a function of the exposure duration and presence of platelets. Overall, our data suggested that altered gravity or altered atmospheric conditions and the presence of platelets do not affect endothelial cell viability, density, or metabolic activity. However, the exposure duration to these conditions had some effects on cell viability and density. Koyama et al. (2009) quantified an increase in ATP release after exposure to mild hypergravity (2G/3G for 5 min), and suggested this may alter proliferation but did not quantify this. This was confirmed by others, using slightly different exposures and measurement techniques (Versari et al., 2007). Our data suggest that mild hypergravity does not alter cell proliferation. Toff et al. quantified various properties of endothelial cells and platelets after exposure to hypobaric hypoxia and deduced that these conditions did not alter various thrombotic, inflammatory, and other processes (Toff et al., 2006). Our data would somewhat agree with these findings and show that the presence of platelets do not alter these endothelial cell responses. Note that there are some statistically significant data points, as a function of time, but the scale of these changes is so minimal that it does not likely induce a biological effect.
Actin cytoskeleton organization
Versari et al. (2007) quantified changes to the actin cytoskeleton after long-term exposure to hypergravity. Their data show a mild reorganization of the actin cytoskeleton after 96 hours of stimulation. Koyama et al. (2009) show a transient change in the actin cytoskeleton in response to short-term hypergravity stimulation. While their data show some subtle qualitative differences, our method of quantification would not necessarily detect these differences (Fig. 3). Overall our data agree with previous quantitative findings. To the best of our ability, we could not identify previous work that investigated the role of hypobaric conditions on the endothelial cell cytoskeleton. Others have shown that hypobaric conditions disrupt the cytoskeleton of cardiac myocytes, skeletal muscle, or smooth muscle cells (Hall et al., 2000; Lemler et al., 2000; Holm et al., 2010). With our short exposure to hypobaric conditions, we did not see an adaptation of the endothelial cell cytoskeleton, but again, we cannot directly compare this to any previous work.
Protein/mRNA expression of ICAM-1 and tissue factor
We quantified the inflammatory and thrombotic response of endothelial cells after exposure to hypergravity and hypobaric conditions. In general, we showed an increase in the surface expression of these pro-inflammatory and pro-thrombotic markers (Fig. 4) that was somewhat matched at the transcript level for ICAM-1 expression (Fig. 5). Tissue factor transcript concentration does not match the surface expression, possibly due to altered transport of stored tissue factor. This is in agreement with previous findings that showed an altered inflammatory and thrombotic response after exposure to hypobaric hypoxia (Nakanishi et al., 1998; Dore-Duffy et al., 1999) or hypergravity (Gueguinou et al., 2012). Interestingly, the presence of platelets did not alter the expression of these markers.
Platelet activation and adhesion potential
The most salient finding of this work is the response of platelets to altered gravity and altered atmospheric conditions. There have been a few studies that reported an increase in the hemostatic response of platelets after exposure to hypergravity (Dai et al., 2009; Li et al., 2009) or hypobaric (Nakanishi et al., 1997) conditions. Here, we report a significant increase in the platelet activation marker P-selectin and adhesion molecule GPIIb/IIIa (Fig. 6). In parallel, we were able to observe an increased platelet-endothelial cell interaction after exposure to hypergravity, which has not been reported before. However, what was most interesting was that this increased platelet activity did not alter basic endothelial cell functions (e.g., metabolism, proliferation) but increased inflammatory and thrombotic responses associated with cardiovascular disease progression.
Conclusion
There have been multiple previous reports detailing the effects of hypergravity and/or hypobaric hypoxia on various endothelial cell or platelet functions. The novelty of this article is that we combined the altered gravity or altered atmospheric pressure incubation with a more physiologically relevant culture system (e.g., platelets and endothelial cells). Most of the previous work showed that both mild and extreme exposures to hypergravity or hypobaric conditions significantly upregulated endothelial cell and platelet pro-inflammatory and pro-thrombotic responses and inhibited cell growth and adaptation. We show here that in a more physiologically relevant setting, the pro-inflammatory/pro-thrombotic responses are still enhanced under these conditions, but the maintenance cellular responses return to baseline. Thus, this may reduce the overall chances for cardiovascular events to occur, since endothelial cells can adapt and proliferate normally, but in a true physiological setting, other responses/factors will occur simultaneously that may affect cardiovascular disease progression. To note, some of the limitations of our study include the use of an in vitro model system to study a complex physiological process, a constant short-term exposure to altered gravity or altered atmospheric pressure and the exposure to altered gravity or altered atmospheric pressure were maintained at a constant value.
Footnotes
Acknowledgments
The authors would like to thank Ms. Tahanee Mujib and Mr. Saravan Shanmugavelayudam for technical assistance with these experiments.
Author Disclosure Statement
The authors have no conflicts to disclose regarding the publication of this work.